Postnatal Guinea Pig Brain Development, as Revealed by Magnetic Resonance and Diffusion Kurtosis Imaging

Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Model
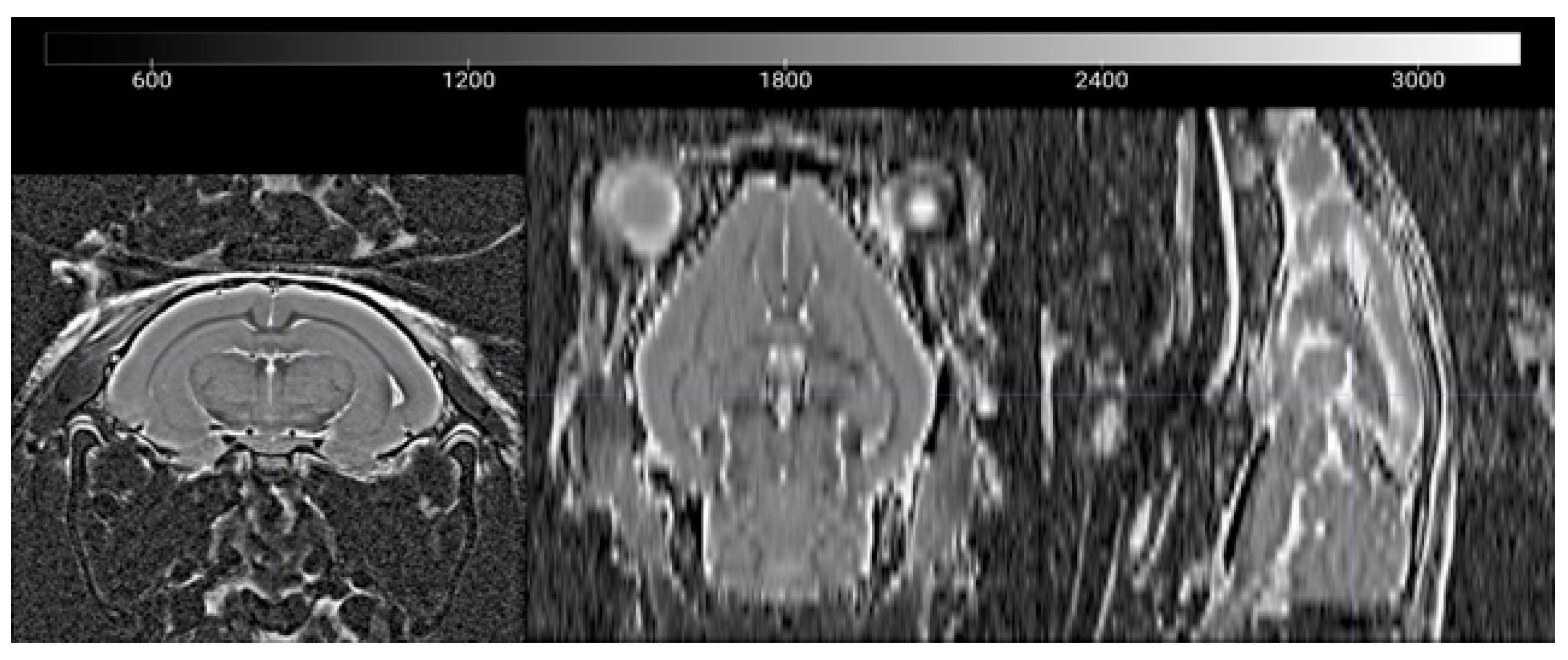
2.2. In Vivo MRI
2.3. Data Processing
2.4. Statistical Analysis
3. Results
3.1. Brain Growth during Postnatal Maturation
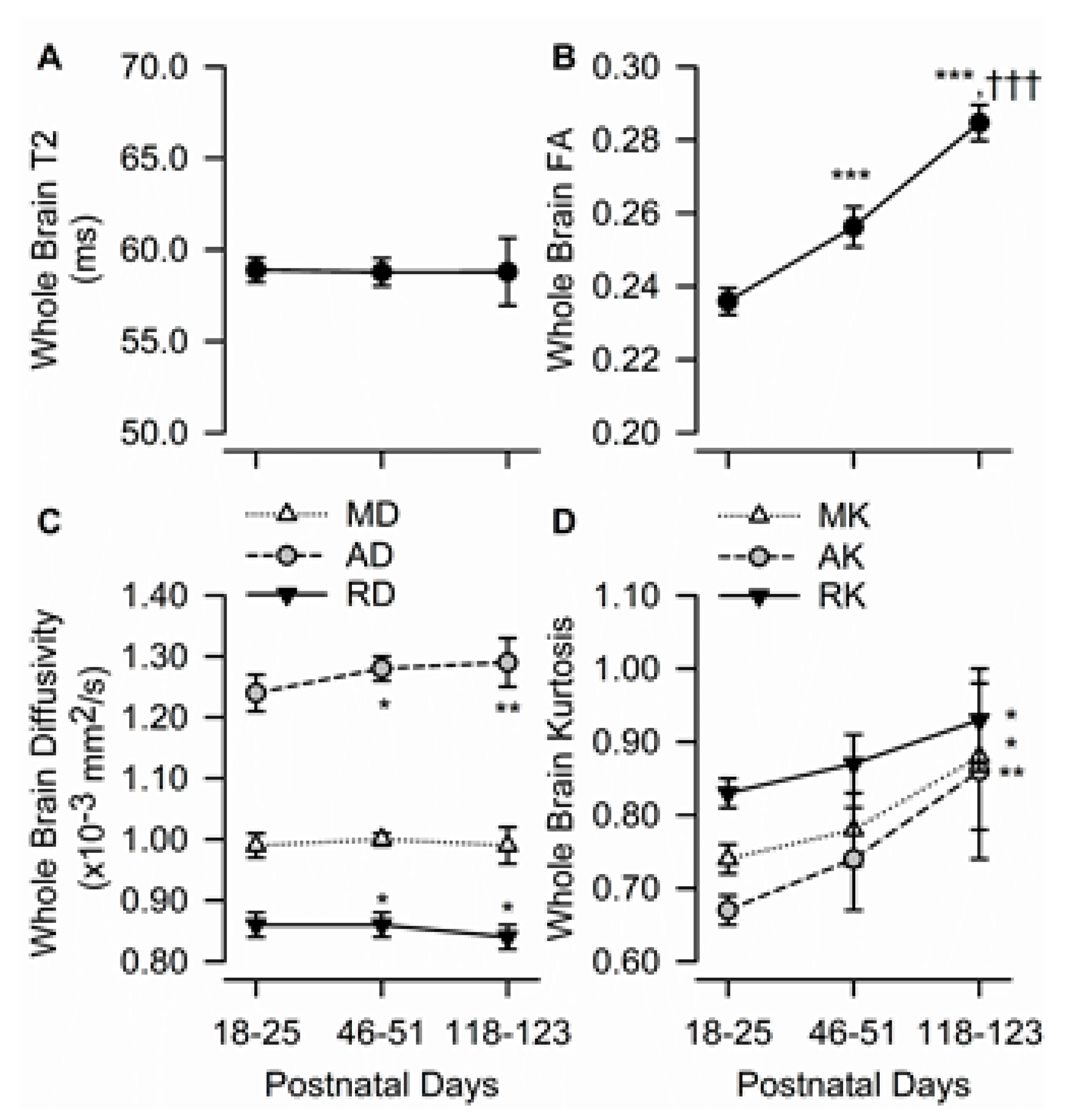
3.2. Age Dependency of Whole Brain T2, FA, Diffusivity, and Kurtosis Measures
3.3. Age Dependence of Regional FA
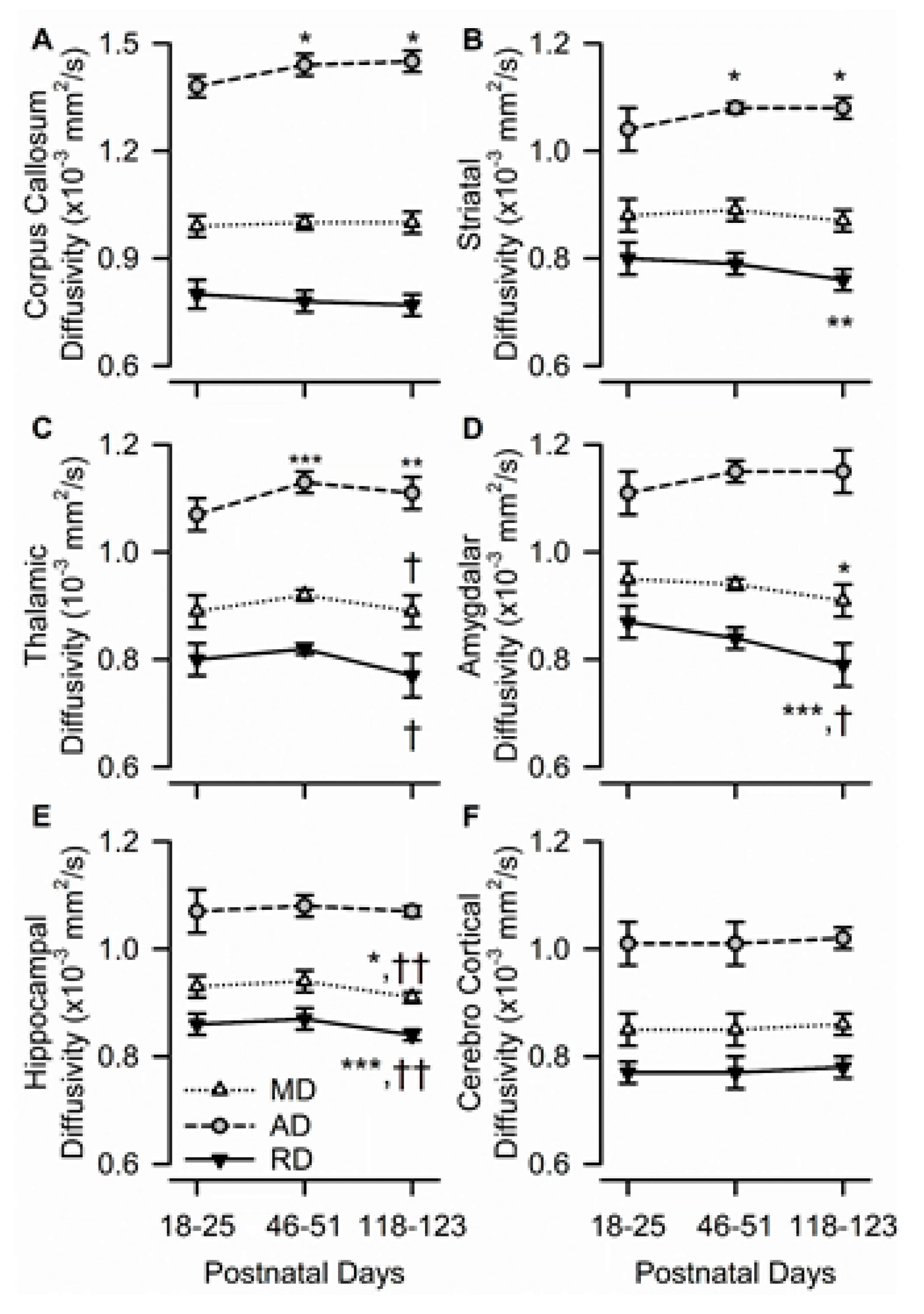
3.4. Age Dependence of Regional Diffusivity Measures
3.5. Age Dependence of Regional Kurtosis Measures
3.6. Age Dependence of Regional T2 Relaxation Time
4. Discussion
Author Contributions
Funding
Conflicts of Interest
Appendix A


References
- Burke, R.D.; Todd, S.W.; Lumsden, E.; Mullins, R.J.; Mamczarz, J.; Fawcett, W.P.; Gullapalli, R.P.; Randall, W.R.; Pereira, E.F.R.; Albuquerque, E.X. Developmental neurotoxicity of the organophosphorus insecticide chlorpyrifos: From clinical findings to preclinical models and potential mechanisms. J. Neurochem. 2017, 142, 162–177. [Google Scholar] [CrossRef]
- Morrison, J.L.; Botting, K.J.; Darby, J.R.T.; David, A.L.; Dyson, R.M.; Gatford, K.L.; Gray, C.; Herrera, E.A.; Hirst, J.J.; Kim, B.; et al. Guinea pig models for translation of the developmental origins of health and disease hypothesis into the clinic. J. Physiol. 2018, 596, 5535–5569. [Google Scholar] [CrossRef]
- Salazar, C.; Valdivia, G.; Ardiles, Á.O.; Ewer, J.; Palacios, A.G. Genetic variants associated with neurodegenerative Alzheimer disease in natural models. Biol. Res. 2016, 49, 14. [Google Scholar] [CrossRef]
- Dobbing, J.; Sands, J. Quantitative growth and development of human brain. Arch. Dis. Child. 1973, 48, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Dobbing, J.; Sands, J. Growth and development of the brain and spinal cord of the guinea pig. Brain Res. 1970, 17, 115–123. [Google Scholar] [CrossRef]
- Ecobichon, D.J.; Dykeman, R.W.; Hansell, M.M. The development of hepatic drug-metabolizing enzymes in perinatal guinea pigs: A biochemical and morphological study. Can. J. Biochem. 1978, 56, 738–745. [Google Scholar] [CrossRef]
