β-Caryophyllene: A Sesquiterpene with Countless Biological Properties




 ,
, 
 and
and 
Abstract
1. Introduction
- The activation of Gi/o α-subunit inhibits the activity of adenylate cyclase, the enzyme which converts ATP in cAMP (cyclic AMP) and activates phospholipase C (PLC), determining calcium release from IP3 (inositol trisphosphate)-sensitive calcium channels, increasing the intracellular level of calcium;
- The activation of the complex formed by Gi/o β- and γ-subunits, which modulate various pathways modifying the expression and the activity of many target proteins [15].
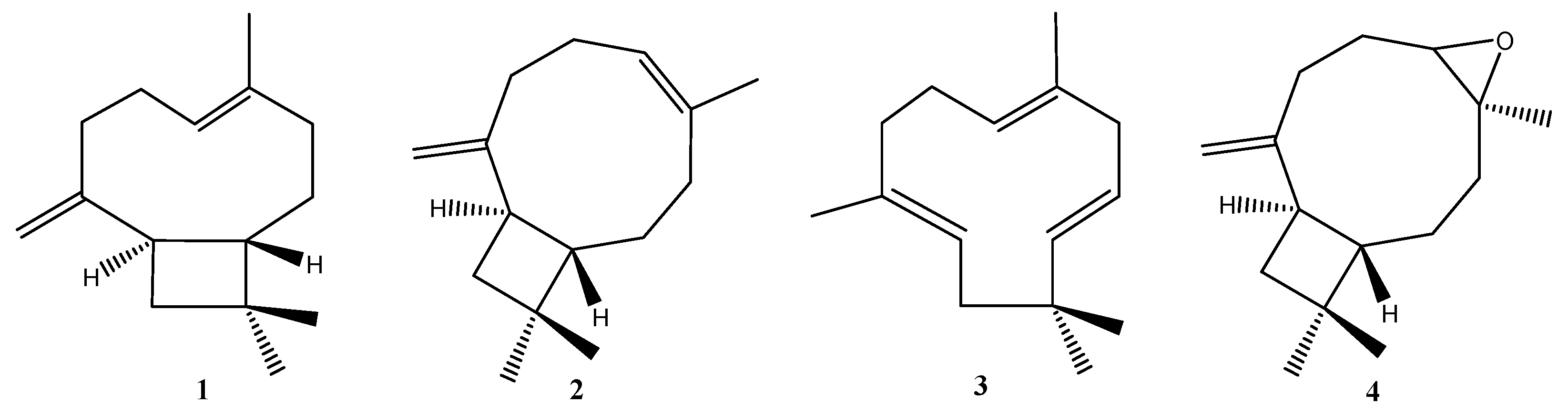
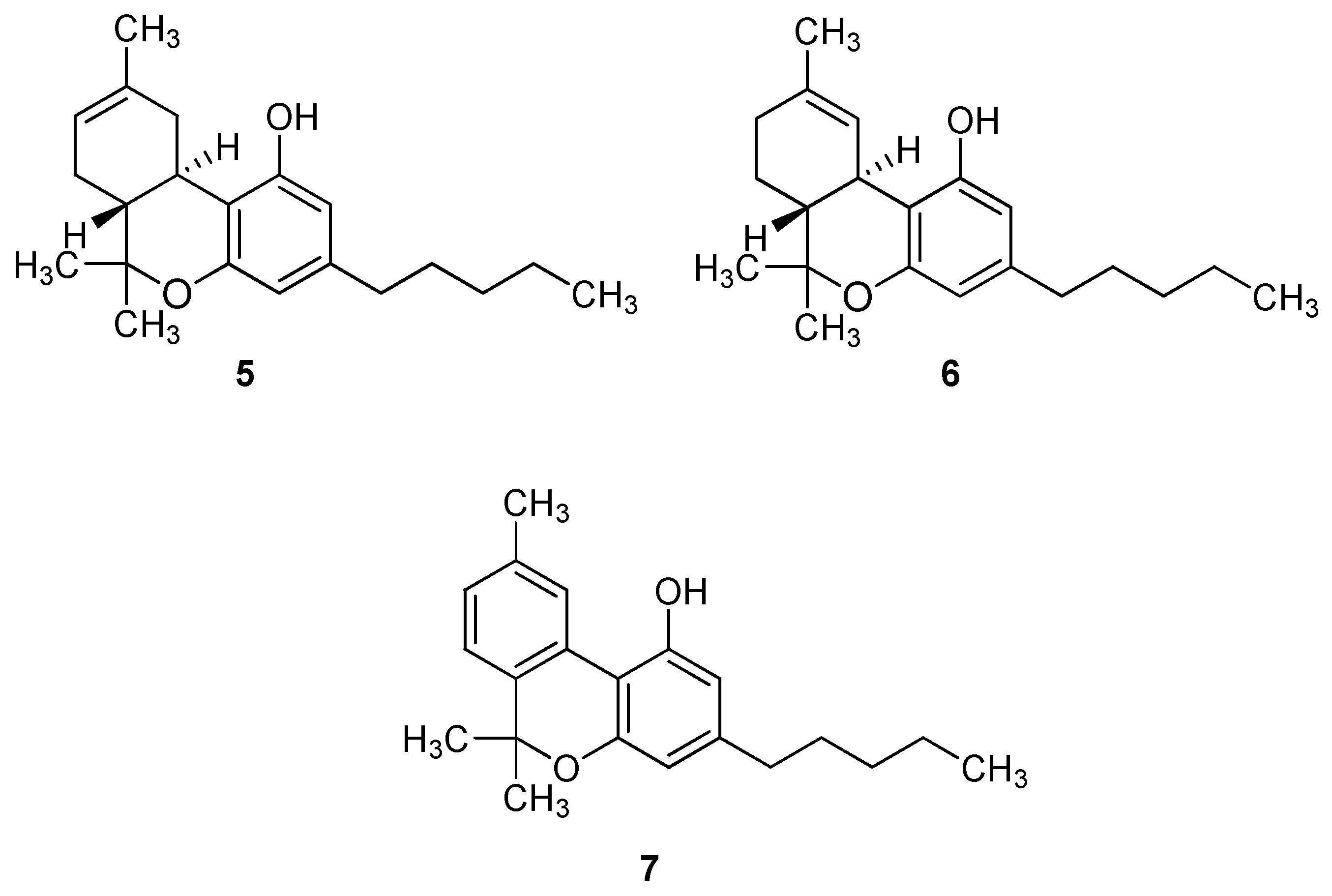
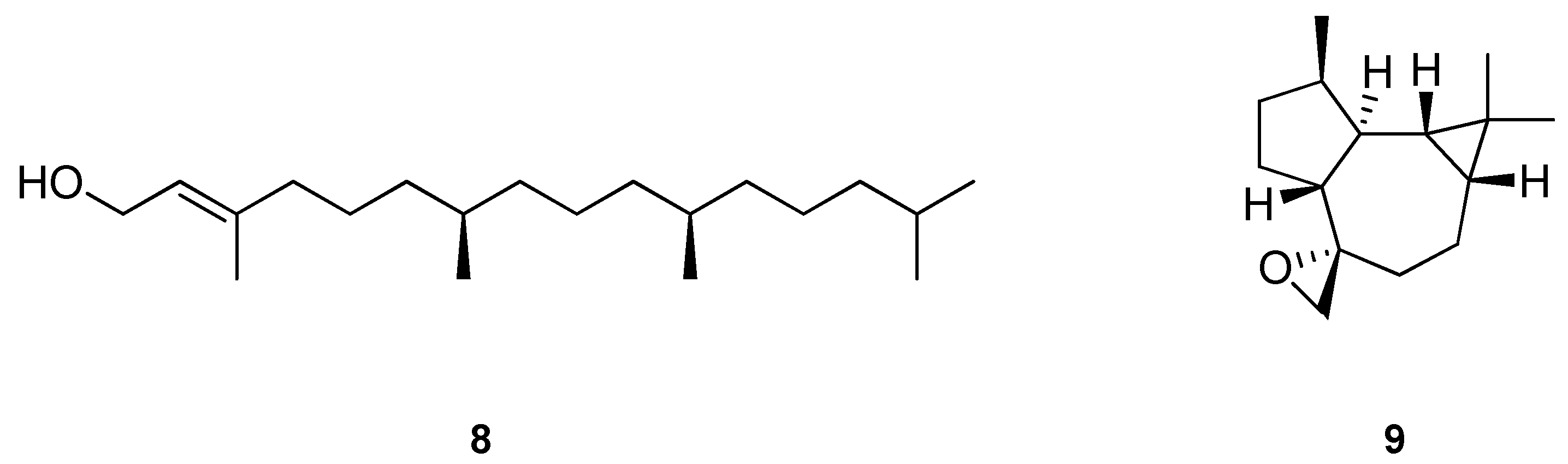
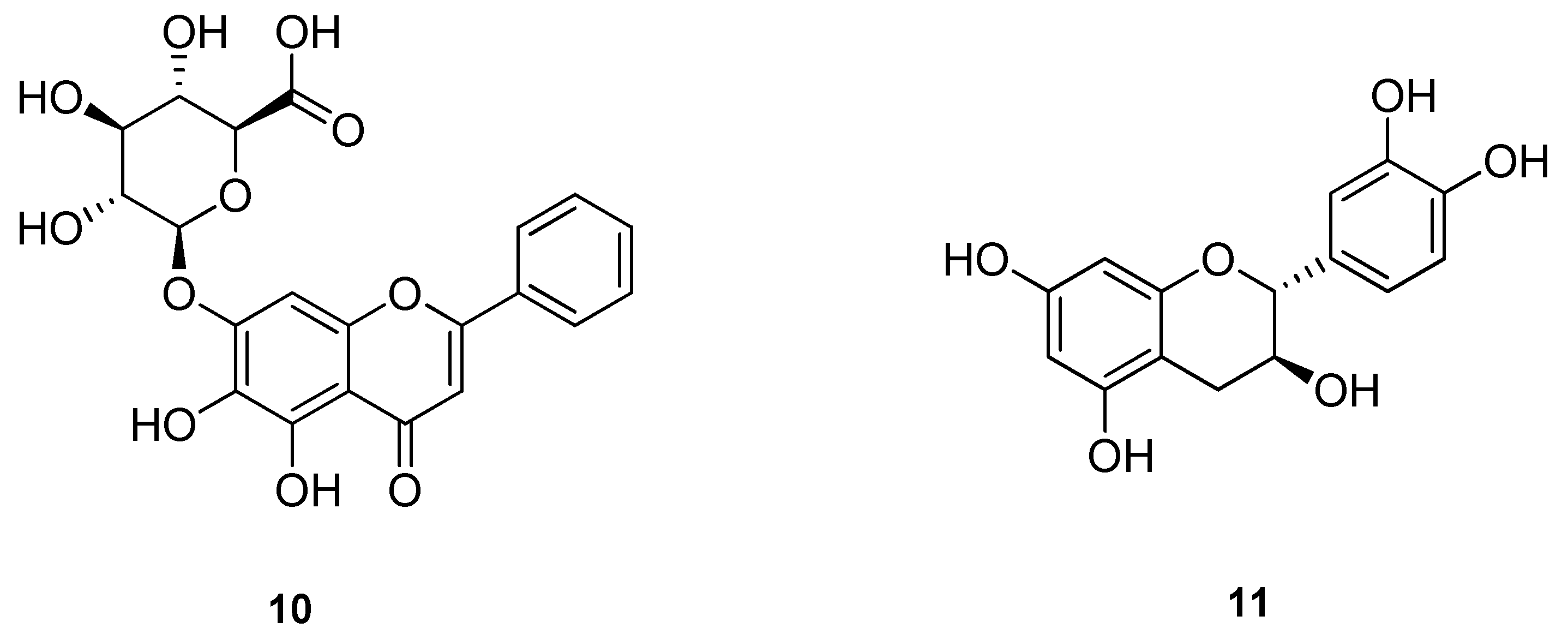
2. Biological properties of β-caryophyllene
2.1. β-Caryophyllene and Nervous System
2.2. β-Caryophyllene and Cancer
2.3. β-Caryophyllene and Inflammatory Diseases
- Reduce carboxylic groups of protein and ROS in the liver, normalizing the levels;
- Increase glutathione levels and reduce lipoperoxidation [56].
2.4. β-Caryophyllene and Micro-Organisms
2.5. β-Caryophyllene and Osteoporosis
2.6. β-Caryophyllene Toxicity
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Russo, A.; Perri, M.; Cione, E.; Di Gioia, M.L.; Nardi, M.; Cristina Caroleo, M. Biochemical and chemical characterization of Cynara cardunculus L. extract and its potential use as co-adjuvant therapy of chronic myeloid leukemia. J. Ethnopharmacol. 2017, 202, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Cappello, A.R.; Dolce, V.; Iacopetta, D.; Martello, M.; Fiorillo, M.; Curcio, R.; Muto, L.; Dhanyalayam, D. Bergamot (Citrus bergamia Risso) Flavonoids and Their Potential Benefits in Human Hyperlipidemia and Atherosclerosis: An Overview. Mini Rev. Med. Chem. 2016, 16, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Tundis, R.; Iacopetta, D.; Sinicropi, M.S.; Bonesi, M.; Leporini, M.; Passalacqua, N.G.; Ceramella, J.; Menichini, F.; Loizzo, M.R. Assessment of antioxidant, antitumor and pro-apoptotic effects of Salvia fruticosa Mill. subsp. thomasii (Lacaita) Brullo, Guglielmo, Pavone & Terrasi (Lamiaceae). Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2017, 106, 155–164. [Google Scholar] [CrossRef]
- Ceramella, J.; Loizzo, M.R.; Iacopetta, D.; Bonesi, M.; Sicari, V.; Pellicano, T.M.; Saturnino, C.; Malzert-Freon, A.; Tundis, R.; Sinicropi, M.S. Anchusa azurea Mill. (Boraginaceae) aerial parts methanol extract interfering with cytoskeleton organization induces programmed cancer cells death. Food Funct. 2019, 10, 4280–4290. [Google Scholar] [CrossRef]
- Chimento, A.; Sirianni, R.; Saturnino, C.; Caruso, A.; Sinicropi, M.S.; Pezzi, V. Resveratrol and Its Analogs As Antitumoral Agents For Breast Cancer Treatment. Mini Rev. Med. Chem. 2016, 16, 699–709. [Google Scholar] [CrossRef]
- Ruffo, M.; Parisi, O.I.; Scrivano, L.; Restuccia, D.; Amone, F.; Sinicropi, M.S.; Malivindi, R.; Aiello, G.; Puoci, F. Role of Calabrian Black Rice in Metabolic Syndrome: In vitro Evaluation of Oryza sativa L. Indica Biological Properties. Curr. Nutr. Food Sci. 2018, 14, 121–127. [Google Scholar] [CrossRef]
- Chimento, A.; De Amicis, F.; Sirianni, R.; Sinicropi, M.S.; Puoci, F.; Casaburi, I.; Saturnino, C.; Pezzi, V. Progress to Improve Oral Bioavailability and Beneficial Effects of Resveratrol. Int. J. Mol. Sci. 2019, 20, 1381. [Google Scholar] [CrossRef]
- Plastina, P.; Apriantini, A.; Meijerink, J.; Witkamp, R.; Gabriele, B.; Fazio, A. In Vitro Anti-Inflammatory and Radical Scavenging Properties of Chinotto (Citrus myrtifolia Raf.) Essential Oils. Nutrients 2018, 10, 783. [Google Scholar] [CrossRef]
- Machado, K.D.C.; Islam, M.T.; Ali, E.S.; Rouf, R.; Uddin, S.J.; Dev, S.; Shilpi, J.A.; Shill, M.C.; Reza, H.M.; Das, A.K.; et al. A systematic review on the neuroprotective perspectives of beta-caryophyllene. Phytother. Res. 2018, 32, 2376–2388. [Google Scholar] [CrossRef]
- Oliveira, G.; Machado, K.C.; Machado, K.C.; da Silva, A.; Feitosa, C.M.; de Castro Almeida, F.R. Non-clinical toxicity of beta-caryophyllene, a dietary cannabinoid: Absence of adverse effects in female Swiss mice. Regul. Toxicol. Pharmacol. 2018, 92, 338–346. [Google Scholar] [CrossRef]
- Ríos, M.Y.; Castrejón, F.; Robledo, N.; León, I.; Rojas, G.; Navarro, V. Chemical Composition and Antimicrobial Activity of the Essential Oils from Annona cherimola (Annonaceae). Rev. De La Soc. Quím. De Méx. 2003, 47, 139–142. [Google Scholar]
- Gertsch, J.; Leonti, M.; Raduner, S.; Racz, I.; Chen, J.Z.; Xie, X.Q.; Altmann, K.H.; Karsak, M.; Zimmer, A. Beta-caryophyllene is a dietary cannabinoid. Proc. Natl. Acad. Sci. USA 2008, 105, 9099–9104. [Google Scholar] [CrossRef]
- Fidyt, K.; Fiedorowicz, A.; Strzadala, L.; Szumny, A. beta-caryophyllene and beta-caryophyllene oxide-natural compounds of anticancer and analgesic properties. Cancer Med. 2016, 5, 3007–3017. [Google Scholar] [CrossRef]
- Hartsel, J.A.; Eades, J.; Hickory, B.; Makriyannis, A. Cannabis sativa and Hemp. In Nutraceuticals: Efficacy, Safety and Toxicity; Gupta, R.C., Ed.; Academic Press: Cambridge, CA, USA, 2016; Volume 53, pp. 735–754. [Google Scholar]
- Demuth, D.G.; Molleman, A. Cannabinoid signalling. Life Sci. 2006, 78, 549–563. [Google Scholar] [CrossRef]
- Saturnino, C.; Iacopetta, D.; Sinicropi, M.S.; Rosano, C.; Caruso, A.; Caporale, A.; Marra, N.; Marengo, B.; Pronzato, M.A.; Parisi, O.I.; et al. N-Alkyl Carbazole Derivatives as New Tools for Alzheimer’s Disease: Preliminary Studies. Molecules 2014, 19, 9307–9317. [Google Scholar] [CrossRef]
- Catalano, A.; Carocci, A.; Corbo, F.; Franchini, C.; Muraglia, M.; Scilimati, A.; De Bellis, M.; De Luca, A.; Camerino, D.C.; Sinicropi, M.S.; et al. Constrained analogues of tocainide as potent skeletal muscle sodium channel blockers towards the development of antimyotonic agents. Eur. J. Med. Chem. 2008, 43, 2535–2540. [Google Scholar] [CrossRef]
- Iacopetta, D.; Rosano, C.; Puoci, F.; Parisi, O.I.; Saturnino, C.; Caruso, A.; Longo, P.; Ceramella, J.; Malzert-Freon, A.; Dallemagne, P.; et al. Multifaceted properties of 1,4-dimethylcarbazoles: Focus on trimethoxybenzamide and trimethoxyphenylurea derivatives as novel human topoisomerase II inhibitors. Eur. J. Pharm. Sci. 2017, 96, 263–272. [Google Scholar] [CrossRef]
- Ceramella, J.; Caruso, A.; Occhiuzzi, M.A.; Iacopetta, D.; Barbarossa, A.; Rizzuti, B.; Dallemagne, P.; Rault, S.; El-Kashef, H.; Saturnino, C.; et al. Benzothienoquinazolinones as new multi-target scaffolds: Dual inhibition of human Topoisomerase I and tubulin polymerization. Eur. J. Med. Chem. 2019, 181, 111583. [Google Scholar] [CrossRef]
- Caruso, A.; Iacopetta, D.; Puoci, F.; Cappello, A.R.; Saturnino, C.; Sinicropi, M.S. Carbazole derivatives: A promising scenario for breast cancer treatment. Mini Rev. Med. Chem. 2016, 16, 630–643. [Google Scholar] [CrossRef]
- Nettore, I.C.; Rocca, C.; Mancino, G.; Albano, L.; Amelio, D.; Grande, F.; Puoci, F.; Pasqua, T.; Desiderio, S.; Mazza, R.; et al. Quercetin and its derivative Q2 modulate chromatin dynamics in adipogenesis and Q2 prevents obesity and metabolic disorders in rats. J. Nutr. Biochem. 2019, 69, 151–162. [Google Scholar] [CrossRef]
- Ojha, S.; Javed, H.; Azimullah, S.; Haque, M.E. beta-Caryophyllene, a phytocannabinoid attenuates oxidative stress, neuroinflammation, glial activation, and salvages dopaminergic neurons in a rat model of Parkinson disease. Mol. Cell. Biochem. 2016, 418, 59–70. [Google Scholar] [CrossRef]
- Sousa, O.V.; Silverio, M.S.; Del-Vechio-Vieira, G.; Matheus, F.C.; Yamamoto, C.H.; Alves, M.S. Antinociceptive and anti-inflammatory effects of the essential oil from Eremanthus erythropappus leaves. J. Pharm. Pharmacol. 2008, 60, 771–777. [Google Scholar] [CrossRef]
- Chang, H.J.; Kim, J.M.; Lee, J.C.; Kim, W.K.; Chun, H.S. Protective effect of beta-caryophyllene, a natural bicyclic sesquiterpene, against cerebral ischemic injury. J. Med. Food 2013, 16, 471–480. [Google Scholar] [CrossRef]
- Guo, K.; Mou, X.; Huang, J.; Xiong, N.; Li, H. Trans-caryophyllene suppresses hypoxia-induced neuroinflammatory responses by inhibiting NF-kappaB activation in microglia. J. Mol. Neurosci. 2014, 54, 41–48. [Google Scholar] [CrossRef]
- Hu, Y.; Zeng, Z.; Wang, B.; Guo, S. Trans-caryophyllene inhibits amyloid beta (Abeta) oligomer-induced neuroinflammation in BV-2 microglial cells. Int. Immunopharmacol. 2017, 51, 91–98. [Google Scholar] [CrossRef]
- Ito, D.; Imai, Y.; Ohsawa, K.; Nakajima, K.; Fukuuchi, Y.; Kohsaka, S. Microglia-specific localisation of a novel calcium binding protein, Iba1. Brain Res. Mol. 1998, 57, 1–9. [Google Scholar] [CrossRef]
- Bunge, M.B.; Bunge, R.P.; Ris, H. Ultrastructural study of remyelination in an experimental lesion in adult cat spinal cord. J. Biophys. Biochem. Cytol. 1961, 10, 67–94. [Google Scholar] [CrossRef]
- Wang, G.F.; Ma, W.B.; Du, J.W. beta-Caryophyllene (BCP) ameliorates MPP plus induced cytotoxicity. Biomed. Pharmacother. 2018, 103, 1086–1091. [Google Scholar] [CrossRef]
- Viveros-Paredes, J.M.; Gonzalez-Castaneda, R.E.; Gertsch, J.; Chaparro-Huerta, V.; Lopez-Roa, R.I.; Vazquez-Valls, E.; Beas-Zarate, C.; Camins-Espuny, A.; Flores-Soto, M.E. Neuroprotective Effects of beta-Caryophyllene against Dopaminergic Neuron Injury in a Murine Model of Parkinson’s Disease Induced by MPTP. Pharmaceuticals 2017, 10, 60. [Google Scholar] [CrossRef]
- DeNicola, G.M.; Karreth, F.A.; Humpton, T.J.; Gopinathan, A.; Wei, C.; Frese, K.; Mangal, D.; Yu, K.H.; Yeo, C.J.; Calhoun, E.S.; et al. Oncogene-induced Nrf2 transcription promotes ROS detoxification and tumorigenesis. Nature 2011, 475, 106–109. [Google Scholar] [CrossRef]
- Tian, X.; Peng, J.; Zhong, J.; Yang, M.; Pang, J.; Lou, J.; Li, M.; An, R.; Zhang, Q.; Xu, L.; et al. beta-Caryophyllene protects in vitro neurovascular unit against oxygen-glucose deprivation and re-oxygenation-induced injury. J. Neurochem. 2016, 139, 757–768. [Google Scholar] [CrossRef]
- Assis, L.C.; Straliotto, M.R.; Engel, D.; Hort, M.A.; Dutra, R.C.; de Bem, A.F. beta-Caryophyllene protects the C6 glioma cells against glutamate-induced excitotoxicity through the Nrf2 pathway. Neuroscience 2014, 279, 220–231. [Google Scholar] [CrossRef]
- Santos, N.A.G.; Martins, N.M.; Sisti, F.M.; Fernandes, L.S.; Ferreira, R.S.; de Freitas, O.; Santos, A.C. The cannabinoid beta-caryophyllene (BCP) induces neuritogenesis in PC12 cells by a cannabinoid-receptor-independent mechanism. Chem. Biol. Interact. 2017, 261, 86–95. [Google Scholar] [CrossRef]
- Mishra, D.; Bisht, G.; Mazumdar, P.M.; Sah, S.P. Chemical composition and analgesic activity of Senecio rufinervis essential oil. Pharm. Biol. 2010, 48, 1297–1301. [Google Scholar] [CrossRef]
- Segat, G.C.; Manjavachi, M.N.; Matias, D.O.; Passos, G.F.; Freitas, C.S.; Costa, R.; Calixto, J.B. Antiallodynic effect of beta-caryophyllene on paclitaxel-induced peripheral neuropathy in mice. Neuropharmacology 2017, 125, 207–219. [Google Scholar] [CrossRef]
- Katsuyama, S.; Mizoguchi, H.; Kuwahata, H.; Komatsu, T.; Nagaoka, K.; Nakamura, H.; Bagetta, G.; Sakurada, T.; Sakurada, S. Involvement of peripheral cannabinoid and opioid receptors in beta-caryophyllene-induced antinociception. Eur. J. Pain 2013, 17, 664–675. [Google Scholar] [CrossRef]
- Alberti, T.B.; Barbosa, W.L.; Vieira, J.L.; Raposo, N.R.; Dutra, R.C. (-)-beta-Caryophyllene, a CB2 Receptor-Selective Phytocannabinoid, Suppresses Motor Paralysis and Neuroinflammation in a Murine Model of Multiple Sclerosis. Int. J. Mol. Sci. 2017, 18, 691. [Google Scholar] [CrossRef]
- Rabbani, M.; Sajjadi, S.E.; Vaezi, A. Evaluation of anxiolytic and sedative effect of essential oil and hydroalcoholic extract of Ocimum basilicum L. and chemical composition of its essential oil. Res. Pharm. Sci. 2015, 10, 535–543. [Google Scholar]
- Ascari, J.; Sens, S.L.; Nunes, D.S.; Wisniewski, A., Jr.; Arbo, M.D.; Linck, V.M.; Lunardi, P.; Leal, M.B.; Elisabetsky, E. Sedative effects of essential oils obtained from Baccharis uncinella. Pharm. Biol. 2012, 50, 113–119. [Google Scholar] [CrossRef]
- Bahi, A.; Al Mansouri, S.; Al Memari, E.; Al Ameri, M.; Nurulain, S.M.; Ojha, S. beta-Caryophyllene, a CB2 receptor agonist produces multiple behavioral changes relevant to anxiety and depression in mice. Physiol. Behav. 2014, 135, 119–124. [Google Scholar] [CrossRef]
- Leonhardt, V.; Leal-Cardoso, J.H.; Lahlou, S.; Albuquerque, A.A.; Porto, R.S.; Celedonio, N.R.; Oliveira, A.C.; Pereira, R.F.; Silva, L.P.; Garcia-Teofilo, T.M.; et al. Antispasmodic effects of essential oil of Pterodon polygalaeflorus and its main constituent beta-caryophyllene on rat isolated ileum. Fundam. Clin. Pharmacol. 2010, 24, 749–758. [Google Scholar] [CrossRef]
- de Oliveira, C.C.; de Oliveira, C.V.; Grigoletto, J.; Ribeiro, L.R.; Funck, V.R.; Grauncke, A.C.; de Souza, T.L.; Souto, N.S.; Furian, A.F.; Menezes, I.R.; et al. Anticonvulsant activity of beta-caryophyllene against pentylenetetrazol-induced seizures. Epilepsy Behav. 2016, 56, 26–31. [Google Scholar] [CrossRef]
- Tchekalarova, J.; da Conceicao Machado, K.; Gomes Junior, A.L.; de Carvalho Melo Cavalcante, A.A.; Momchilova, A.; Tzoneva, R. Pharmacological characterization of the cannabinoid receptor 2 agonist, beta-caryophyllene on seizure models in mice. Seizure 2018, 57, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Dahham, S.S.; Tabana, Y.M.; Iqbal, M.A.; Ahamed, M.B.; Ezzat, M.O.; Majid, A.S.; Majid, A.M. The Anticancer, Antioxidant and Antimicrobial Properties of the Sesquiterpene beta-Caryophyllene from the Essential Oil of Aquilaria crassna. Molecules 2015, 20, 11808–11829. [Google Scholar] [CrossRef] [PubMed]
- Ambroz, M.; Bousova, I.; Skarka, A.; Hanusova, V.; Kralova, V.; Matouskova, P.; Szotakova, B.; Skalova, L. The Influence of Sesquiterpenes from Myrica rubra on the Antiproliferative and Pro-Oxidative Effects of Doxorubicin and Its Accumulation in Cancer Cells. Molecules 2015, 20, 15343–15358. [Google Scholar] [CrossRef] [PubMed]
- Legault, J.; Pichette, A. Potentiating effect of beta-caryophyllene on anticancer activity of alpha-humulene, isocaryophyllene and paclitaxel. J. Pharm. Pharmacol. 2007, 59, 1643–1647. [Google Scholar] [CrossRef]
- Jung, J.I.; Kim, E.J.; Kwon, G.T.; Jung, Y.J.; Park, T.; Kim, Y.; Yu, R.; Choi, M.S.; Chun, H.S.; Kwon, S.H.; et al. beta-Caryophyllene potently inhibits solid tumor growth and lymph node metastasis of B16F10 melanoma cells in high-fat diet-induced obese C57BL/6N mice. Carcinogenesis 2015, 36, 1028–1039. [Google Scholar] [CrossRef]
- Pavithra, P.S.; Mehta, A.; Verma, R.S. Synergistic interaction of beta-caryophyllene with aromadendrene oxide 2 and phytol induces apoptosis on skin epidermoid cancer cells. Phytomed. Int. J. Phytother. Phytopharm. 2018, 47, 121–134. [Google Scholar] [CrossRef]
- Amiel, E.; Ofir, R.; Dudai, N.; Soloway, E.; Rabinsky, T.; Rachmilevitch, S. β-Caryophyllene, a Compound Isolated from the Biblical Balm of Gilead (Commiphora gileadensis), Is a Selective Apoptosis Inducer for Tumor Cell Lines. Evid. Based Complement. Altern. 2012. [Google Scholar] [CrossRef]
- Zhou, L.; Zhan, M.L.; Tang, Y.; Xiao, M.; Li, M.; Li, Q.S.; Yang, L.; Li, X.; Chen, W.W.; Wang, Y.L. Effects of beta-caryophyllene on arginine ADP-ribosyltransferase 1-mediated regulation of glycolysis in colorectal cancer under high-glucose conditions. Int. J. Oncol. 2018, 53, 1613–1624. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Levy, R.M. The combination of beta-caryophyllene, baicalin and catechin synergistically suppresses the proliferation and promotes the death of RAW267.4 macrophages in vitro. Int. J. Mol. Med. 2016, 38, 1940–1946. [Google Scholar] [CrossRef] [PubMed]
- Youssef, D.A.; El-Fayoumi, H.M.; Mahmoud, M.F. Beta-caryophyllene protects against diet-induced dyslipidemia and vascular inflammation in rats: Involvement of CB2 and PPAR-gamma receptors. Chem. Biol. Interact. 2019, 297, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, C.; Dai, X.; Ao, Y.; Li, Y. Inhibitory effect of trans-caryophyllene (TC) on leukocyte-endothelial attachment. Toxicol. Appl. Pharmacol. 2017, 329, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Poddighe, L.; Carta, G.; Serra, M.P.; Melis, T.; Boi, M.; Lisai, S.; Murru, E.; Muredda, L.; Collu, M.; Banni, S.; et al. Acute administration of beta-caryophyllene prevents endocannabinoid system activation during transient common carotid artery occlusion and reperfusion. Lipids Health Dis. 2018, 17, 23. [Google Scholar] [CrossRef]
- Ames-Sibin, A.P.; Barizao, C.L.; Castro-Ghizoni, C.V.; Silva, F.M.S.; Sa-Nakanishi, A.B.; Bracht, L.; Bersani-Amado, C.A.; Marcal-Natali, M.R.; Bracht, A.; Comar, J.F. β-Caryophyllene, the major constituent of copaiba oil, reduces systemic inflammation and oxidative stress in arthritic rats. J. Cell. Biochem. 2018, 119, 10262–10277. [Google Scholar] [CrossRef]
- Castro Ghizoni, C.V.; Arssufi Ames, A.P.; Lameira, O.A.; Bersani Amado, C.A.; Sa Nakanishi, A.B.; Bracht, L.; Marcal Natali, M.R.; Peralta, R.M.; Bracht, A.; Comar, J.F. Anti-Inflammatory and Antioxidant Actions of Copaiba Oil Are Related to Liver Cell Modifications in Arthritic Rats. J. Cell. Biochem. 2017, 118, 3409–3423. [Google Scholar] [CrossRef]
- Arizuka, N.; Murakami, T.; Suzuki, K. The effect of beta-caryophyllene on nonalcoholic steatohepatitis. J. Toxicol. Pathol. 2017, 30, 263–273. [Google Scholar] [CrossRef]
- Yoo, H.J.; Jwa, S.K. Inhibitory effects of beta-caryophyllene on Streptococcus mutans biofilm. Arch. Oral Biol. 2018, 88, 42–46. [Google Scholar] [CrossRef]
- Cha, J.D.; Jung, E.K.; Kil, B.S.; Lee, K.Y. Chemical composition and antibacterial activity of essential oil from Artemisia feddei. J. Microbiol. Biotechnol. 2007, 17, 2061–2065. [Google Scholar]
- Yamaguchi, M.; Levy, R.M. beta-Caryophyllene promotes osteoblastic mineralization, and suppresses osteoclastogenesis and adipogenesis in mouse bone marrow cultures in vitro. Exp. Ther. Med. 2016, 12, 3602–3606. [Google Scholar] [CrossRef]
- Schmitt, D.; Levy, R.; Carroll, B. Toxicological Evaluation of beta-Caryophyllene Oil: Subchronic Toxicity in Rats. Int. J. Toxicol. 2016, 35, 558–567. [Google Scholar] [CrossRef] [PubMed]
- Tambe, Y.; Tsujiuchi, H.; Honda, G.; Ikeshiro, Y.; Tanaka, S. Gastric cytoprotection of the non-steroidal anti-inflammatory sesquiterpene, beta-caryophyllene. Planta Med. 1996, 62, 469–470. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Mysliveckova, Z.; Szotakova, B.; Spicakova, A.; Lnenickova, K.; Ambroz, M.; Kubicek, V.; Krasulova, K.; Anzenbacher, P.; Skalova, L. The inhibitory effects of beta-caryophyllene, beta-caryophyllene oxide and alpha-humulene on the activities of the main drug-metabolizing enzymes in rat and human liver in vitro. Chem. Biol. Interact. 2017, 278, 123–128. [Google Scholar] [CrossRef] [PubMed]
- (EFSA), E.F.S.A. Flavouring Group Evaluation 78 (FGE.78Rev2): Consideration of Aliphatic and alicyclic and aromatic hydrocarbons evaluated by JECFA (63rd meeting) structurally related to aliphatic and aromati hydrocarbons evaluated by EFSA in FGE.25Rev3. EFSA J. 2009, 13, 931. [Google Scholar]
- Gallelli, L.; Cione, E.; Caroleo, M.C.; Carotenuto, M.; Lagana, P.; Siniscalchi, A.; Guidetti, V. microRNAs to Monitor Pain-migraine and Drug Treatment. MicroRNA 2017, 6, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Gallelli, L.; Cione, E.; Peltrone, F.; Siviglia, S.; Verano, A.; Chirchiglia, D.; Zampogna, S.; Guidetti, V.; Sammartino, L.; Montana, A.; et al. Hsa-miR-34a-5p and hsa-miR-375 as Biomarkers for Monitoring the Effects of Drug Treatment for Migraine Pain in Children and Adolescents: A Pilot Study. J. Clin. Med. 2019, 8, 928. [Google Scholar] [CrossRef]
- Caroleo, M.C.; Carito, V.; Pingitore, A.; Perrotta, I.D.; Perri, M.; Mancuso, D.; Russo, A.; Cione, E. Human kidney podocyte cell population as a novel biological target of nerve growth factor. Growth Factors 2015, 33, 14–22. [Google Scholar] [CrossRef]
- Iacopetta, D.; Grande, F.; Caruso, A.; Mordocco, R.A.; Plutino, M.R.; Scrivano, L.; Ceramella, J.; Muia, N.; Saturnino, C.; Puoci, F.; et al. New insights for the use of quercetin analogs in cancer treatment. Future Med. Chem. 2017, 9, 2011–2028. [Google Scholar] [CrossRef]
- Rizza, P.; Pellegrino, M.; Caruso, A.; Iacopetta, D.; Sinicropi, M.S.; Rault, S.; Lancelot, J.C.; El-Kashef, H.; Lesnard, A.; Rochais, C.; et al. 3-(Dipropylamino)-5-hydroxybenzofuro[2,3-f]quinazolin-1(2H)-one (DPA-HBFQ-1) plays an inhibitory role on breast cancer cell growth and progression. Eur. J. Med. Chem. 2016, 107, 275–287. [Google Scholar] [CrossRef]
- Iacopetta, D.; Mariconda, A.; Saturnino, C.; Caruso, A.; Palma, G.; Ceramella, J.; Muia, N.; Perri, M.; Sinicropi, M.S.; Caroleo, M.C.; et al. Novel Gold and Silver Carbene Complexes Exert Antitumor Effects Triggering the Reactive Oxygen Species Dependent Intrinsic Apoptotic Pathway. ChemMedChem 2017, 12, 2054–2065. [Google Scholar] [CrossRef]
- Saturnino, C.; Barone, I.; Iacopetta, D.; Mariconda, A.; Sinicropi, M.S.; Rosano, C.; Campana, A.; Catalano, S.; Longo, P.; Ando, S. N-heterocyclic carbene complexes of silver and gold as novel tools against breast cancer progression. Future Med. Chem. 2016, 8, 2213–2229. [Google Scholar] [CrossRef] [PubMed]
- Fazio, A.; Iacopetta, D.; La Torre, C.; Ceramella, J.; Muia, N.; Catalano, A.; Carocci, A.; Sinicropi, M.S. Finding solutions for agricultural wastes: Antioxidant and antitumor properties of pomegranate Akko peel extracts and beta-glucan recovery. Food Funct. 2018, 9, 6618–6631. [Google Scholar] [CrossRef] [PubMed]
- Sinicropi, M.S.; Iacopetta, D.; Rosano, C.; Randino, R.; Caruso, A.; Saturnino, C.; Muia, N.; Ceramella, J.; Puoci, F.; Rodriquez, M.; et al. N-thioalkylcarbazoles derivatives as new anti-proliferative agents: Synthesis, characterisation and molecular mechanism evaluation. J. Enzym. Inhib. Med. Chem. 2018, 33, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Perri, M.; Pingitore, A.; Cione, E.; Vilardi, E.; Perrone, V.; Genchi, G. Proliferative and anti-proliferative effects of retinoic acid at doses similar to endogenous levels in Leydig MLTC-1/R2C/TM-3 cells. Biochim. Biophys. Acta 2010, 1800, 993–1001. [Google Scholar] [CrossRef]
- Sirignano, E.; Saturnino, C.; Botta, A.; Sinicropi, M.S.; Caruso, A.; Pisano, A.; Lappano, R.; Maggiolini, M.; Longo, P. Synthesis, characterization and cytotoxic activity on breast cancer cells of new half-titanocene derivatives. Bioorg. Med. Chem. Lett. 2013, 23, 3458–3462. [Google Scholar] [CrossRef]
- Chimento, A.; Saturnino, C.; Iacopetta, D.; Mazzotta, R.; Caruso, A.; Plutino, M.R.; Mariconda, A.; Ramunno, A.; Sinicropi, M.S.; Pezzi, V.; et al. Inhibition of human topoisomerase I and II and anti-proliferative effects on MCF-7 cells by new titanocene complexes. Bioorg. Med. Chem. 2015, 23, 7302–7312. [Google Scholar] [CrossRef]
- Saturnino, C.; Caruso, A.; Iacopetta, D.; Rosano, C.; Ceramella, J.; Muia, N.; Mariconda, A.; Bonomo, M.G.; Ponassi, M.; Rosace, G.; et al. Inhibition of Human Topoisomerase II by N,N,N-Trimethylethanammonium Iodide Alkylcarbazole Derivatives. ChemMedChem 2018, 13, 2635–2643. [Google Scholar] [CrossRef]
- Iacopetta, D.; Carocci, A.; Sinicropi, M.S.; Catalano, A.; Lentini, G.; Ceramella, J.; Curcio, R.; Caroleo, M.C. Old Drug Scaffold, New Activity: Thalidomide-Correlated Compounds Exert Different Effects on Breast Cancer Cell Growth and Progression. ChemMedChem 2017, 12, 381–389. [Google Scholar] [CrossRef]
- Sirignano, E.; Pisano, A.; Caruso, A.; Saturnino, C.; Sinicropi, M.S.; Lappano, R.; Botta, A.; Iacopetta, D.; Maggiolini, M.; Longo, P. Different 6-Aryl-Fulvenes Exert Anti-proliferative effects on Cancer Cells. Anti Cancer Agents Med. Chem. 2015, 15, 468–474. [Google Scholar] [CrossRef]
- Saturnino, C.; Popolo, A.; Ramunno, A.; Adesso, S.; Pecoraro, M.; Plutino, M.R.; Rizzato, S.; Albinati, A.; Marzocco, S.; Sala, M.; et al. Anti-Inflammatory, Antioxidant and Crystallographic Studies of N-Palmitoyl-ethanol Amine (PEA) Derivatives. Molecules 2017, 22, 616. [Google Scholar] [CrossRef]
- Augimeri, G.; Plastina, P.; Gionfriddo, G.; Rovito, D.; Giordano, C.; Fazio, A.; Barone, I.; Catalano, S.; Ando, S.; Bonofiglio, D.; et al. N-Eicosapentaenoyl Dopamine, A Conjugate of Dopamine and Eicosapentaenoic Acid (EPA), Exerts Anti-inflammatory Properties in Mouse and Human Macrophages. Nutrients 2019, 11, 2247. [Google Scholar] [CrossRef] [PubMed]
- Plastina, P.; Benincasa, C.; Perri, E.; Fazio, A.; Augimeri, G.; Poland, M.; Witkamp, R.; Meijerink, J. Identification of hydroxytyrosyl oleate, a derivative of hydroxytyrosol with anti-inflammatory properties, in olive oil by-products. Food Chem. 2019, 279, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Sinicropi, M.S.; Caruso, A.; Conforti, F.; Marrelli, M.; El Kashef, H.; Lancelot, J.C.; Rault, S.; Statti, G.A.; Menichini, F. Synthesis, inhibition of NO production and antiproliferative activities of some indole derivatives. J. Enzym. Inhib. Med. Chem. 2009, 24, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Gabriele, B.; Fazio, A.; Carchedi, M.; Plastina, P. In vitro antioxidant activity of extracts of Sybaris liquorice roots from Southern Italy. Nat. Prod. Res. 2012, 26, 2176–2181. [Google Scholar] [CrossRef] [PubMed]
- Plastina, P.; Fazio, A.; Gabriele, B. Comparison of fatty acid profile and antioxidant potential of extracts of seven Citrus rootstock seeds. Nat. Prod. Res. 2012, 26, 2182–2187. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Conditions | Effects | Possible Mechanisms | References |
|---|---|---|---|
| 50 mg/kg (i.p.) in Wistar rats (models of Alzheimer’s disease) | Neuroprotective | Decreased NO synthesis; Decreased activation of astrocytes and microglia; Reduced expression of Iba-1 e GFAP | Ojha et al. (2016) [22] |
| 10 mg/kg in rat Clone 6 (C6) microglial cells | Anti-inflammatory | Reduced production of iNOS, IL-1β, IL-6 and COX-2 | Chang et al. (2013) [24] |
| 10, 25, and 50 μM in BV2 mouse cells | Suppression of neuroinflammation due to hypoxia | NF-κB inhibition; Decreased production of NO and Prostaglandin E2 (PGE2) | Hu et al. (2017) [26] |
| 10 mg/kg in SH-SY5Y mouse cells treated with MPTP (model of Parkinson’s disease) | Neuroprotective; enhancement of motor coordination | Nrf2 activation | Viveros-Paredes et al. (2017) [30] |
| 0.5 and 1 µM in rat C6 glioma cells | Antioxidant | Nrf2 activation | Assis et al. (2014) [33] |
| 25 mg/kg twice a day (p.o.) in mice treated with paclitaxel (model of peripheral neuropathy) | Attenuation of peripheral neuropathy | mitogen-activated protein kinase (MAPK) p38/NF-κB inhibition | Segat et al. (2017) [36] |
| 2.25–18 µg (s.c.) | Strengthening of analgesic effect of morphine | CB2-R activation; Stimulation of β-endorphins release and µ-opioid receptors | Katsuyama et al. (2013) [37] |
| 50 mg/kg twice a day in mice with experimental autoimmune encephalomyelitis (model of multiple sclerosis) | Decreased pain; decreased cerebral damage; antioxidant | Decreased production of IFN-γ; Increased expression of IL-10; Increased activity of catalase, superoxide dismutase and glutathione peroxidase. | Alberti et al. (2017) [38] |
| 50, 100, and 150 mg/kg of the hydroalcoholic extract and 200 mg/kg of the essential oil from basil in Syrian mice. | Sedative | Rabbani et al. (2015) [39] | |
| 50, 100 mg/kg of the essential oil from Baccharis uncinella (p.o.) | Sedative | Ascari et al. (2012) [40] | |
| 50, 100, 150 mg/kg of the hydroalcoholic extract and 200 mg/kg of the essential oil from basil (i.p.) in Syrian mice | Anxiolytic | Rabbani et al. (2015) [39] | |
| 50 mg/kg in (C57 black 6) C57BL/6 mice (p.o.) | Anxiolytic | Activation of CB2-Rs | Bahi et al. (2014) [41] |
| 100 and 300 µg/mL of essential oil from Pterodon polygalaeflorus and 30 and 100 µg/mL of BCP in rats | Muscle relaxant | Leonhardt et al. (2010) [42] | |
| 50 mg/kg in C57BL/6 mice (p.o.) | Antidepressive | Activation of CB2-Rs | Bahi et al. (2014) [41] |
| 10, 30 and 100 mg/kg in C57BL/6 mice | Anticonvulsant | de Oliveira et al. (2016) [43] | |
| 30 mg/kg every four hours in mice | Anticonvulsant | Tchekalarova et al. (2018) [44] | |
| In HT29 and HTC116 (cell lines of colon cancer) | Antitumour | Dahham et al. (2015) [45] | |
| In PANC-1 (cell line of pancreas cancer) | Antitumour | Dahham et al. (2015) [45] | |
| 2.5, 5 and 10 μmol/L in B16F10 melanoma cells in high-fat diet-induced obese C57BL/6N mice | Reduction of obesity-related cancer risk | Jung et al. (2015) [48] | |
| Essential oil from Pamburus missionis (containing also phytolandaromadendrene oxide) in A431 and HaCaT cell lines (skin epidermoid cancer) | Antitumour | ROS production; Increased Bcl-2 Associated X protein (Bax) expression; Reduced B-cell lymphoma 2 (Bcl-2) expression; Cytochrome C release in cytoplasm; Apoptosis. | Pavithra et al. (2018) [49] |
| In BS-24-1 (mouse cell line of lymphoma) and MoFir (human lymphocytes B transformed through Epstein-Barr virus) | Antitumour | Pavithra et al. (2018) [49] | |
| In MCF-7 (breast cancer), L-929 (mouse fibroblasts), DLD-1 (colon cancer) | Increased intracellular levels of paclitaxel | Increased permeability of cell membrane; CYP3A4 inhibition | Fidyt et al. (2016) [13]; Legault et al. (2007) [47]; Nguyen et al. (2017) [64] |
| In CT26 cells transplanted in BALB/c mice and exposed to high levels of glucose (model of colorectal cancer) | Antitumour | NF-κB and arginine ADP-ribosyltransferase 1 (ART1) inhibition | Zhou et al. (2018) [51] |
| 1 or 10 µg/mL + baicalin (1, 10 µg/mL) and (+)-catechin (1, 10 µg/mL) in macrophages of RAW267.4 mouse | Reduction of macrophage proliferation | Cell cycle arrest G2/M phase; Reduction of the expression of Akt, MAPK e p65 NF-κB; Caspase-3 activation | