Event-Related Potentials (ERP) Indices of Motivation during the Effort Expenditure for Reward Task

 , , and
, , and
Abstract
1. Introduction
1.1. Previous Neuroimaging Studies on Motivation
1.2. Evaluation Methods
1.3. Study Rationale
2. Methods
2.1. Participants
2.2. Experimental Task and Measurements
2.3. Data Acquisition
2.4. Data Analysis
2.5. Source Imaging
3. Results
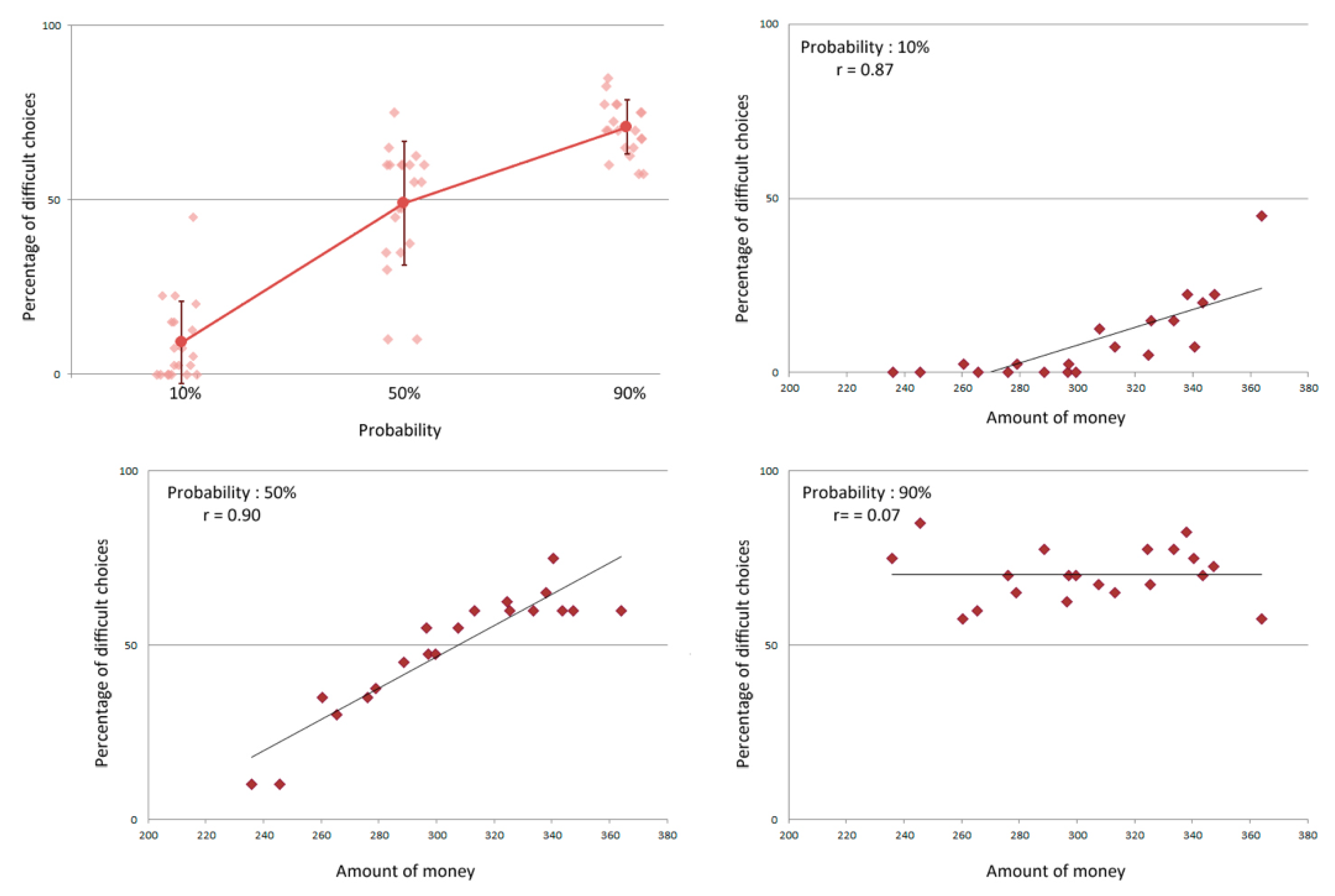
3.1. Behavioral Results on the EEfRT
3.2. ERP Results on the EEfRT
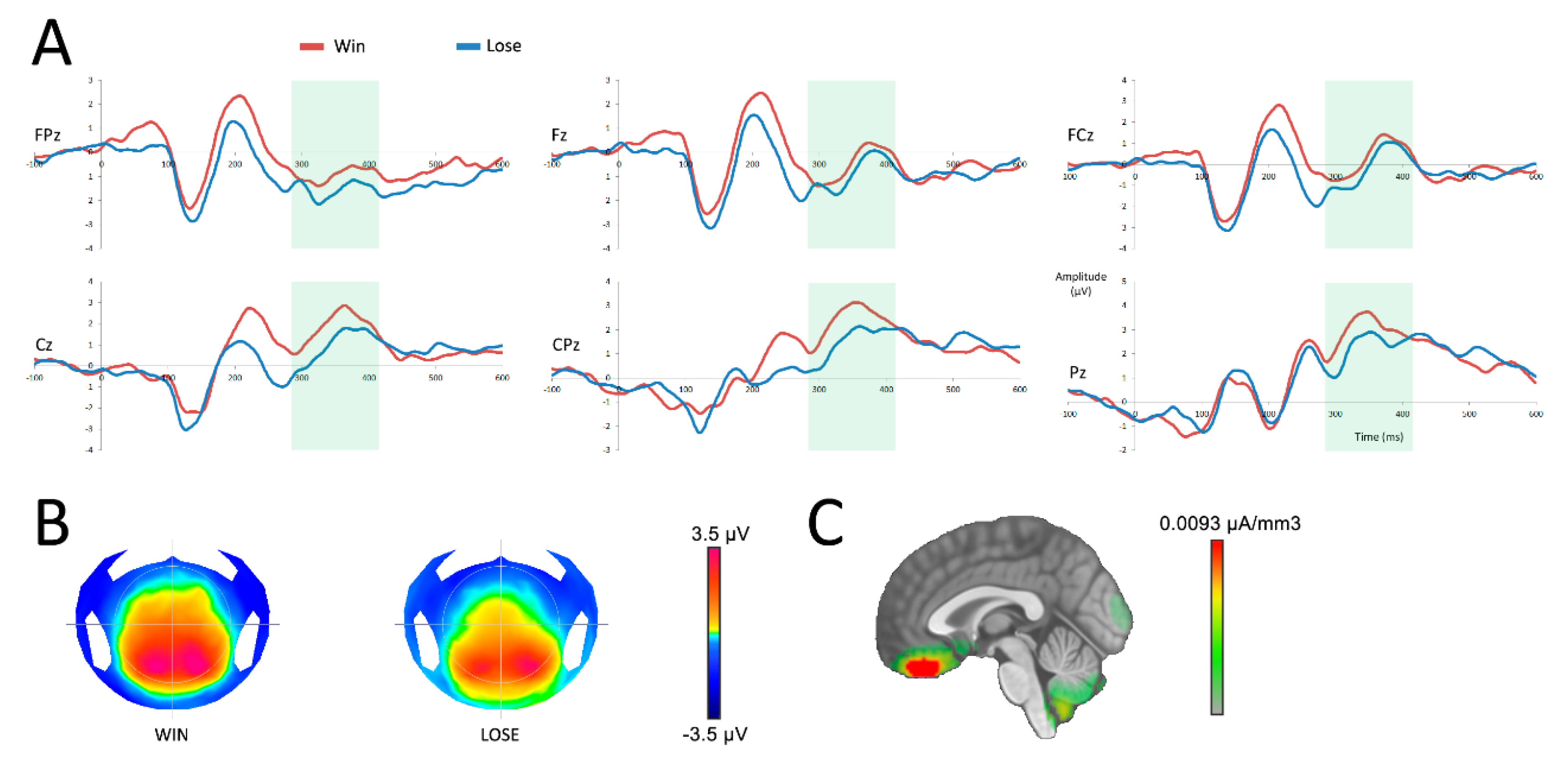
3.2.1. Reward Anticipation
3.2.2. Outcome Processing
3.3. Relationship between Behavioral and Electrophysiological Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Murphy, P.K.; Alexander, P.A. A Motivated Exploration of Motivation Terminology. Contemp. Educ. Psychol. 2000, 25, 3–53. [Google Scholar] [CrossRef]
- Braver, T.S.; Krug, M.K.; Chiew, K.S.; Kool, W.; Westbrook, J.A.; Clement, N.J.; Adcock, R.A.; Barch, D.M.; Botvinick, M.M.; Carver, C.S.; et al. Mechanisms of motivation-cognition interaction: Challenges and opportunities. Cogn. Affect. Behav. Neurosci. 2014, 14, 443–472. [Google Scholar] [CrossRef]
- Schunk, D.H. Coming to Terms with Motivation Constructs. Contemp. Educ. Psychol. 2000, 25, 116–119. [Google Scholar] [CrossRef]
- Mellers, B.A.; McGraw, A.P. Anticipated Emotions as Guides to Choice. Curr. Dir. Psychol. Sci. 2001, 10, 210–214. [Google Scholar] [CrossRef]
- Chong, T.T.-J.; Apps, M.; Giehl, K.; Sillence, A.; Grima, L.L.; Husain, M. Neurocomputational mechanisms underlying subjective valuation of effort costs. PLoS Biol. 2017, 15, e1002598. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, L.; Lebreton, M.; Cléry-Melin, M.-L.; Daunizeau, J.; Pessiglione, M. Neural mechanisms underlying motivation of mental versus physical effort. PLoS Biol. 2012, 10, e1001266. [Google Scholar] [CrossRef] [PubMed]
- Croxson, P.L.; Walton, M.E.; O’Reilly, J.X.; Behrens, T.E.J.; Rushworth, M.F.S. Effort-based cost-benefit valuation and the human brain. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 4531–4541. [Google Scholar] [CrossRef] [PubMed]
- Engström, M.; Karlsson, T.; Landtblom, A.-M.; Craig, A.D.B. Evidence of Conjoint Activation of the Anterior Insular and Cingulate Cortices during Effortful Tasks. Front. Hum. Neurosci. 2014, 8, 1071. [Google Scholar] [CrossRef]
- Kuhnen, C.M.; Knutson, B. The Neural Basis of Financial Risk Taking; Social Science Research Network: Rochester, NY, USA, 2005. [Google Scholar]
- Pessiglione, M.; Seymour, B.; Flandin, G.; Dolan, R.J.; Frith, C.D. Dopamine-dependent prediction errors underpin reward-seeking behaviour in humans. Nature 2006, 442, 1042–1045. [Google Scholar] [CrossRef]
- Prévost, C.; Pessiglione, M.; Météreau, E.; Cléry-Melin, M.-L.; Dreher, J.-C. Separate valuation subsystems for delay and effort decision costs. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 14080–14090. [Google Scholar] [CrossRef]
- Jensen, J.; Smith, A.J.; Willeit, M.; Crawley, A.P.; Mikulis, D.J.; Vitcu, I.; Kapur, S. Separate brain regions code for salience vs. valence during reward prediction in humans. Hum. Brain Mapp. 2007, 28, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Shenhav, A.; Straccia, M.A.; Botvinick, M.M.; Cohen, J.D. Dorsal anterior cingulate and ventromedial prefrontal cortex have inverse roles in both foraging and economic choice. Cogn. Affect. Behav. Neurosci. 2016, 16, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Blair, K.; Marsh, A.A.; Morton, J.; Vythilingam, M.; Jones, M.; Mondillo, K.; Pine, D.C.; Drevets, W.C.; Blair, J.R. Choosing the lesser of two evils, the better of two goods: Specifying the roles of ventromedial prefrontal cortex and dorsal anterior cingulate in object choice. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 11379–11386. [Google Scholar] [CrossRef]
- Treadway, M.T.; Buckholtz, J.W.; Cowan, R.L.; Woodward, N.D.; Li, R.; Ansari, M.S.; Baldwin, R.M.; Schwartzman, A.N.; Kessler, R.M.; Zald, D.H. Dopaminergic mechanisms of individual differences in human effort-based decision-making. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 6170–6176. [Google Scholar] [CrossRef]
- Yang, X.; Huang, J.; Lan, Y.; Zhu, C.; Liu, X.; Wang, Y.; Cheung, E.F.C.; Xie, G.; Chan, R.C.K. Diminished caudate and superior temporal gyrus responses to effort-based decision making in patients with first-episode major depressive disorder. Prog. Neuropsychopharmacol. Biol. Psychiatry 2016, 64, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Yang, X.-H.; Lan, Y.; Zhu, C.-Y.; Liu, X.-Q.; Wang, Y.-F.; Cheung, E.F.C.; Xie, G.-R.; Chan, R.C.K. Neural substrates of the impaired effort expenditure decision making in schizophrenia. Neuropsychology 2016, 30, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.M.; Yates, M.J.; Morton, E.E.; Smillie, L.D. Asymmetric frontal cortical activity predicts effort expenditure for reward. Soc. Cogn. Affect. Neurosci. 2015, 10, 1015–1019. [Google Scholar] [CrossRef]
- Spielberg, J.M.; Miller, G.A.; Warren, S.L.; Engels, A.S.; Crocker, L.D.; Banich, M.T.; Sutton, B.P.; Heller, W. A brain network instantiating approach and avoidance motivation. Psychophysiology 2012, 49, 1200–1214. [Google Scholar] [CrossRef]