- Beck, M.; Müller, D.; Bigl, V. Amyloid precursor protein in guinea pigs--complete cDNA sequence and alternative splicing. Biochim. Biophys. Acta 1997, 1351, 17–21. [Google Scholar] [CrossRef]
- Johnstone, E.M.; Chaney, M.O.; Norris, F.H.; Pascual, R.; Little, S.P. Conservation of the sequence of the Alzheimer’s disease amyloid peptide in dog, polar bear and five other mammals by cross-species polymerase chain reaction analysis. Brain Res. Mol. Brain Res. 1991, 10, 299–305. [Google Scholar] [CrossRef]
- Sharman, M.J.; Moussavi Nik, S.H.; Chen, M.M.; Ong, D.; Wijaya, L.; Laws, S.M.; Taddei, K.; Newman, M.; Lardelli, M.; Martins, R.N.; et al. The Guinea Pig as a Model for Sporadic Alzheimer’s Disease (AD): The Impact of Cholesterol Intake on Expression of AD-Related Genes. PLoS ONE 2013, 8, e66235. [Google Scholar] [CrossRef] [PubMed]
- Potter, G.; Brueck, W. Nervous System of Guinea Pig. Bios 1958, 29, 185–196. [Google Scholar]
- Rapisarda, C.; Bacchelli, B. The brain of the guinea pig in stereotaxic coordinates. Arch. Sci. Biol. (Bologna) 1977, 61, 1–37. [Google Scholar]
- Tindal, J.S. The forebrain of the guinea pig in stereotaxic coordinates. J. Comp. Neurol. 1965, 124, 259–266. [Google Scholar] [CrossRef]
- Cheung, M.M.; Hui, E.S.; Chan, K.C.; Helpern, J.A.; Qi, L.; Wu, E.X. Does diffusion kurtosis imaging lead to better neural tissue characterization? A rodent brain maturation study. Neuroimage 2009, 45, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Basser, P.J. Inferring microstructural features and the physiological state of tissues from diffusion-weighted images. NMR Biomed. 1995, 8, 333–344. [Google Scholar] [CrossRef]
- Saunders, M.; Magnanti, B.L.; Correia Carreira, S.; Yang, A.; Alamo-Hernández, U.; Riojas-Rodriguez, H.; Calamandrei, G.; Koppe, J.G.; Krayer von Krauss, M.; Keune, H.; et al. Chlorpyrifos and neurodevelopmental effects: A literature review and expert elicitation on research and policy. Environ. Health 2012, 11 (Suppl. 1), S5. [Google Scholar] [CrossRef]
- Hui, E.S.; Cheung, M.M.; Qi, L.; Wu, E.X. Towards better MR characterization of neural tissues using directional diffusion kurtosis analysis. Neuroimage 2008, 42, 122–134. [Google Scholar] [CrossRef]
