Incubation of Negative Affect during Protracted Alcohol Withdrawal Is Age-, but Not Sex-Selective


 and
and
Abstract
1. Introduction
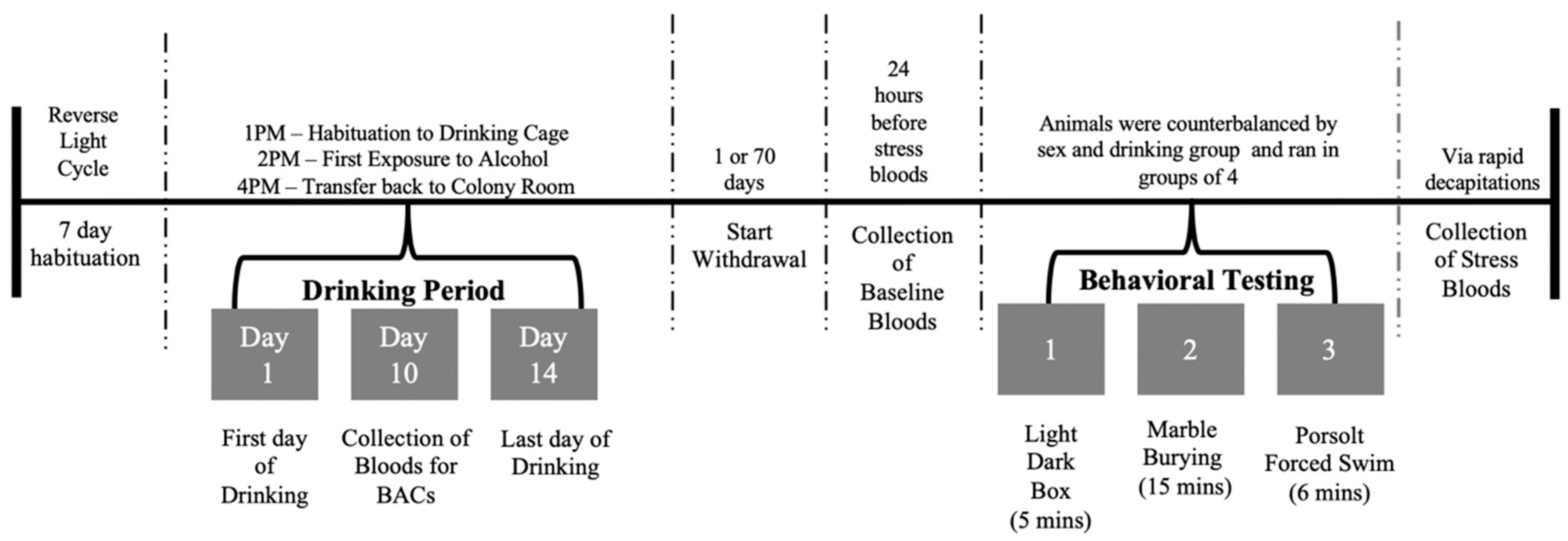
2. Materials and Methods
2.1. Subjects
2.2. Drinking-In-The-Dark (DID) Procedures
2.3. Blood Alcohol Concentration Sampling
2.4. Gas Chromatography
2.5. Baseline and Stressor-Induced CORT Assay
2.6. Behavioral Testing
2.6.1. Light–Dark Shuttle-Box
2.6.2. Marble-Burying Test
2.6.3. Porsolt Forced Swim Test
2.7. Statistical Analysis
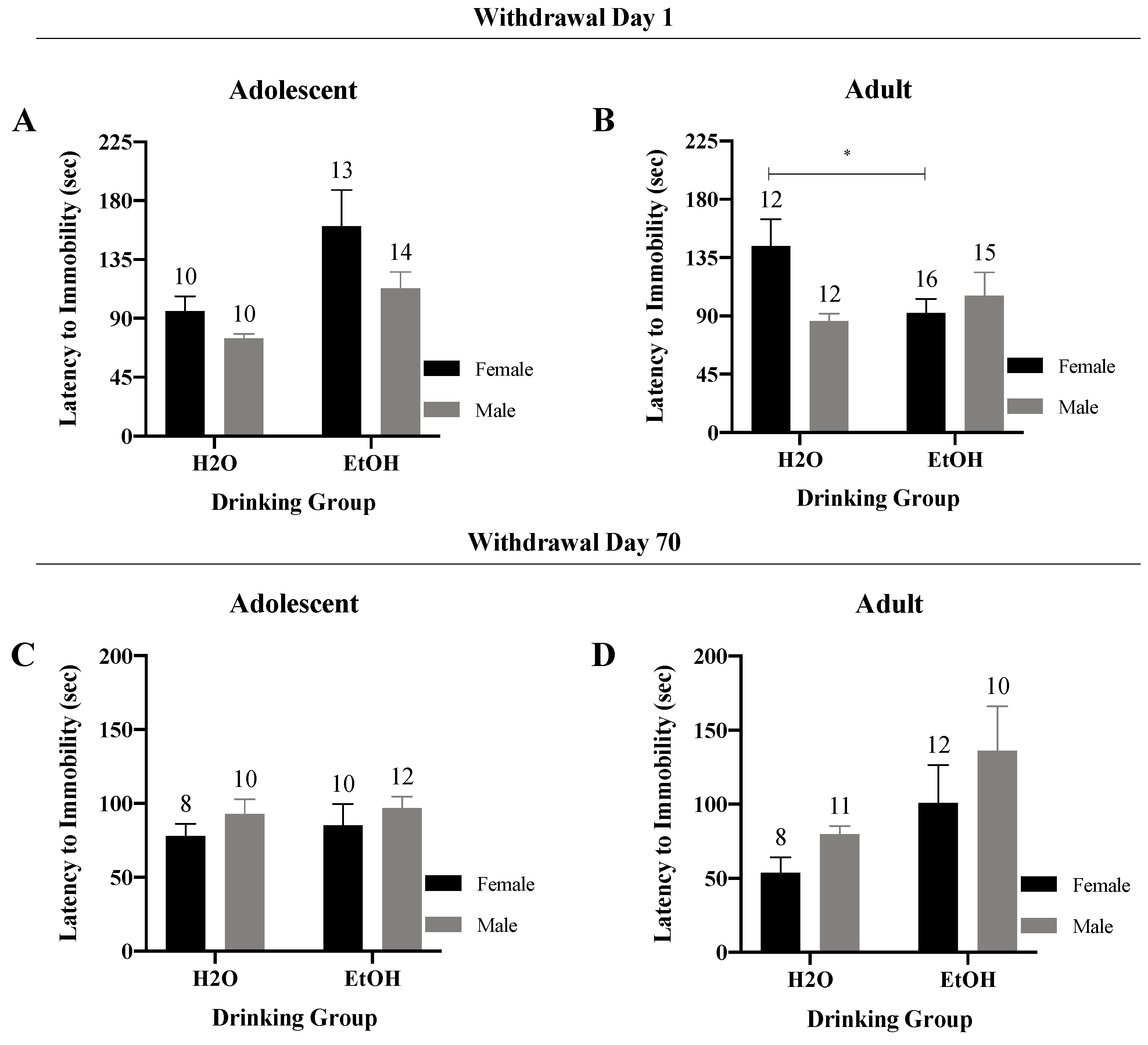
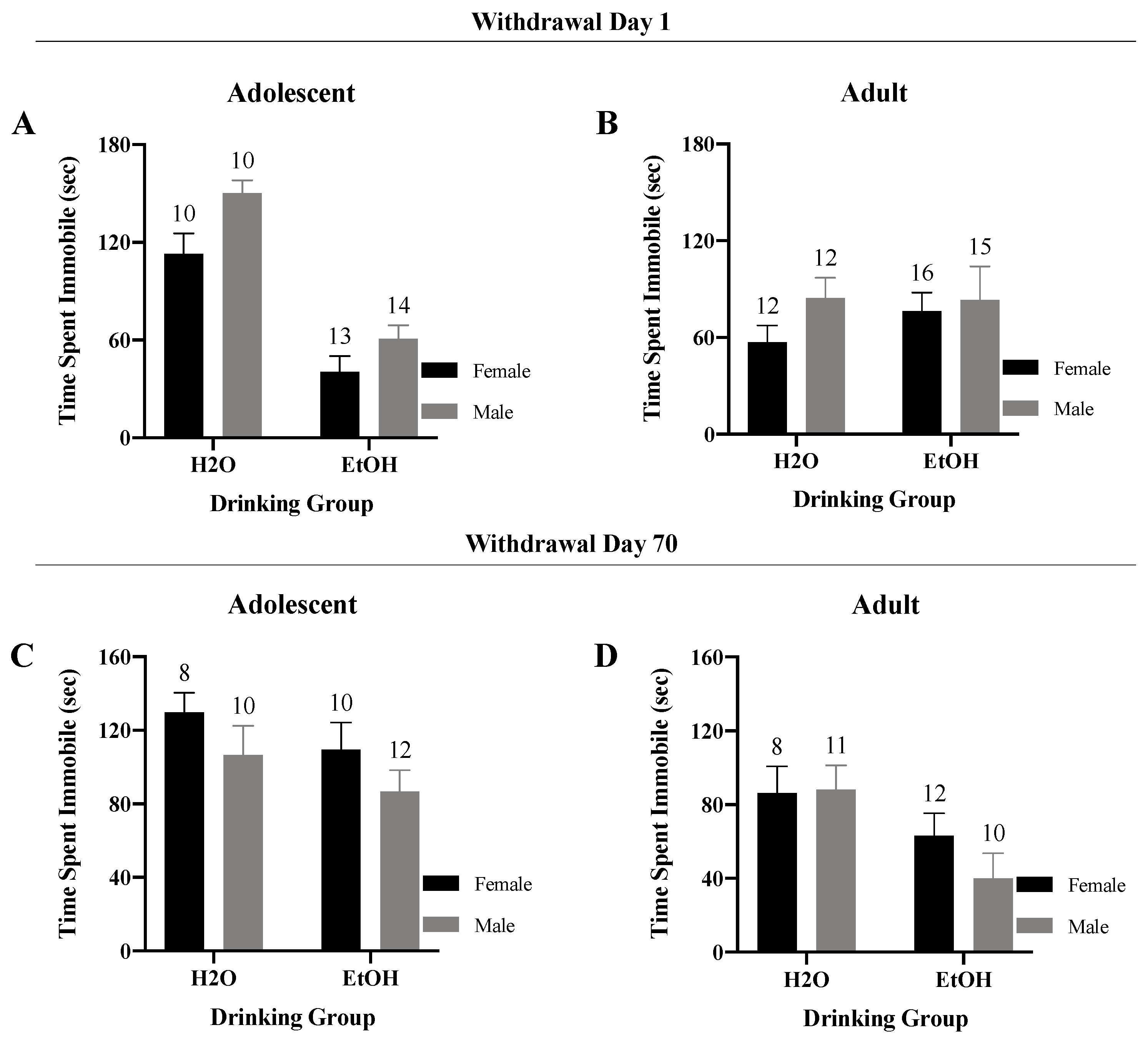
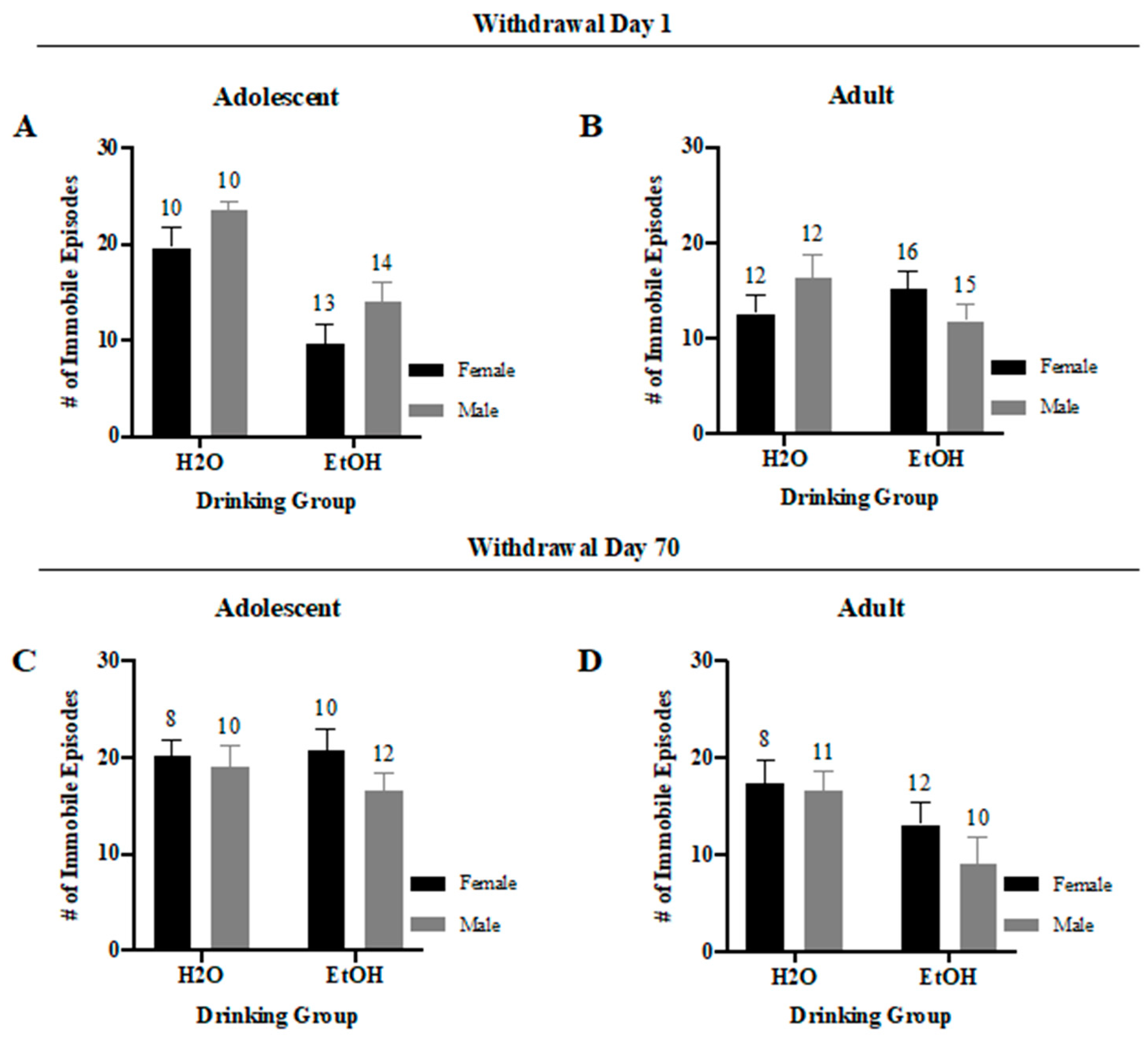
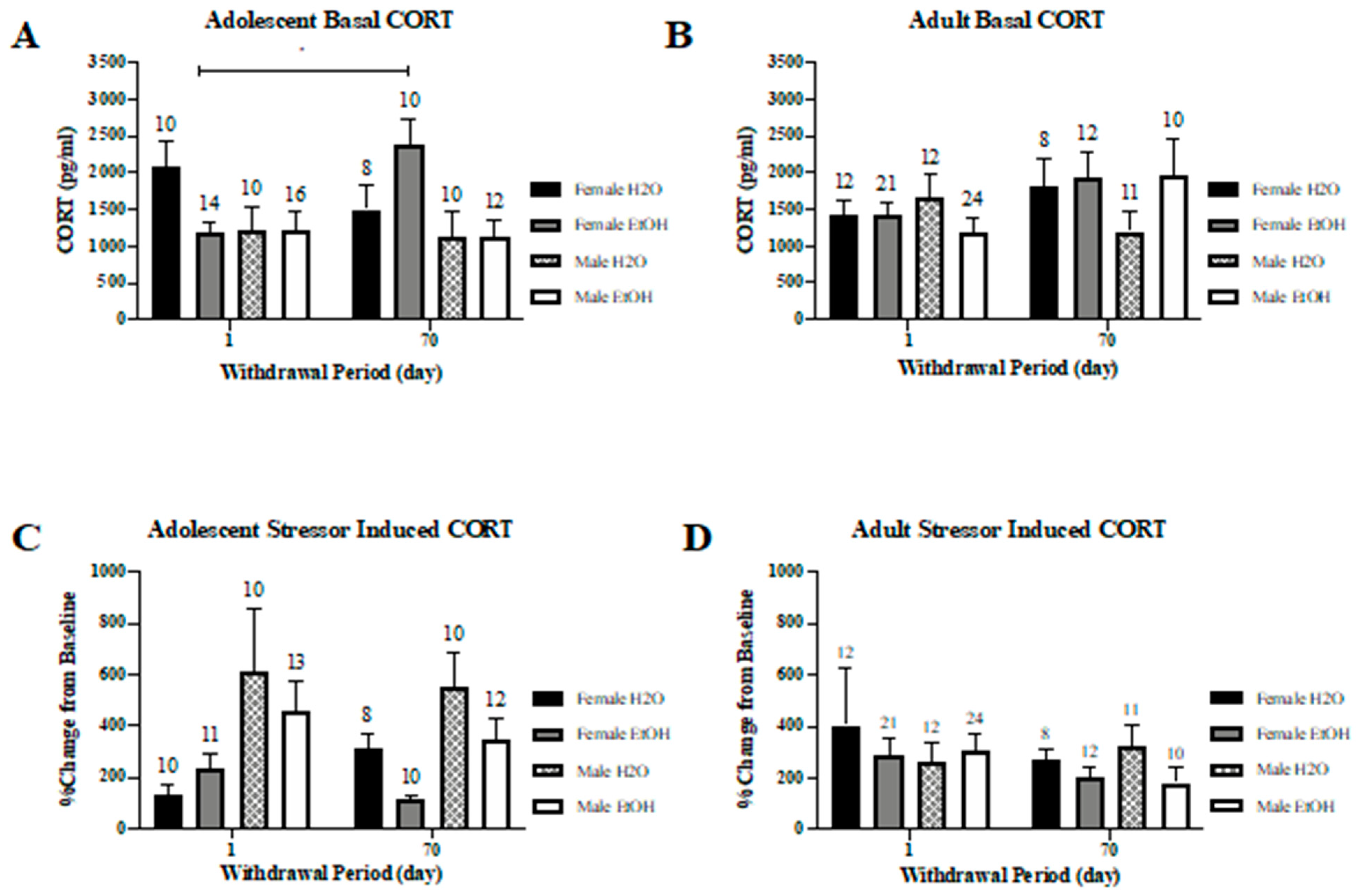
3. Results
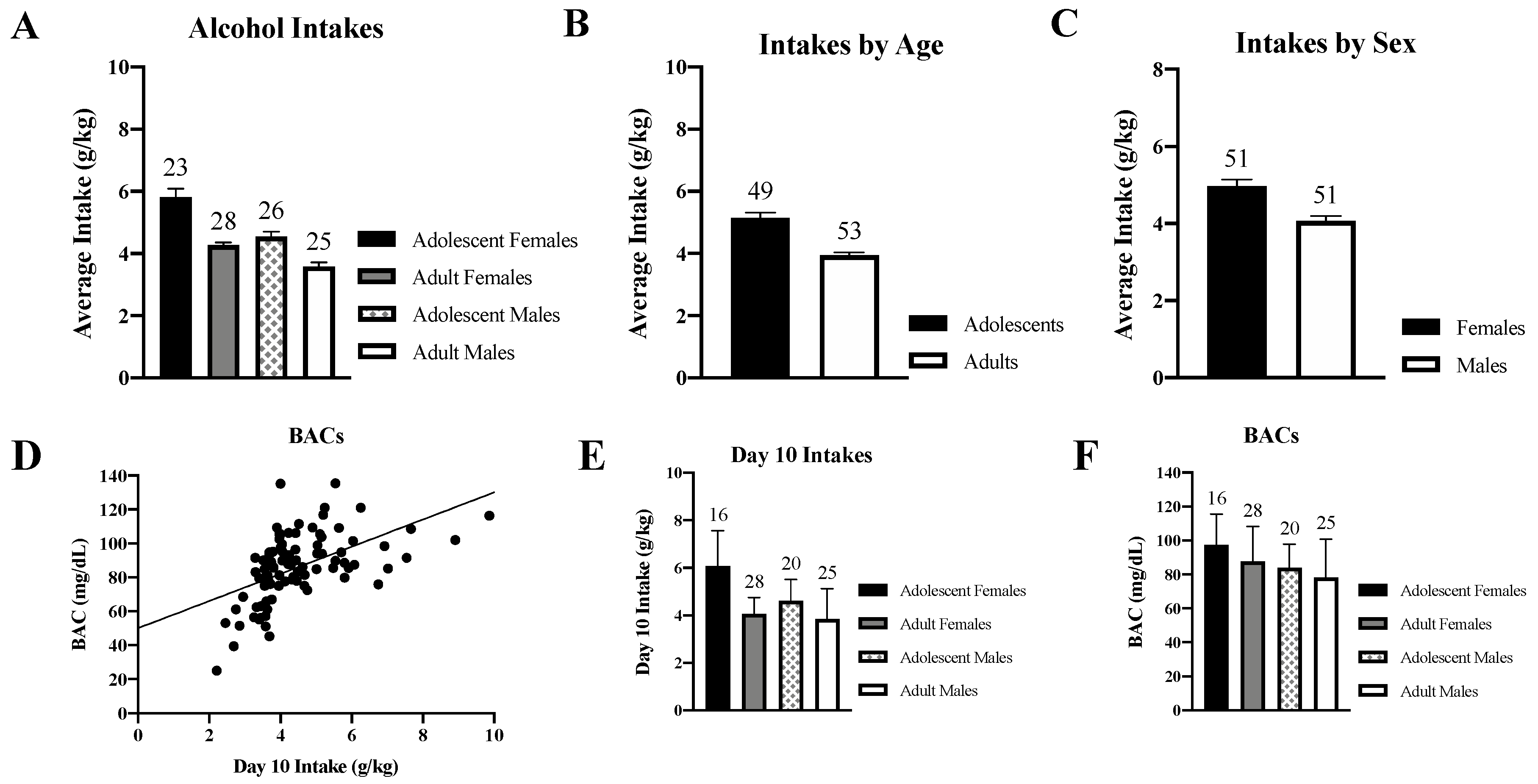
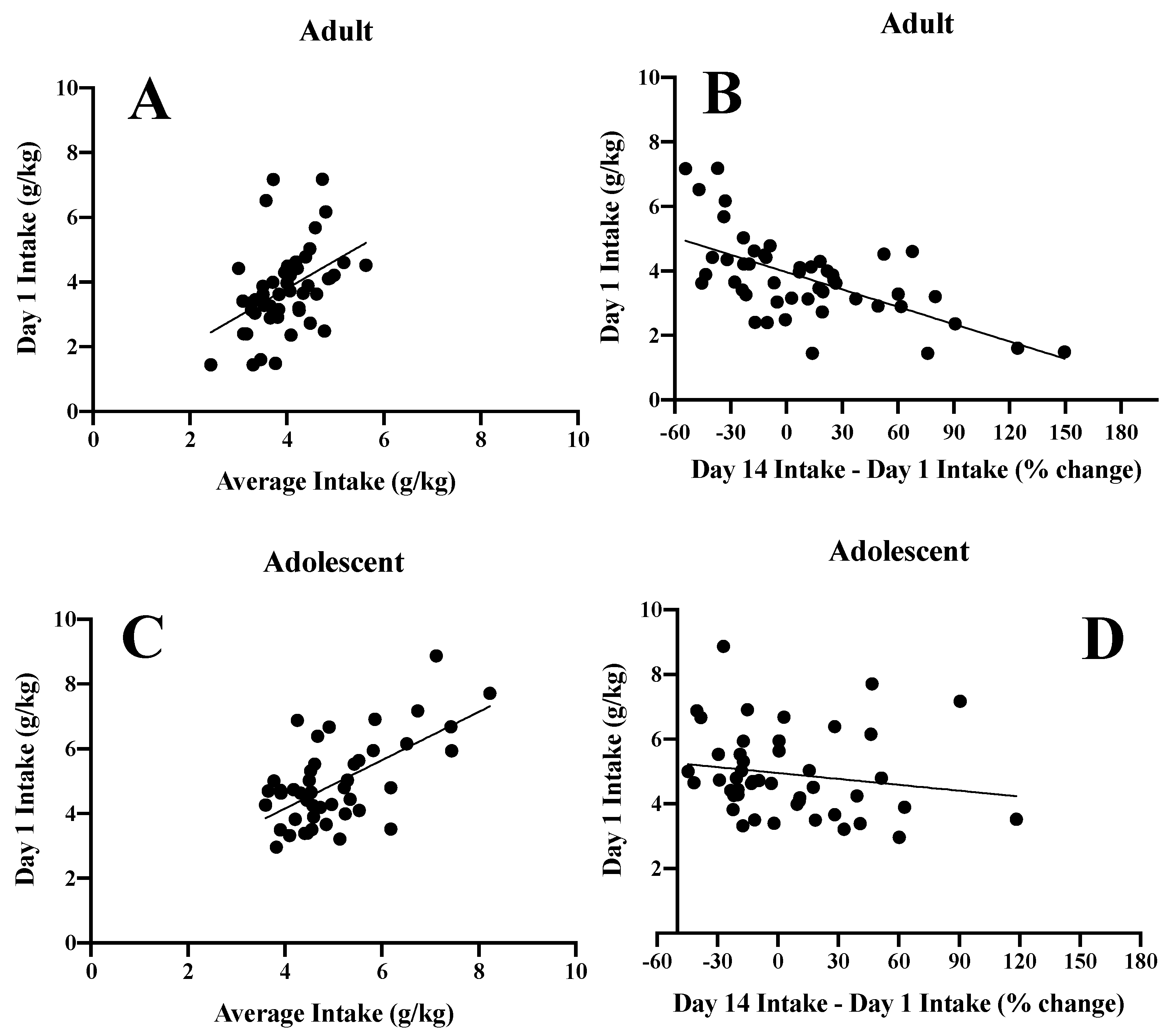
3.1. Alcohol Intake and Blood Alcohol Concentrations