- Spielberg, J.M.; Stewart, J.L.; Levin, R.L.; Miller, G.A.; Heller, W. Prefrontal Cortex, Emotion, and Approach/Withdrawal Motivation. Soc. Personal. Psychol. Compass 2008, 2, 135–153. [Google Scholar] [CrossRef]
- Berridge, K.C.; Robinson, T.E.; Aldridge, J.W. Dissecting components of reward: ‘liking’, ‘wanting’, and learning. Curr. Opin. Pharmacol. 2009, 9, 65–73. [Google Scholar] [CrossRef]
- Berridge, K.C.; Kringelbach, M.L. Pleasure Systems in the Brain. Neuron 2015, 86, 646–664. [Google Scholar] [CrossRef] [PubMed]
- Brunia, C.H.; Damen, E.J. Distribution of slow brain potentials related to motor preparation and stimulus anticipation in a time estimation task. Electroencephalogr. Clin. Neurophysiol. 1988, 69, 234–243. [Google Scholar] [CrossRef]
- Böcker, K.B.E.; Baas, J.M.P.; Kenemans, J.L.; Verbaten, M.N. Stimulus-preceding negativity induced by fear: A manifestation of affective anticipation. Int. J. Psychophysiol. 2001, 43, 77–90. [Google Scholar] [CrossRef]
- Fuentemilla, L.; Cucurell, D.; Marco-Pallarés, J.; Guitart-Masip, M.; Morís, J.; Rodríguez-Fornells, A. Electrophysiological correlates of anticipating improbable but desired events. NeuroImage 2013, 78, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.D.B. How do you feel--now? The anterior insula and human awareness. Nat. Rev. Neurosci. 2009, 10, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Böcker, K.B.; Brunia, C.H.; van den Berg-Lenssen, M.M. A spatiotemporal dipole model of the stimulus preceding negativity (SPN) prior to feedback stimuli. Brain Topogr. 1994, 7, 71–88. [Google Scholar] [CrossRef]
- Brunia, C.H.M.; Hackley, S.A.; van Boxtel, G.J.M.; Kotani, Y.; Ohgami, Y. Waiting to perceive: Reward or punishment? Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2011, 122, 858–868. [Google Scholar] [CrossRef]
- Brunia, C.H.; de Jong, B.M.; van den Berg-Lenssen, M.M.; Paans, A.M. Visual feedback about time estimation is related to a right hemisphere activation measured by PET. Exp. Brain Res. 2000, 130, 328–337. [Google Scholar] [CrossRef]
- Sutton, S.; Braren, M.; Zubin, J.; John, E.R. Evoked-potential correlates of stimulus uncertainty. Science 1965, 150, 1187–1188. [Google Scholar] [CrossRef]
- Nieuwenhuis, S.; Aston-Jones, G.; Cohen, J.D. Decision making, the P3, and the locus coeruleus-norepinephrine system. Psychol. Bull. 2005, 131, 510–532. [Google Scholar] [CrossRef]
- Vuillier, L.; Whitebread, D.; Szucs, D. ERP evidence of cognitive strategy change in motivational conditions with varying level of difficulty. Neuropsychologia 2015, 70, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Hughes, G.; Mathan, S.; Yeung, N. EEG indices of reward motivation and target detectability in a rapid visual detection task. NeuroImage 2013, 64, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, R.Z.; Cottone, L.A.; Jia, Z.; Maloney, T.; Volkow, N.D.; Squires, N.K. The effect of graded monetary reward on cognitive event-related potentials and behavior in young healthy adults. Int. J. Psychophysiol. Off. J. Int. Organ. Psychophysiol. 2006, 62, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Kleih, S.C.; Nijboer, F.; Halder, S.; Kübler, A. Motivation modulates the P300 amplitude during brain-computer interface use. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2010, 121, 1023–1031. [Google Scholar] [CrossRef]
- Yeung, N.; Sanfey, A.G. Independent coding of reward magnitude and valence in the human brain. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 6258–6264. [Google Scholar] [CrossRef]
- Pessiglione, M.; Schmidt, L.; Draganski, B.; Kalisch, R.; Lau, H.; Dolan, R.J.; Frith, C.D. How the brain translates money into force: A neuroimaging study of subliminal motivation. Science 2007, 316, 904–906. [Google Scholar] [CrossRef]
- Docx, L.; de la Asuncion, J.; Sabbe, B.; Hoste, L.; Baeten, R.; Warnaerts, N.; Morrens, M. Effort discounting and its association with negative symptoms in schizophrenia. Cognit. Neuropsychiatry 2015, 20, 172–185. [Google Scholar] [CrossRef]
- Treadway, M.T.; Buckholtz, J.W.; Schwartzman, A.N.; Lambert, W.E.; Zald, D.H. Worth the ‘EEfRT’? The Effort Expenditure for Rewards Task as an Objective Measure of Motivation and Anhedonia. PLoS ONE 2009, 4, e6598. [Google Scholar] [CrossRef]
- Ubl, B.; Kuehner, C.; Kirsch, P.; Ruttorf, M.; Flor, H.; Diener, C. Neural reward processing in individuals remitted from major depression. Psychol. Med. 2015, 45, 3549–3558. [Google Scholar] [CrossRef]
- Chumbley, J.; Fehr, E. Does general motivation energize financial reward-seeking behavior? Evidence from an effort task. PLoS ONE 2014, 9, e101936. [Google Scholar] [CrossRef]
- Wolf, D.H.; Satterthwaite, T.D.; Kantrowitz, J.J.; Katchmar, N.; Vandekar, L.; Elliott, M.A.; Ruparel, K. Amotivation in schizophrenia: Integrated assessment with behavioral, clinical, and imaging measures. Schizophr. Bull. 2014, 40, 1328–1337. [Google Scholar] [CrossRef]
- St Onge, J.R.; Floresco, S.B. Dopaminergic modulation of risk-based decision making. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2009, 34, 681–697. [Google Scholar] [CrossRef]