- Zhuo, J.; Xu, S.; Proctor, J.L.; Mullins, R.J.; Simon, J.Z.; Fiskum, G.; Gullapalli, R.P. Diffusion kurtosis as an in vivo imaging marker for reactive astrogliosis in traumatic brain injury. Neuroimage 2012, 59, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Zhuo, J.; Racz, J.; Shi, D.; Roys, S.; Fiskum, G.; Gullapalli, R. Early microstructural and metabolic changes following controlled cortical impact injury in rat: A magnetic resonance imaging and spectroscopy study. J. Neurotrauma 2011, 28, 2091–2102. [Google Scholar] [CrossRef] [PubMed]
- Arab, A.; Ruda-Kucerova, J.; Minsterova, A.; Drazanova, E.; Szabó, N.; Starcuk, Z.; Rektorova, I.; Khairnar, A. Diffusion Kurtosis Imaging Detects Microstructural Changes in a Methamphetamine-Induced Mouse Model of Parkinson’s Disease. Neurotox. Res. 2019, 36, 724–735. [Google Scholar] [CrossRef] [PubMed]
- Khairnar, A.; Ruda-Kucerova, J.; Szabó, N.; Drazanova, E.; Arab, A.; Hutter-Paier, B.; Neddens, J.; Latta, P.; Starcuk, Z.; Rektorova, I. Early and progressive microstructural brain changes in mice overexpressing human α-Synuclein detected by diffusion kurtosis imaging. Brain Behav. Immun. 2017, 61, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zeng, J.; Shen, Z.-W.; Kong, L.; Zheng, W. Diffusion Kurtosis Imaging Detects Microstructural Changes in the Brain after Acute Alcohol Intoxication in Rats. Biomed Res. Int. 2017, 2017, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Shukla, D.K.; Armstrong, R.C.; Marion, C.M.; Radomski, K.L.; Selwyn, R.G.; Dardzinski, B.J. Repetitive Model of Mild Traumatic Brain Injury Produces Cortical Abnormalities Detectable by Magnetic Resonance Diffusion Imaging, Histopathology, and Behavior. J. Neurotrauma 2017, 34, 1364–1381. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-L.; Yu, M.-M.; Yang, D.-X.; Liu, Y.-L.; Wei, X.-E.; Li, W.-B. Longitudinal Microstructural Changes in Traumatic Brain Injury in Rats: A Diffusional Kurtosis Imaging, Histology, and Behavior Study. Am. J. Neuroradiol. 2018, 39, 1650–1656. [Google Scholar] [CrossRef]
- Wang, J.-J.; Lin, W.-Y.; Lu, C.-S.; Weng, Y.-H.; Ng, S.-H.; Wang, C.-H.; Liu, H.-L.; Hsieh, R.-H.; Wan, Y.-L.; Wai, Y.-Y. Parkinson Disease: Diagnostic Utility of Diffusion Kurtosis Imaging. Radiology 2011, 261, 210–217. [Google Scholar] [CrossRef]