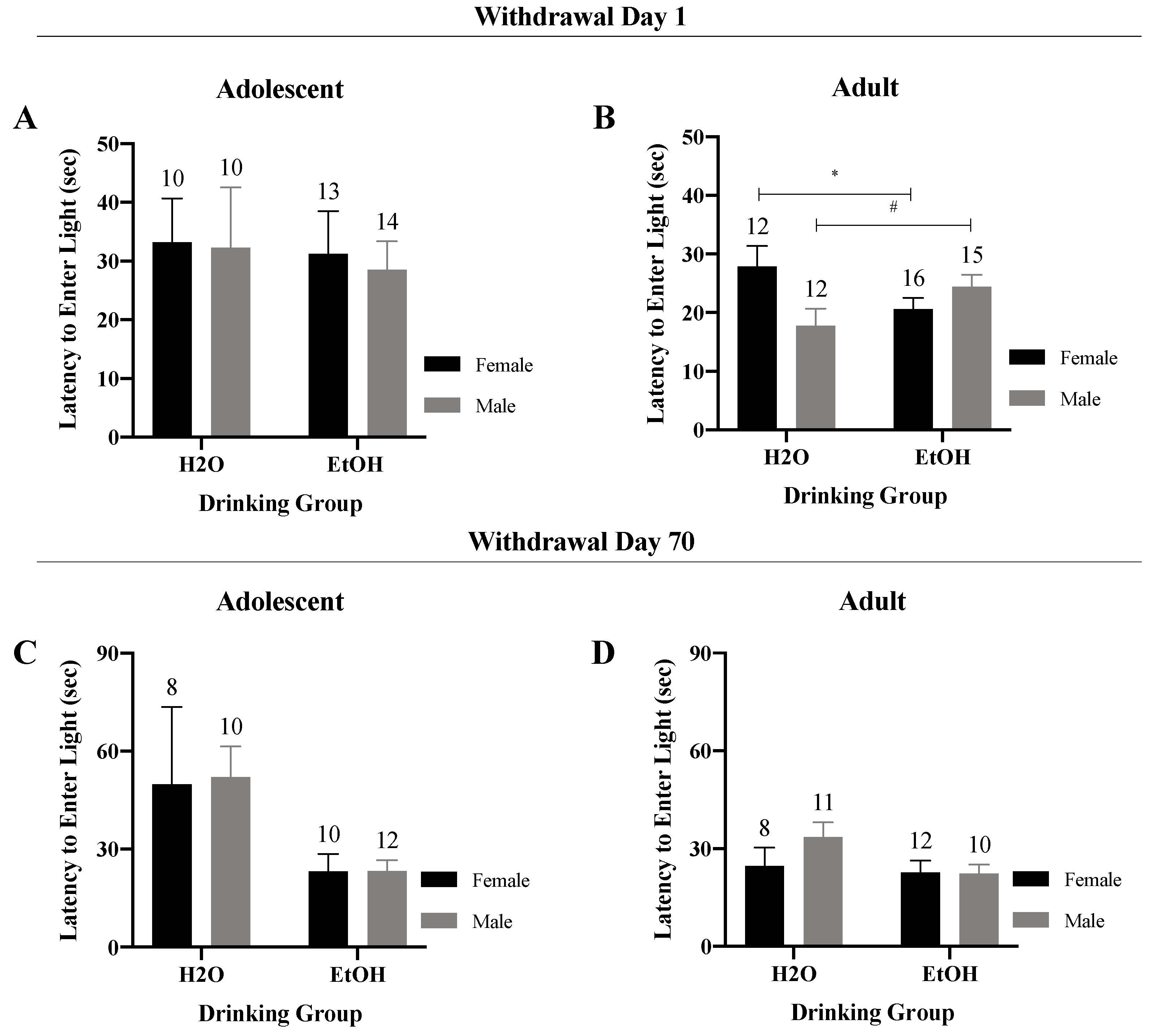
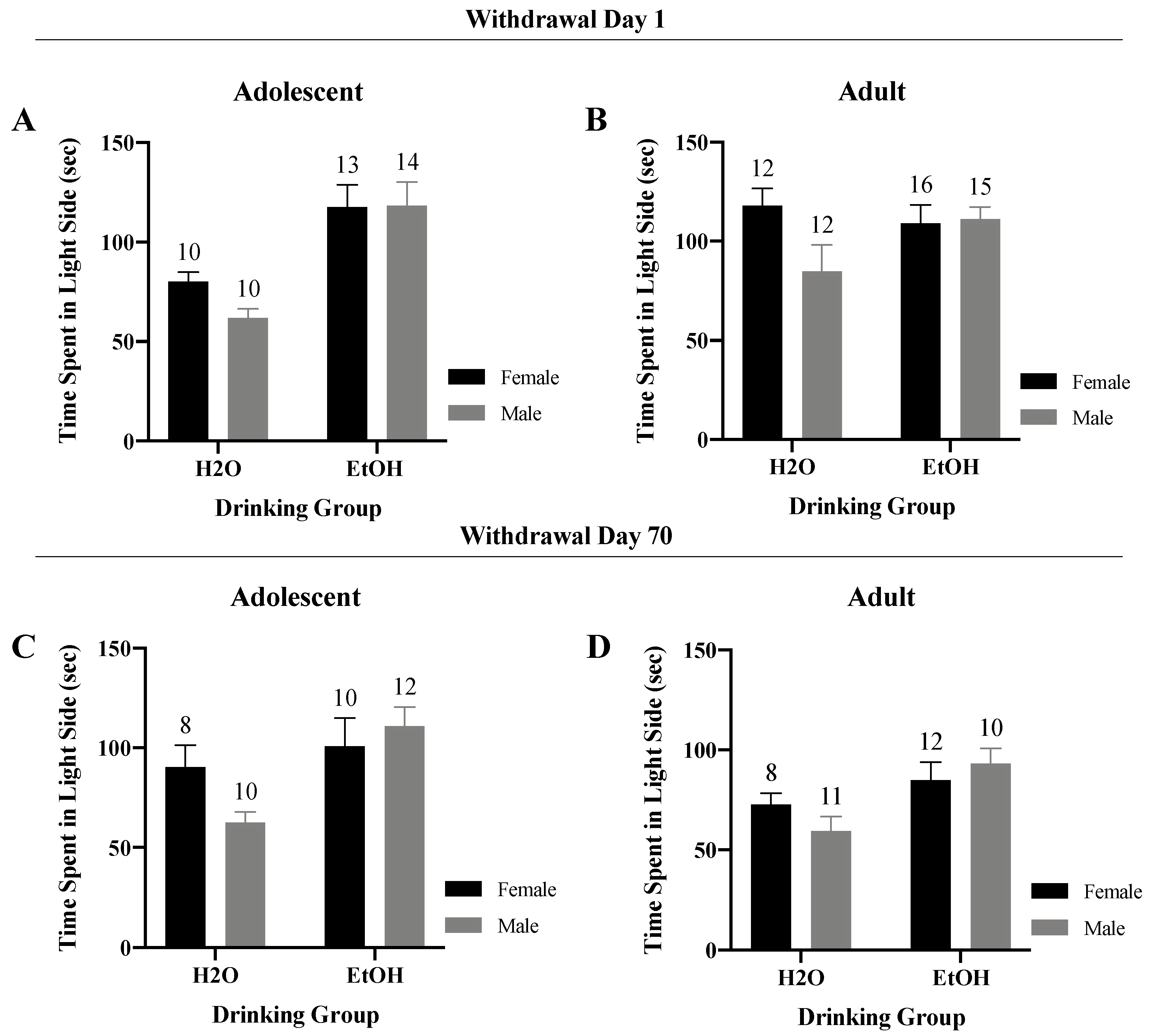
3.2. Light–Dark Shuttle-Box
3.2.1. Latency to Enter the Light Side
3.2.2. Time Spent in the Light Side
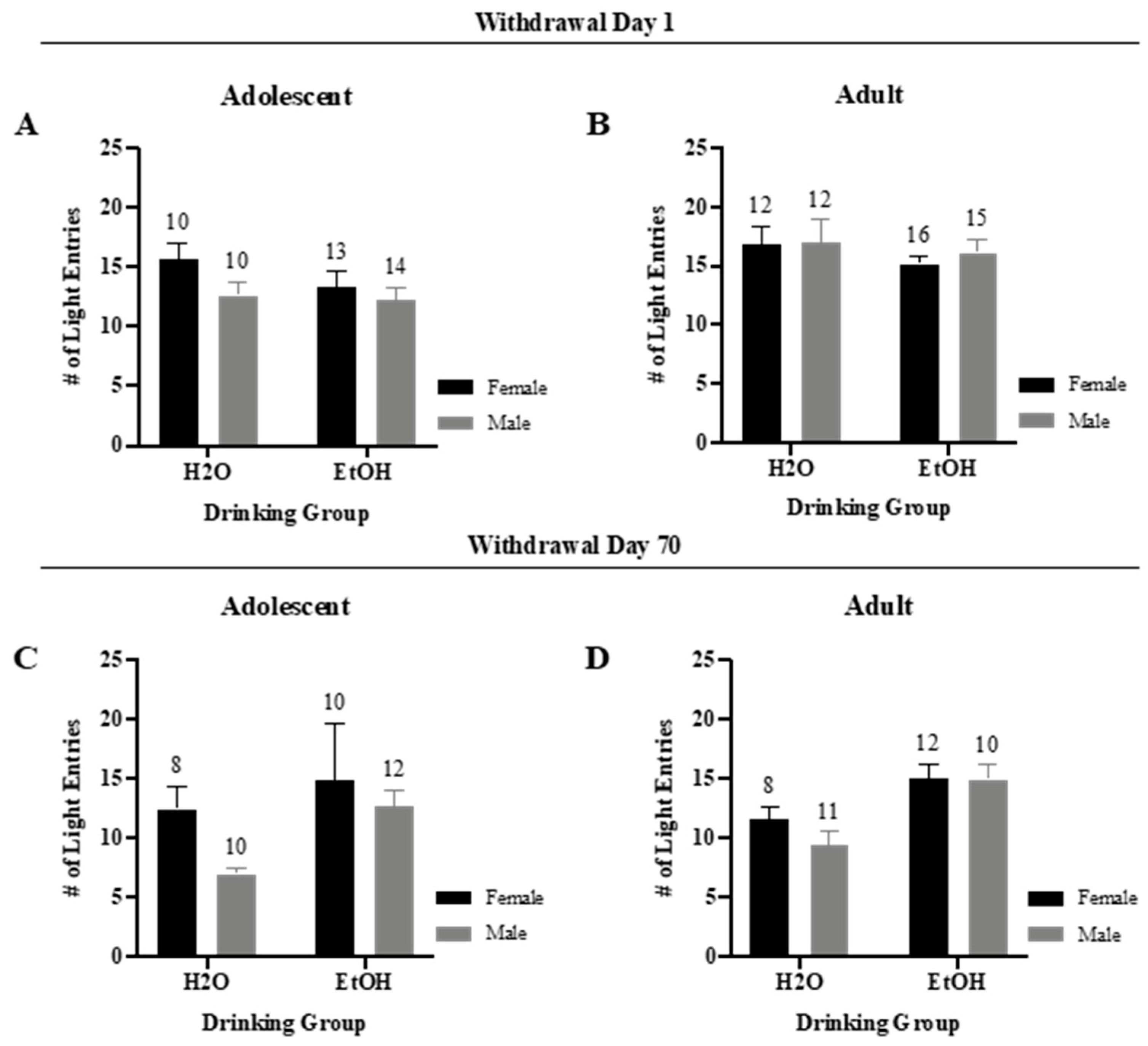
3.2.3. Number of Light Entries
3.3. Marble Burying Test
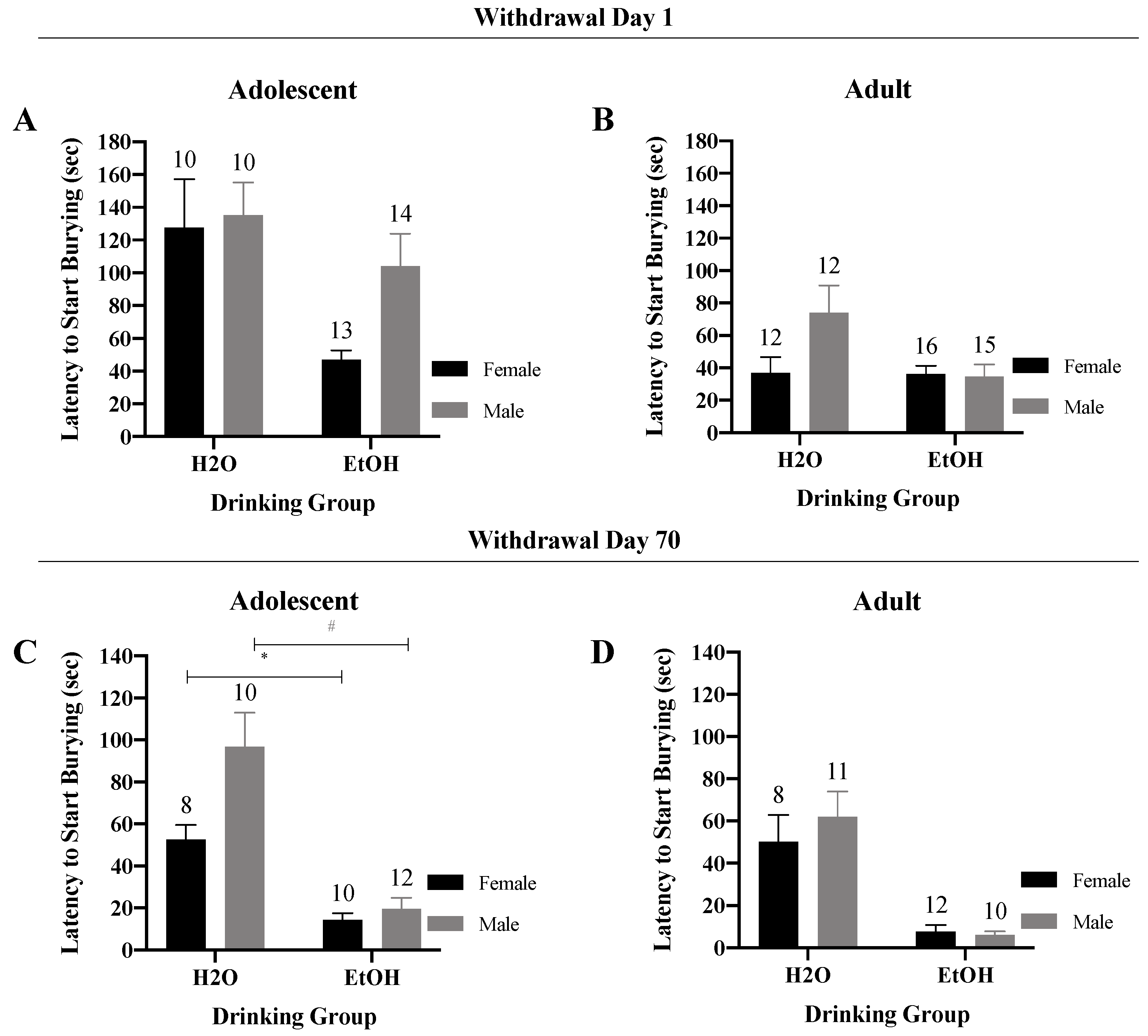
3.3.1. Latency to Start Burying
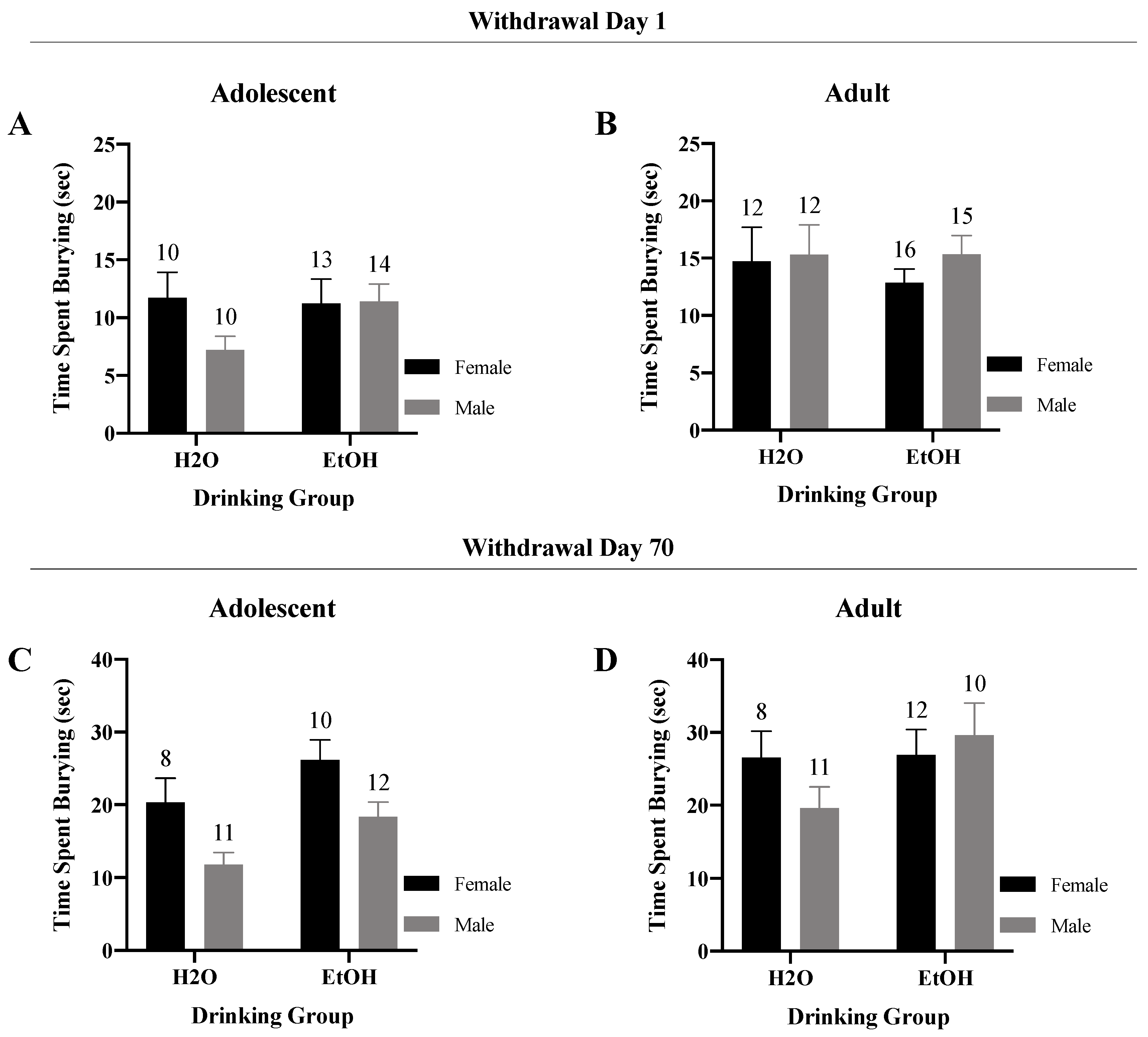