- Floresco, S.B.; Tse, M.T.L.; Ghods-Sharifi, S. Dopaminergic and glutamatergic regulation of effort- and delay-based decision making. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2008, 33, 1966–1979. [Google Scholar] [CrossRef]
- Treadway, M.T.; Zald, D.H. Parsing Anhedonia: Translational Models of Reward-Processing Deficits in Psychopathology. Curr. Dir. Psychol. Sci. 2013, 22, 244–249. [Google Scholar] [CrossRef]
- Whitton, A.E.; Treadway, M.T.; Pizzagalli, D.A. Reward processing dysfunction in major depression, bipolar disorder and schizophrenia. Curr. Opin. Psychiatry 2015, 28, 7–12. [Google Scholar] [CrossRef]
- Treadway, M.T.; Peterman, J.S.; Zald, D.H.; Park, S. Impaired effort allocation in patients with schizophrenia. Schizophr. Res. 2015, 161, 382–385. [Google Scholar] [CrossRef]
- Mata, F.; Treadway, M.; Kwok, A.; Truby, H.; Yücel, M.; Stout, J.C.; Verdejo-Garcia, A. Reduced Willingness to Expend Effort for Reward in Obesity: Link to Adherence to a 3-Month Weight Loss Intervention. Obes. Silver Spring Md 2017, 25, 1676–1681. [Google Scholar] [CrossRef]
- Lawn, W.; Freeman, T.P.; Pope, R.A.; Joye, A.; Harvey, L.; Hindocha, C.; Mokrysz, C.; Moss, A.; Wall, M.B.; Bloomfield, M.A.; et al. Acute and chronic effects of cannabinoids on effort-related decision-making and reward learning: An evaluation of the cannabis “amotivational” hypotheses. Psychopharmacology (Berl.) 2016, 233, 3537–3552. [Google Scholar] [CrossRef]
- Gehring, W.J.; Willoughby, A.R. The medial frontal cortex and the rapid processing of monetary gains and losses. Science 2002, 295, 2279–2282. [Google Scholar] [CrossRef]
- Holroyd, C.B.; Hajcak, G.; Larsen, J.T. The good, the bad and the neutral: Electrophysiological responses to feedback stimuli. Brain Res. 2006, 1105, 93–101. [Google Scholar] [CrossRef]
- Plaisant, O.; Courtois, R.; Réveillère, C.; Mendelsohn, G.A.; John, O.P. Validation par analyse factorielle du Big Five Inventory français (BFI-Fr). Analyse convergente avec le NEO-PI-R. Ann. Méd.-Psychol. Rev. Psychiatr. 2010, 168, 97–106. [Google Scholar] [CrossRef]
- Fervaha, G.; Graff-Guerrero, A.; Zakzanis, K.K.; Foussias, G.; Agid, O.; Remington, G. Incentive motivation deficits in schizophrenia reflect effort computation impairments during cost-benefit decision-making. J. Psychiatr. Res. 2013, 47, 1590–1596. [Google Scholar] [CrossRef] [PubMed]
- Downloads - Cartool Community. Available online: https://sites.google.com/site/cartoolcommunity/files (accessed on 7 May 2020).