- Blockx, I.; Verhoye, M.; Van Audekerke, J.; Bergwerf, I.; Kane, J.X.; Delgado y Palacios, R.; Veraart, J.; Jeurissen, B.; Raber, K.; von Hörsten, S.; et al. Identification and characterization of Huntington related pathology: An in vivo DKI imaging study. Neuroimage 2012, 63, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Gong, N.-J.; Wong, C.-S.; Chan, C.-C.; Leung, L.-M.; Chu, Y.-C. Correlations between microstructural alterations and severity of cognitive deficiency in Alzheimer’s disease and mild cognitive impairment: A diffusional kurtosis imaging study. Magn. Reson. Imaging 2013, 31, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Zhang, Z.; Wen, C.; Liu, H.; Wang, S.; Li, J.; Zhuge, Q.; Chen, W.; Ye, Q. Characterization of Alzheimer’s Disease Using Ultra-high b-values Apparent Diffusion Coefficient and Diffusion Kurtosis Imaging. Aging Dis. 2019, 10, 1026. [Google Scholar] [CrossRef]
- Paydar, A.; Fieremans, E.; Nwankwo, J.I.; Lazar, M.; Sheth, H.D.; Adisetiyo, V.; Helpern, J.A.; Jensen, J.H.; Milla, S.S. Diffusional Kurtosis Imaging of the Developing Brain. Am. J. Neuroradiol. 2014, 35, 808–814. [Google Scholar] [CrossRef]
- Ferrie, J.C.; Barantin, L.; Saliba, E.; Akoka, S.; Tranquart, F.; Sirinelli, D.; Pourcelot, L. MR assessment of the brain maturation during the perinatal period: Quantitative T2 MR study in premature newborns. Magn. Reson. Imaging 1999, 17, 1275–1288. [Google Scholar] [CrossRef]
- Wender, M.; Hierowski, M. The Concentration of electrolytes in the developing nervous system with special reference to the period of myelination. J. Neurochem. 1960, 5, 105–108. [Google Scholar] [CrossRef]
- Holland, B.A.; Haas, D.K.; Norman, D.; Brant-Zawadzki, M.; Newton, T.H. MRI of normal brain maturation. AJNR. Am. J. Neuroradiol. 1986, 7, 201–208. [Google Scholar] [PubMed]
- Leppert, I.R.; Almli, C.R.; McKinstry, R.C.; Mulkern, R.V.; Pierpaoli, C.; Rivkin, M.J.; Pike, G.B. T(2) relaxometry of normal pediatric brain development. J. Magn. Reson. Imaging 2009, 29, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Bartesaghi, R.; Guidi, S.; Severi, S.; Contestabile, A.; Ciani, E. Sex differences in the hippocampal dentate gyrus of the guinea-pig before puberty. Neuroscience 2003, 121, 327–339. [Google Scholar] [CrossRef]
- Wolfer, D.P.; Lipp, H.P. Evidence for physiological growth of hippocampal mossy fiber collaterals in the guinea pig during puberty and adulthood. Hippocampus 1995, 5, 329–340. [Google Scholar] [CrossRef]
- Hennessy, M.B.; Hornschuh, G.; Kaiser, S.; Sachser, N. Cortisol responses and social buffering: A study throughout the life span. Horm. Behav. 2006, 49, 383–390. [Google Scholar] [CrossRef]
- McAuliffe, M.J.; Lalonde, F.M.; McGarry, D.; Gandler, W.; Csaky, K.; Trus, B.L. Medical Image Processing, Analysis and Visualization in clinical research. In Proceedings of the 14th IEEE Symposium on Computer-Based Medical Systems, CBMS, Bethesda, MD, USA, 26–27 July 2001; pp. 381–386. [Google Scholar]
- Altman, J.; Das, G.D. Postnatal neurogenesis in the guinea-pig. Nature 1967, 214, 1098–1101. [Google Scholar] [CrossRef]
- Slotnick, B.M.; Leonard, C.M. Stereotaxic atlas of the albino mouse forebrain. Ann. Neurol. 1981, 10, 403. [Google Scholar]
- Pfefferbaum, A.; Mathalon, D.H.; Sullivan, E.V.; Rawles, J.M.; Zipursky, R.B.; Lim, K.O. A quantitative magnetic resonance imaging study of changes in brain morphology from infancy to late adulthood. Arch. Neurol. 1994, 51, 874–887. [Google Scholar] [CrossRef]
- Bockhorst, K.H.; Narayana, P.A.; Liu, R.; Ahobila-Vijjula, P.; Ramu, J.; Kamel, M.; Wosik, J.; Bockhorst, T.; Hahn, K.; Hasan, K.M.; et al. Early postnatal development of rat brain: In vivo diffusion tensor imaging. J. Neurosci. Res. 2008, 86, 1520–1528. [Google Scholar] [CrossRef]
- Calabrese, E.; Johnson, G.A. Diffusion tensor magnetic resonance histology reveals microstructural changes in the developing rat brain. Neuroimage 2013, 79, 329–339. [Google Scholar] [CrossRef]
- Hüppi, P.S.; Dubois, J. Diffusion tensor imaging of brain development. Semin. Fetal Neonatal Med. 2006, 11, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Larvaron, P.; Bielicki, G.; Boespflug-Tanguy, O.; Renou, J.-P. Proton MRS of early post-natal mouse brain modifications in vivo. NMR Biomed. 2006, 19, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.; Miller, J.H.; Shimony, J.S.; Philip, J.V.; Nehra, D.; Snyder, A.Z.; Conturo, T.E.; Neil, J.J.; McKinstry, R.C. Diffusion-tensor MR imaging of gray and white matter development during normal human brain maturation. AJNR Am. J. Neuroradiol. 2002, 23, 1445–1456. [Google Scholar] [PubMed]
- Jones, D.G.; Dittmer, M.M.; Reading, L.C. Synaptogenesis in guinea-pig cerebral cortex: A glutaral-dehyde-PTA study. Brain Res. 1974, 70, 245–259. [Google Scholar] [CrossRef]
- Steven, A.J.; Zhuo, J.; Melhem, E.R. Diffusion kurtosis imaging: An emerging technique for evaluating the microstructural environment of the brain. AJR Am. J. Roentgenol. 2014, 202, W26–W33. [Google Scholar] [CrossRef] [PubMed]
- Brownson, R.H. The effect of x-irradiation on the perineuronal satellite cells in the cortex of aging brains. J. Neuropathol. Exp. Neurol. 1960, 19, 407–414. [Google Scholar] [CrossRef]
- Schüz, A. Comparison between the dimensions of dendritic spines in the cerebral cortex of newborn and adult guinea pigs. J. Comp. Neurol. 1986, 244, 277–285. [Google Scholar] [CrossRef]
- Mathur-De Vré, R. Biomedical implications of the relaxation behaviour of water related to NMR imaging. Br. J. Radiol. 1984, 57, 955–976. [Google Scholar] [CrossRef] [PubMed]
- Masumura, M. Proton relaxation time of immature brain. II. In vivo measurement of proton relaxation time (T1 and T2) in pediatric brain by MRI. Childs Nerv. Syst. 1987, 3, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Irfanoglu, M.O.; Modi, P.; Nayak, A.; Hutchinson, E.B.; Sarlls, J.; Pierpaoli, C. DR-BUDDI (Diffeomorphic Registration for Blip-Up blip-Down Diffusion Imaging) method for correcting echo planar imaging distortions. Neuroimage 2015, 106, 284–299. [Google Scholar] [CrossRef]
- Culley, D.J.; Baxter, M.G.; Yukhananov, R.; Crosby, G. Long-term impairment of acquisition of a spatial memory task following isoflurane-nitrous oxide anesthesia in rats. Anesthesiology 2004, 100, 309–314. [Google Scholar] [CrossRef]
- Jevtovic-Todorovic, V. Exposure of Developing Brain to General Anesthesia. Anesthesiology 2018, 128, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Rauh, V.A.; Perera, F.P.; Horton, M.K.; Whyatt, R.M.; Bansal, R.; Hao, X.; Liu, J.; Barr, D.B.; Slotkin, T.A.; Peterson, B.S. Brain anomalies in children exposed prenatally to a common organophosphate pesticide. Proc. Natl. Acad. Sci. USA 2012, 109, 7871–7876. [Google Scholar] [CrossRef]
- Struyfs, H.; Van Hecke, W.; Veraart, J.; Sijbers, J.; Slaets, S.; De Belder, M.; Wuyts, L.; Peters, B.; Sleegers, K.; Robberecht, C.; et al. Diffusion Kurtosis Imaging: A Possible MRI Biomarker for AD Diagnosis? J. Alzheimers Dis. 2015, 48, 937–948. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measures | Juvenile | Adolescent | Adult | |
|---|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | ANOVA p | |
| Body Weight (g) | 230.9 ± 26.5 | 435.3 ± 25.3 | 806.9 ± 36.9 | <0.001 |
| Volume (mm3) | ||||
| Intracranial | 3512.4 ± 164.0 | 4045.8 ± 163.8 | 4530.8 ± 133.5 | <0.001 |
| Parenchyma | 3347.7 ± 153.6 | 3829.7 ± 159.1 | 4282.9 ± 141.4 | <0.001 |
| Cerebrospinal Fluid (CSF) | 164.7 ± 31.7 | 216.1 ± 28.8 | 247.9 ± 17.2 | <0.001 |
| Brain Length (mm) | 33.5 ± 1.2 | 36.5 ± 0.5 | 38.5 ± 0.8 | <0.001 |
| Brain Width (mm) | 20.4 ± 0.4 | 21.4 ± 0.3 | 22.1 ± 0.4 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mullins, R.J.; Xu, S.; Zhuo, J.; Roys, S.; Pereira, E.F.R.; Albuquerque, E.X.; Gullapalli, R.P. Postnatal Guinea Pig Brain Development, as Revealed by Magnetic Resonance and Diffusion Kurtosis Imaging. Brain Sci. 2020, 10, 365. https://doi.org/10.3390/brainsci10060365
Mullins RJ, Xu S, Zhuo J, Roys S, Pereira EFR, Albuquerque EX, Gullapalli RP. Postnatal Guinea Pig Brain Development, as Revealed by Magnetic Resonance and Diffusion Kurtosis Imaging. Brain Sciences. 2020; 10(6):365. https://doi.org/10.3390/brainsci10060365
Chicago/Turabian StyleMullins, Roger J., Su Xu, Jiachen Zhuo, Steve Roys, Edna F.R. Pereira, Edson X. Albuquerque, and Rao P. Gullapalli. 2020. "Postnatal Guinea Pig Brain Development, as Revealed by Magnetic Resonance and Diffusion Kurtosis Imaging" Brain Sciences 10, no. 6: 365. https://doi.org/10.3390/brainsci10060365
APA StyleMullins, R. J., Xu, S., Zhuo, J., Roys, S., Pereira, E. F. R., Albuquerque, E. X., & Gullapalli, R. P. (2020). Postnatal Guinea Pig Brain Development, as Revealed by Magnetic Resonance and Diffusion Kurtosis Imaging. Brain Sciences, 10(6), 365. https://doi.org/10.3390/brainsci10060365