3.3.2. Time Spent-Burying
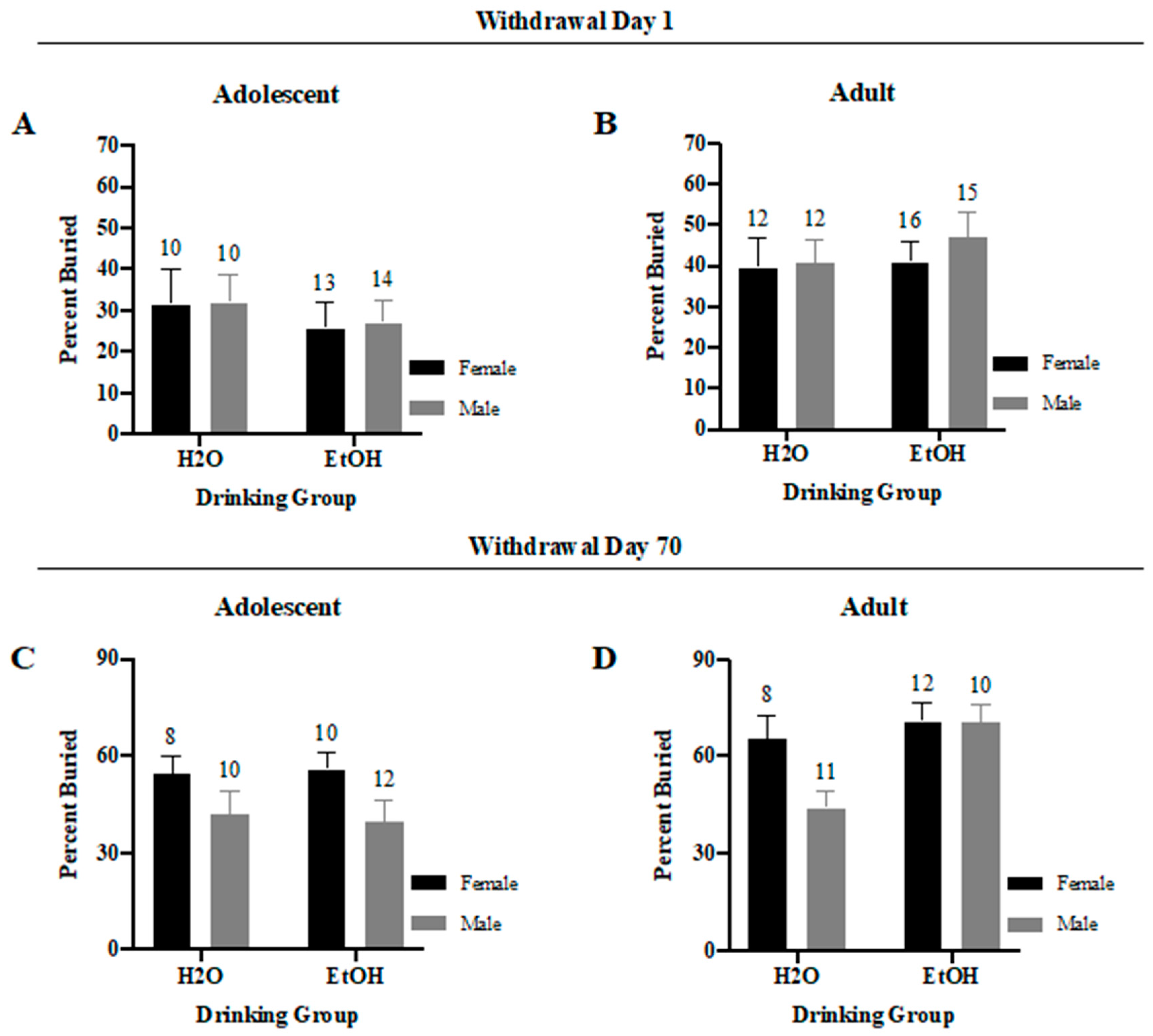
3.3.3. Percent of Marbles Buried
3.4. Porsolt Forced Swim Test
3.4.1. Latency to Immobility/Floating
3.4.2. Time Spent Immobile
3.4.3. Immobile Episodes
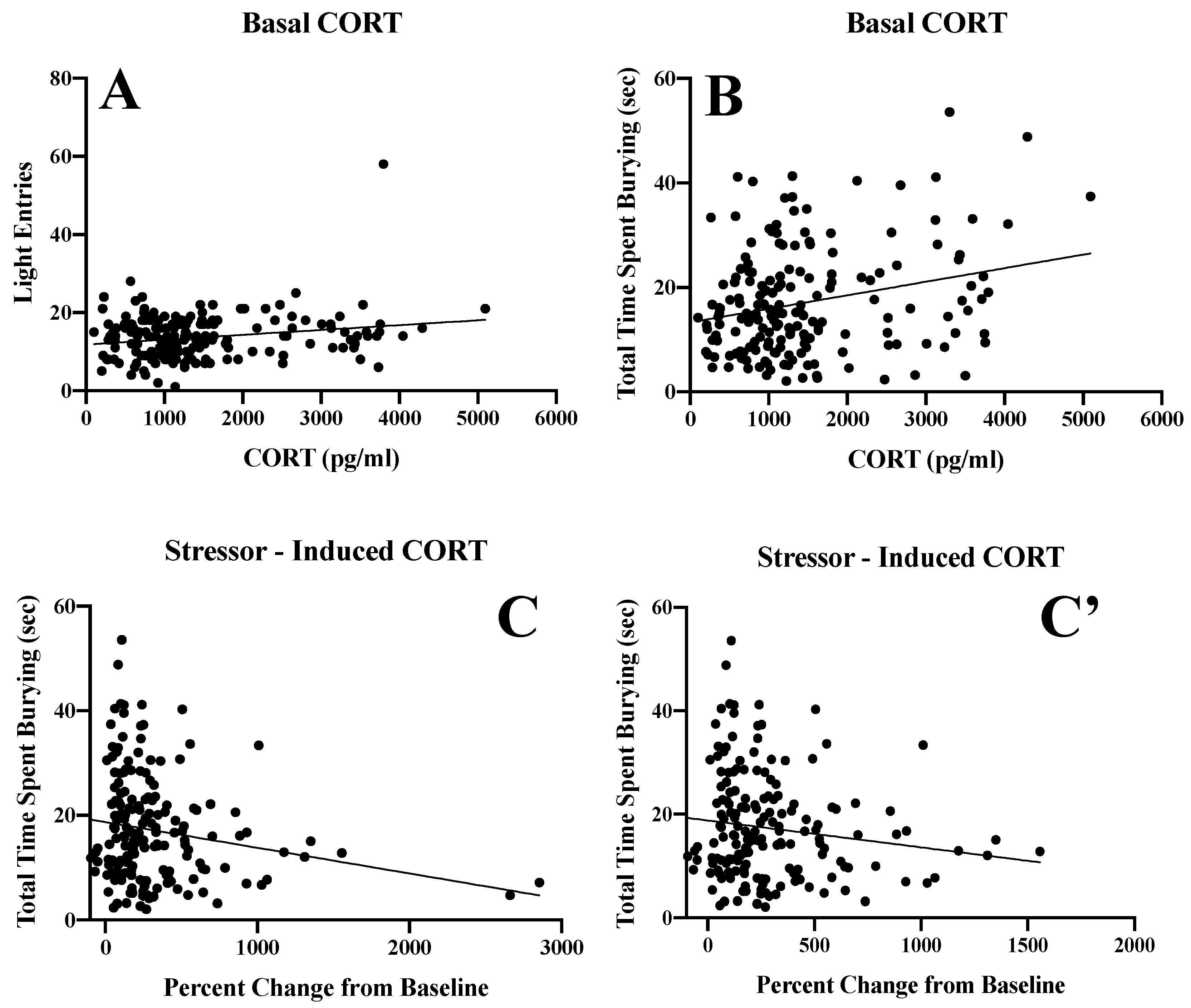
3.5. Corticosterone Assay
3.5.1. Basal Corticosterone
3.5.2. Stressor-Induced Corticosterone
3.5.3. Inter-Relations between Alcohol Intake and Corticosterone Levels
3.5.4. Inter-Relations between Corticosterone Levels and Behavioral Indices of Negative Affect
3.6. Inter-Relations between Initial Alcohol Intake and Subsequent Alcohol Consumption
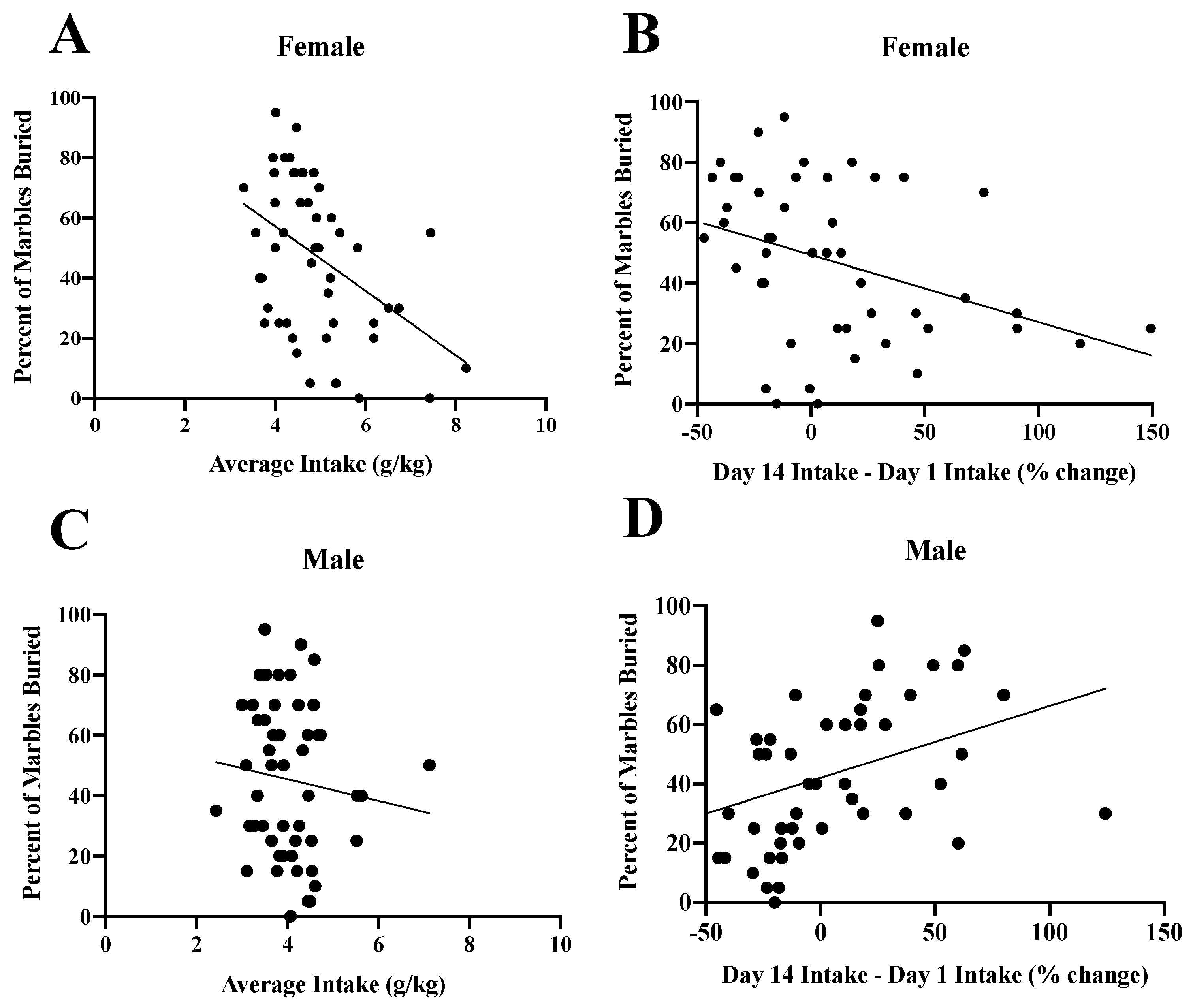
3.7. Inter-Relations between Indices of Alcohol Intake and Behavioral Indices of Negative Affect
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- National Institute on Alcohol Abuse and Alcoholism (NIAAA). NIAAA Council Approves Definition of Binge Drinking; NIAAA Newsletter: Rockville, MD, USA, 2004.
- National Institute on Alcohol Abuse and Alcoholism (NIAAA). Underage Drinking: Fact Sheet. Available online: https://pubs.niaaa.nih.gov/publications/underagedrinking/Underage_Fact.pdf (accessed on 21 January 2018).
- Guerri, C.; Pascual, M. Mechanisms involved in the neurotoxic, cognitive, and neurobehavioral effects of alcohol consumption during adolescence. Alcohol 2010, 44, 15–26. [Google Scholar] [CrossRef]
- Kelley, A.; Schochet, T.; Landry, C. Risk taking and novelty seeking in adolescence: Introduction to part I. Ann. N. Y. Acad. Sci. 2004, 1021, 27–32. [Google Scholar] [CrossRef]
- Spear, L.P. The adolescent brain and age-related behavioral manifestations. Neurosci. Biobehav. Rev. 2000, 24, 417–463. [Google Scholar] [CrossRef]
- Steinberg, L.; Albert, D.; Cauffman, E.; Banich, M.; Graham, S.; Woolard, J. Age differences in sensation seeking and impulsivity as indexed by behavior and self-report: Evidence for a dual systems model. Dev. Psychol. 2008, 44, 1764. [Google Scholar] [CrossRef] [PubMed]
- Spear, L.P. The adolescent brain and the college drinker: Biological basis of propensity to use and misuse alcohol. J. Stud. Alcoholsuppl. 2002, 41, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Novier, A.; Diaz-Granados, J.; Matthews, D. Alcohol use across the lifespan: An analysis of adolescent and aged rodents and humans. Pharmacol. Biochem. Behav. 2015, 133, 65–82. [Google Scholar] [CrossRef]
- Varlinskaya, E.I.; Spear, L.P. Acute ethanol withdrawal (hangover) and social behavior in adolescent and adult male and female Sprague-Dawley rats. Alcohol. Clin. Exp. Res. 2004, 28, 40–50. [Google Scholar] [CrossRef]
- Lee, K.M.; Coelho, M.A.; McGregor, H.A.; Solton, N.R.; Cohen, M.; Szumlinski, K.K. Adolescent Mice Are Resilient to Alcohol Withdrawal-Induced Anxiety and Changes in Indices of Glutamate Function within the Nucleus Accumbens. Front. Cell. Neurosci. 2016, 10, 265. [Google Scholar] [CrossRef]
- Lees, B.; Mewton, L.; Stapinski, L.A.; Squeglia, L.M.; Rae, C.D.; Teesson, M. Neurobiological and cognitive profile of young binge drinkers: A systematic review and meta-analysis. Neuropsychol. Rev. 2019, 29, 357–385. [Google Scholar] [CrossRef]
- Squeglia, L.M.; Jacobus, J.; Tapert, S.F. The effect of alcohol use on human adolescent brain structures and systems. Handb. Clin. Neurol. 2014, 125, 501–510. [Google Scholar]
- Conegundes, L.S.O.; Valente, J.Y.; Martins, C.B.; Andreoni, S.; Sanchez, Z.M. Binge drinking and frequent or heavy drinking among adolescents: Prevalence and associated factors. J. Pediatr. 2020, 2, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Kelly, A.; Chan, G.; Toumbourou, J.; Patton, G.; Williams, J. Beyond the primary influences of parents and peers on very young adolescent alcohol use. J. Early Adolesc. 2014, 34, 569–584. [Google Scholar] [CrossRef]
- Dukes, R.; Lorch, B. Concept of self, mediating factors, and adolescent deviance. Sociol. Spectr. 1989, 9, 301–319. [Google Scholar] [CrossRef]
- Brown, S.; Tapert, S. Adolescence and the trajectory of alcohol use: Basic to clinical studies. Ann. N. Y. Acad. Sci. 2004, 1021, 234–244. [Google Scholar] [CrossRef]
- Blakemore, S.J. The social brain in adolescence. Nat. Rev. Neurosci. 2008, 9, 267–277. [Google Scholar] [CrossRef]
- Ali, M.; Dwyer, D.; Vanner, E.; Lopez, A. Adolescent propensity to engage in health risky behaviors: The role of individual resilience. Int. J. Environ. Res. Public Health 2010, 7, 2161–2176. [Google Scholar] [CrossRef]
- Crews, F.T.; Braun, C.J.; Hoplight, B.; Switzer III, R.C.; Knapp, D.J. Binge ethanol consumption causes differential brain damage in young adolescent rats compared with adult rats. Alcohol. Clin. Exp. Res. 2000, 24, 1712–1723. [Google Scholar] [CrossRef]
- Aoki, M.; Shimozuru, M.; Kikusui, T.; Takeuchi, Y.; Mori, Y. Sex differences in behavioral and corticosterone responses to mild stressors in ICR mice are altered by ovariectomy in peripubertal period. Zool. Sci. 2010, 10, 783–789. [Google Scholar] [CrossRef]
- Torcaso, A.; Asimes, A.; Meagher, M.; Pak, T. Adolescent binge alcohol exposure increases risk assessment behaviors in male Wistar rats after exposure to an acute psychological stressor in adulthood. Psychoneuroendocrinology 2017, 76, 154–161. [Google Scholar] [CrossRef]
- Becker, H.C. Influence of stress associated with chronic alcohol exposure on drinking. Neuropharmacology 2017, 122, 115–126. [Google Scholar] [CrossRef]
- Koob, G.F. Neurocircuitry of alcohol addiction: Synthesis from animal models. Handb. Clin. Neurol. 2014, 125, 33–54. [Google Scholar] [PubMed]
- Lee, K.M.; Coehlo, M.A.; Solton, N.R.; Szumlinski, K.K. Negative affect and excessive alcohol intake incubate during protracted withdrawal from binge-drinking in adolescent, but not, adult mice. Front. Psychol. 2017, 8, 1128. [Google Scholar] [CrossRef]
- Lee, K.M.; Coelho, M.A.; Sern, K.R.; Szumlinski, K.K. Homer2 within the central nucleus of the amygdala gates withdrawal-induced anxiety in a mouse model of binge-drinking. Neuropharmacology 2018, 128, 448–459. [Google Scholar] [CrossRef]
- Lee, K.M.; Coelho, M.A.; Class, M.A.; Sern, K.R.; Bocz, M.D.; Szumlinski, K.K. mGlu5 blockade within the nucleus accumbens shell reduces alcohol withdrawal-induced anxiety in adult and adolescent mice. Front. Pharmacol. 2018, 9, 1306. [Google Scholar] [CrossRef]
- World Health Organization. Management of Substance Abuse Unit. Global Status Report on Alcohol and Health, 2014; World Health Organization Press: Geneva, Switzerland, 2014. [Google Scholar]
- Wilsnack, R.W.; Wilsnack, S.C.; Gmel, G.; Kantor, L.W. Gender differences in binge drinking. Alcohol Res. 2018, 39, 57. [Google Scholar]
- Agabio, R.; Pisanu, C.; Gessa, G.L.; Franconi, F. Sex differences in alcohol use disorder. Curr. Med. Chem. 2017, 24, 2661–2670. [Google Scholar] [CrossRef]
- Becker, J.B.; McClellan, M.L.; Reed, B.G. Sex differences, gender and addiction. J. Neurosci. Res. 2017, 95, 136–147. [Google Scholar] [CrossRef]
- Finn, D.A.; Beckley, E.H.; Kaufman, K.R.; Ford, M.M. Manipulation of GABAergic steroids: Sex differences in the effects on alcohol drinking- and withdrawal-related behaviors. Horm. Behav. 2010, 57, 12–22. [Google Scholar] [CrossRef]
- Logrip, M.L.; Milivojevic, V.; Bertholomey, M.L.; Torregrossa, M.M. Sexual dimorphism in the neural impact of stress and alcohol. Alcohol 2018, 72, 49–59. [Google Scholar] [CrossRef]
- Strong, M.N.; Yoneyama, N.; Fretwell, A.M.; Snelling, C.; Tanchuck, M.A.; Finn, D.A. Binge drinking experience in adolescent mice shows sex differences and elevated ethanol intake in adulthood. Horm. Behav. 2010, 58, 82–90. [Google Scholar] [CrossRef]
- Cozzoli, D.K.; Strong-Kaufman, M.N.; Tanchuck, M.A.; Hashimoto, J.G.; Wiren, K.M.; Finn, D.A. The effect of mGluR5 antagonism during binge drinking on subsequent ethanol intake in C57BL/6J mice: Sex- and age-induced differences. Alcohol. Clin. Exp. Res. 2014, 38, 730–738. [Google Scholar] [CrossRef]
- Szumlinski, K.K.; Coelho, M.A.; Lee, K.M.; Tran, T.; Sern, K.; Bernal, A.; Kippin, T.E. DID it or DIDn’t it? Exploration of a failure to replicate binge-like alcohol-drinking in C57BL/6J mice. Pharmacol. Biochem. Behav. 2019, 178, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Melón, L.C.; Wray, K.N.; Moore, E.M.; Boehm II, S.L. Sex and age differences in heavy binge drinking and its effects on alcohol responsivity following abstinence. Pharmacol. Biochem. Behav. 2013, 104, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Moore, E.M.; Mariani, J.N.; Linsenbardt, D.N.; Melón, L.C.; Boehm II, S.L. Adolescent C57BL/6J (but not DBA/2J) mice consume greater amounts of limited-access ethanol compared to adults and display continued elevated ethanol intake into adulthood. Alcohol. Clin. Exp. Res. 2010, 34, 732–742. [Google Scholar] [CrossRef]
- Wellman, C.L.; Bangasser, D.A.; Bollinger, J.L.; Coutellier, L.; Logrip, M.L.; Moench, K.M.; Urban, K.R. Sex differences in risk and resilience: Stress effects on the neural substrates of emotion and motivation. J. Neurosci. 2018, 38, 9423–9432. [Google Scholar] [CrossRef]
- Romeo, R.D.; Patel, R.; Pham, L.; So, V.M. Adolescence and the ontogeny of the hormonal stress response in male and female rats and mice. Neurosci. Biobehav. Rev. 2016, 70, 206–216. [Google Scholar] [CrossRef]
- Romeo, R.D. The metamorphosis of adolescent hormonal stress reactivity: A focus on animal models. Front. Neuroendocrinol. 2018, 49, 43–51. [Google Scholar] [CrossRef]
- Lee, K.M.; Coelho, M.A.; Sern, K.R.; Class, M.A.; Bocz, M.D.; Szumlinski, K.K. Anxiolytic effects of buspirone and MTEP in the Porsolt Forced Swim Test. Chronic Stress 2017, 1, 2470547017712985. [Google Scholar] [CrossRef]
- Lee, K.M.; Coelho, M.A.; Class, M.A.; Szumlinski, K.K. mGlu5-dependent modulation of anxiety during early withdrawal from binge-drinking in adult and adolescent male mice. Drug Alcohol Depend. 2018, 184, 1–11. [Google Scholar] [CrossRef]
- Lee, K.M.; Coehlo, M.; McGregor, H.A.; Waltermire, R.S.; Szumlinski, K.K. Binge alcohol drinking elicits persistent negative affect in mice. Behav. Brain Res. 2015, 291, 385–398. [Google Scholar] [CrossRef]
- Tiscione, N.; Alford, I.; Yeatman, D.; Shan, X. Ethanol analysis by headspace gas chromatography with simultaneous flame-ionization and mass spectrometry detection. J. Anal. Toxicol. 2011, 35, 501–511. [Google Scholar] [CrossRef] [PubMed]
- McCall, N.M.; Sprow, G.M.; Delpire, E.; Thiele, T.E.; Kash, T.L.; Pleil, K.E. Effects of sex and deletion of neuropeptide Y2 receptors from GABAergic neurons on affective and alcohol drinking behaviors in mice. Front. Integr. Neurosci. 2013, 7, 100. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, L.H.; Bettler, B.; Kaupmann, K.; Cryan, J.F. Behavioral evaluation of mice deficient in GABA B (1) receptor isoforms in tests of unconditioned anxiety. Psychopharmacology 2007, 190, 541–553. [Google Scholar] [CrossRef][Green Version]
- Crawley, J. Exploratory behavior models of anxiety in mice. Neurosci. Biobehav. Rev. 1985, 9, 37–44. [Google Scholar] [CrossRef]
- Gallo, I.; Rattazzi, L.; Piras, G.; Gobbetti, T.; Panza, E.; Perretti, M.; D’Acquisto, F. Formyl peptide receptor as a novel therapeutic target for anxiety-related disorders. PLoS ONE 2004, 9, e114626. [Google Scholar] [CrossRef]
- Porsolt, R.D.; Brossard, G.; Hautbois, C.; Roux, S. Rodent models of depression: Forced swimming and tail suspension behavioral despair tests in rats and mice. Curr. Protoc. Neurosci. 2001, 14, 8–10. [Google Scholar] [CrossRef]
- Young, R.; Batkai, S.; Dukat, M.; Glennon, R.A. TDIQ (5,6,7,8-tetrahydro-1,3-dioxolo[4,5-g]isoquinoline) exhibits anxiolytic-like activity in a marble-burying assay in mice. Pharmacol. Biochem. Behav. 2006, 84, 62–73. [Google Scholar] [CrossRef]
- Romeo, R.; Parfitt, D.; Richardson, H.; Sisk, C. Pheromones elicit equivalent levels of fos-Immunoreactivity in prepubertal and adult male syrian hamsters. Horm. Behav. 1998, 34, 48–55. [Google Scholar] [CrossRef]
- Bell, M.; Lorme, K.; Figueira, R.; Kashy, D.; Sisk, C. Adolescent gain in positive valence of a socially relevant stimulus: Engagement of the mesocorticolimbic reward circuitry. Eur. J. Neurosci. 2013, 37, 457–468. [Google Scholar] [CrossRef]
- Bell, M.R.; Meerts, S.H.; Sisk, C.L. Adolescent brain maturation is necessary for adult-typical mesocorticolimbic responses to a rewarding social cue. Dev. Neurobiol. 2013, 73, 856–869. [Google Scholar] [CrossRef]
- Aikey, J.L.; Nyby, J.G.; Anmuth, D.M.; James, P.J. Testosterone rapidly reduces anxiety in male house mice (Mus musculus). Horm. Behav. 2002, 42, 448–460. [Google Scholar] [CrossRef]
- Fernández-Guasti, A.; Martínez-Mota, L. Anxiolytic-like actions of testosterone in the burying behavior test: Role of androgen and GABA-benzodiazepine receptors. Psychoneuroendocrinology 2005, 30, 762–770. [Google Scholar] [CrossRef]
- Frye, C.A.; Koonce, C.J.; Edinger, K.L.; Osborne, D.M.; Walf, A.A. Androgens with activity at estrogen receptor beta have anxiolytic and cognitive-enhancing effects in male rats and mice. Horm. Behav. 2008, 54, 726–734. [Google Scholar] [CrossRef]
- Fernandez-Guasti, A.; Picazo, O. Changes in burying behavior during the estrous cycle: Effect of estrogen and progesterone. Psychoneuroendocrinology 1992, 17, 681–689. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drinking History | Dependent Variable | Basal CORT | Stressor-Induced CORT |
|---|---|---|---|
| Water (N = 81) | Latency to Bury | r = 0.155, p = 0.168 | r = −0.137, p = 0.223 |
| Time in light | r = 0.065, p = 0.562 | r = 0.041, p = 0.717 | |
| Light Entries | r = 0.229, p = 0.039 | r = 0.011, p = 0.921 | |
| Latency to Bury | r = 0.155, p = 0.168 | r = −0.137, p = 0.223 | |
| Time Spent Burying | r = 0.068, p = 0.547 | r = −0.111, p = 0.325 | |
| Percent Buried | r = −0.069, p = 0.541 | r = 0.077, p = 0.493 | |
| Latency to Immobility | r = 0.049, p = 0.667 | r = −0.082, p = 0.466 | |
| Time Spent Immobile | r = −0.042, p = 0.711 | r = 0.067, p = 0.552 | |
| Immobile Episodes | r = −0.092, p = 0.412 | r = 0.076, p = 0.501 | |
| Alcohol (N = 98) | Latency to enter light | r = −0.107, p = 0.296 | r = 0.025, p = 0.807 |
| Time in light | r = −0.133, p = 0.193 | r = 0.116, p = 0.254 | |
| Light Entries | r = 0.203, p = 0.045 | r = 0.018, p = 0.860 | |
| Latency to Bury | r = −0.216, p = 0.033 | r =0.354, p < 0.001 | |
| Time Spent Burying | r = 0.397, p< 0.001 | r = −0.262, p = 0.009 | |
| Percent Buried | r = 0.216, p = 0.033 | r = −0.171, p = 0.093 | |
| Latency to Immobility | r = −0.005, p = 0.962 | r = −0.003, p = 0.974 | |
| Time Spent Immobile | r = 0.031, p = 0.763 | r = 0.077, p = 0.448 | |
| Immobile Episodes | r = 0.084, p = 0.410 | r = 0.059, p = 0.561 |
| Subject Factor | Measure | Latency to Bury | Time Burying | % Buried |
|---|---|---|---|---|
| Females (N = 49) | Ave. Intake | r = 0.313, p = 0.028 | r = −0.204, p = 0.160 | r = −0.442, p = 0.001 |
| Escalation | r = 0.327, p = 0.023 | r = −0.309, p = 0.033 | r = −0.373, p = 0.009 | |
| Males (N = 50) | Ave. Intake | r = 0.224, p = 0.119 | r = −0.146, p = 0.313 | r = −0.113, p = 0.435 |
| Escalation | r = −0.136, p = 0.361 | r = 0.298, p = 0.042 | r = 0.372, p = 0.010 | |
| Adolescents (N = 46) | Ave. Intake | r = −0.117, p = 0.439 | r = −0.141, p = 0.350 | r = −0.145, p = 0.335 |
| Escalation | r = −0.123, p = 0.415 | r = 0.135, p = 0.371 | r = 0.150, p = 0.319 | |
| Adults (N = 53) | Ave. Intake | r = 0.260, p = 0.060 | r = −0.023, p = 0.873 | r = 0.015, p = 0.915 |
| Escalation | r = 0.271, p = 0.060 | r = −0.106, p = 0.272 | r = −0.215, p = 0.139 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jimenez Chavez, C.L.; Coelho, M.A.; Brewin, L.W.; Swauncy, I.; Tran, T.; Albanese, T.; Laguna, A.; Gabriela, I.; Szumlinski, K.K. Incubation of Negative Affect during Protracted Alcohol Withdrawal Is Age-, but Not Sex-Selective. Brain Sci. 2020, 10, 405. https://doi.org/10.3390/brainsci10060405
Jimenez Chavez CL, Coelho MA, Brewin LW, Swauncy I, Tran T, Albanese T, Laguna A, Gabriela I, Szumlinski KK. Incubation of Negative Affect during Protracted Alcohol Withdrawal Is Age-, but Not Sex-Selective. Brain Sciences. 2020; 10(6):405. https://doi.org/10.3390/brainsci10060405
Chicago/Turabian StyleJimenez Chavez, C. Leonardo, Michal A. Coelho, Lindsey W. Brewin, Isaiah Swauncy, Tori Tran, Taylor Albanese, Angie Laguna, Ivette Gabriela, and Karen K. Szumlinski. 2020. "Incubation of Negative Affect during Protracted Alcohol Withdrawal Is Age-, but Not Sex-Selective" Brain Sciences 10, no. 6: 405. https://doi.org/10.3390/brainsci10060405
APA StyleJimenez Chavez, C. L., Coelho, M. A., Brewin, L. W., Swauncy, I., Tran, T., Albanese, T., Laguna, A., Gabriela, I., & Szumlinski, K. K. (2020). Incubation of Negative Affect during Protracted Alcohol Withdrawal Is Age-, but Not Sex-Selective. Brain Sciences, 10(6), 405. https://doi.org/10.3390/brainsci10060405