- Walentowska, W.; Paul, K.; Severo, M.C.; Moors, A.; Pourtois, G. Relevance and uncertainty jointly influence reward anticipation at the level of the SPN ERP component. Int. J. Psychophysiol. Off. J. Int. Organ. Psychophysiol. 2018, 132, 287–297. [Google Scholar] [CrossRef]
- Perrin, F.; Bertrand, O.; Pernier, J. Scalp current density mapping: Value and estimation from potential data. IEEE Trans. Biomed. Eng. 1987, 34, 283–288. [Google Scholar] [CrossRef]
- Cui, J.; Chen, Y.; Wang, Y.; Shum, D.H.K.; Chan, R.C.K. Neural correlates of uncertain decision making: ERP evidence from the Iowa Gambling Task. Front. Hum. Neurosci. 2013, 7, 776. [Google Scholar] [CrossRef]
- Bianchin, M.; Angrilli, A. Decision Preceding Negativity in the Iowa Gambling Task: An ERP study. Brain Cogn. 2011, 75, 273–280. [Google Scholar] [CrossRef]
- Hajcak, G.; Moser, J.S.; Holroyd, C.B.; Simons, R.F. The feedback-related negativity reflects the binary evaluation of good versus bad outcomes. Biol. Psychol. 2006, 71, 148–154. [Google Scholar] [CrossRef]
- Hajcak, G.; Holroyd, C.B.; Moser, J.S.; Simons, R.F. Brain potentials associated with expected and unexpected good and bad outcomes. Psychophysiology 2005, 42, 161–170. [Google Scholar] [CrossRef]
- Bland, A.R.; Schaefer, A. Electrophysiological correlates of decision making under varying levels of uncertainty. Brain Res. 2011, 1417, 55–66. [Google Scholar] [CrossRef]
- Chapman, L.J.; Chapman, J.P.; Raulin, M.L. Scales for physical and social anhedonia. J. Abnorm. Psychol. 1976, 85, 374–382. [Google Scholar] [CrossRef]
- Loas, G.; Dubal, S.; Perot, P.; Tirel, F.; Nowaczkowski, P.; Pierson, A. [Validation of the French version of the Snaith-Hamilton Pleasure Scale (SHAPS, Snaith et al. 1995). Determination of the statistical parameters in 208 normal subjects and 103 hospitalized patients presenting with depression or schizophrenia]. L’Encéphale 1997, 23, 454–458. [Google Scholar]
- Wardle, M.C.; Treadway, M.T.; Mayo, L.M.; Zald, D.H.; de Wit, H. Amping up effort: Effects of d-amphetamine on human effort-based decision-making. J. Neurosci. Off. J. Soc. Neurosci. 2011, 31, 16597–16602. [Google Scholar] [CrossRef]
- Kotani, Y.; Hiraku, S.; Suda, K.; Aihara, Y. Effect of positive and negative emotion on stimulus-preceding negativity prior to feedback stimuli. Psychophysiology 2001, 38, 873–878. [Google Scholar] [CrossRef]
- Hillman, C.H.; Apparies, R.J.; Hatfield, B.D. Motor and nonmotor event-related potentials during a complex processing task. Psychophysiology 2000, 37, 731–736. [Google Scholar] [CrossRef]
- Arulpragasam, A.R.; Cooper, J.A.; Nuutinen, M.R.; Treadway, M.T. Corticoinsular circuits encode subjective value expectation and violation for effortful goal-directed behavior. Proc. Natl. Acad. Sci. USA 2018, 115, E5233–E5242. [Google Scholar] [CrossRef]
- Wager, T.D.; Barrett, L.F. From affect to control: Functional specialization of the insula in motivation and regulation. BioRxiv 2017, 102368. [Google Scholar] [CrossRef]
- Kennis, M.; Rademaker, A.R.; Geuze, E. Neural correlates of personality: An integrative review. Neurosci. Biobehav. Rev. 2013, 37, 73–95. [Google Scholar] [CrossRef]
- Deng, Y.; Li, S.; Zhou, R.; Walter, M. Motivation but not valence modulates neuroticism-dependent cingulate cortex and insula activity. Hum. Brain Mapp. 2018, 39, 1664–1672. [Google Scholar] [CrossRef]
- Ferdinand, N.K.; Kray, J. Age-related changes in processing positive and negative feedback: Is there a positivity effect for older adults? Biol. Psychol. 2013, 94, 235–241. [Google Scholar] [CrossRef]
- Rigoni, D.; Polezzi, D.; Rumiati, R.; Guarino, R.; Sartori, G. When people matter more than money: An ERPs study. Brain Res. Bull. 2010, 81, 445–452. [Google Scholar] [CrossRef]
- Giustiniani, J.; Gabriel, D.; Nicolier, M.; Monnin, J.; Haffen, E. Neural Correlates of Successful and Unsuccessful Strategical Mechanisms Involved in Uncertain Decision-Making. PLoS ONE 2015, 10, e0130871. [Google Scholar] [CrossRef]
- Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2007, 118, 2128–2148. [Google Scholar] [CrossRef]
- Polich, J. Clinical application of the P300 event-related brain potential. Phys. Med. Rehabil. Clin. N. Am. 2004, 15, 133–161. [Google Scholar] [CrossRef]
- Chase, H.W.; Swainson, R.; Durham, L.; Benham, L.; Cools, R. Feedback-related Negativity Codes Prediction Error but Not Behavioral Adjustment during Probabilistic Reversal Learning. J. Cogn. Neurosci. 2010, 23, 936–946. [Google Scholar] [CrossRef]
- Locke, H.S.; Braver, T.S. Motivational influences on cognitive control: Behavior, brain activation, and individual differences. Cogn. Affect. Behav. Neurosci. 2008, 8, 99–112. [Google Scholar] [CrossRef]
- Polezzi, D.; Sartori, G.; Rumiati, R.; Vidotto, G.; Daum, I. Brain correlates of risky decision-making. NeuroImage 2010, 49, 1886–1894. [Google Scholar] [CrossRef]
- Takayoshi, H.; Onoda, K.; Yamaguchi, S. Do Event-Related Evoked Potentials Reflect Apathy Tendency and Motivation? Front. Hum. Neurosci. 2018, 12. [Google Scholar] [CrossRef]
- Dubal, S.; Pierson, A.; Jouvent, R. Focused attention in anhedonia: A P3 study. Psychophysiology 2000, 37, 711–714. [Google Scholar] [CrossRef]
- Horovitz, S.G.; Skudlarski, P.; Gore, J.C. Correlations and dissociations between BOLD signal and P300 amplitude in an auditory oddball task: A parametric approach to combining fMRI and ERP. Magn. Reson. Imaging 2002, 20, 319–325. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, Y.; Sui, D.; Tang, Y.-Y. Neural correlates of cultural differences in moral decision making: A combined ERP and sLORETA study. Neuroreport 2014, 25, 110–116. [Google Scholar] [CrossRef]
- Yacubian, J.; Gläscher, J.; Schroeder, K.; Sommer, T.; Braus, D.F.; Büchel, C. Dissociable systems for gain- and loss-related value predictions and errors of prediction in the human brain. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 9530–9537. [Google Scholar] [CrossRef]
- Rogers, R.D.; Ramnani, N.; Mackay, C.; Wilson, J.L.; Jezzard, P.; Carter, C.S.; Smith, S.M. Distinct portions of anterior cingulate cortex and medial prefrontal cortex are activated by reward processing in separable phases of decision-making cognition. Biol. Psychiatry 2004, 55, 594–602. [Google Scholar] [CrossRef]
- Raymond, J. Interactions of attention, emotion and motivation. Prog. Brain Res. 2009, 176, 293–308. [Google Scholar] [CrossRef]
- Umemoto, A.; Holroyd, C.B. Neural mechanisms of reward processing associated with depression-related personality traits. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2017, 128, 1184–1196. [Google Scholar] [CrossRef]
- Salamone, J.D.; Correa, M.; Farrar, A.; Mingote, S.M. Effort-related functions of nucleus accumbens dopamine and associated forebrain circuits. Psychopharmacology (Berl.) 2007, 191, 461–482. [Google Scholar] [CrossRef]
- Wade, T.R.; de Wit, H.; Richards, J.B. Effects of dopaminergic drugs on delayed reward as a measure of impulsive behavior in rats. Psychopharmacology (Berl.) 2000, 150, 90–101. [Google Scholar]
- Massar, S.A.A.; Libedinsky, C.; Weiyan, C.; Huettel, S.A.; Chee, M.W.L. Separate and overlapping brain areas encode subjective value during delay and effort discounting. NeuroImage 2015, 120, 104–113. [Google Scholar] [CrossRef]
- Winstanley, C.A.; Theobald, D.E.H.; Dalley, J.W.; Cardinal, R.N.; Robbins, T.W. Double dissociation between serotonergic and dopaminergic modulation of medial prefrontal and orbitofrontal cortex during a test of impulsive choice. Cereb. Cortex N.Y. N 1991 2006, 16, 106–114. [Google Scholar] [CrossRef]
- Schuermann, B.; Endrass, T.; Kathmann, N. Neural correlates of feedback processing in decision-making under risk. Front. Hum. Neurosci. 2012, 6, 204. [Google Scholar] [CrossRef]
- Gheza, D.; De Raedt, R.; Baeken, C.; Pourtois, G. Integration of reward with cost anticipation during performance monitoring revealed by ERPs and EEG spectral perturbations. NeuroImage 2018, 173, 153–164. [Google Scholar] [CrossRef]
- Protopopescu, X.; Pan, H.; Altemus, M.; Tuescher, O.; Polanecsky, M.; McEwen, B.; Silbersweig, D.; Stern, E. Orbitofrontal cortex activity related to emotional processing changes across the menstrual cycle. Proc. Natl. Acad. Sci. USA 2005, 102, 16060–16065. [Google Scholar] [CrossRef]
- Tanner, D.; Morgan-Short, K.; Luck, S.J. How inappropriate high-pass filters can produce artifactual effects and incorrect conclusions in ERP studies of language and cognition. Psychophysiology 2015, 52, 997–1009. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Easy Task | Hard Task | |||||
|---|---|---|---|---|---|---|
| 10% | 50% | 90% | 10% | 50% | 90% | |
| Win | 3.8 ± 0.5 | 10.6 ± 3.4 | 10.7 ± 3.0 | 0.2 ± 0.5 | 9.5 ± 3.4 | 25.4 ± 3.0 |
| Loss | 32.6 ± 4.3 | 10.1 ± 3.8 | 1.3 ± 0.6 | 3.4 ± 4.3 | 10.0 ± 3.8 | 2.8 ± 0.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giustiniani, J.; Nicolier, M.; Teti Mayer, J.; Chabin, T.; Masse, C.; Galmès, N.; Pazart, L.; Trojak, B.; Bennabi, D.; Vandel, P.; et al. Event-Related Potentials (ERP) Indices of Motivation during the Effort Expenditure for Reward Task. Brain Sci. 2020, 10, 283. https://doi.org/10.3390/brainsci10050283
Giustiniani J, Nicolier M, Teti Mayer J, Chabin T, Masse C, Galmès N, Pazart L, Trojak B, Bennabi D, Vandel P, et al. Event-Related Potentials (ERP) Indices of Motivation during the Effort Expenditure for Reward Task. Brain Sciences. 2020; 10(5):283. https://doi.org/10.3390/brainsci10050283
Chicago/Turabian StyleGiustiniani, Julie, Magali Nicolier, Juliana Teti Mayer, Thibault Chabin, Caroline Masse, Nathan Galmès, Lionel Pazart, Benoit Trojak, Djamila Bennabi, Pierre Vandel, and et al. 2020. "Event-Related Potentials (ERP) Indices of Motivation during the Effort Expenditure for Reward Task" Brain Sciences 10, no. 5: 283. https://doi.org/10.3390/brainsci10050283
APA StyleGiustiniani, J., Nicolier, M., Teti Mayer, J., Chabin, T., Masse, C., Galmès, N., Pazart, L., Trojak, B., Bennabi, D., Vandel, P., Haffen, E., & Gabriel, D. (2020). Event-Related Potentials (ERP) Indices of Motivation during the Effort Expenditure for Reward Task. Brain Sciences, 10(5), 283. https://doi.org/10.3390/brainsci10050283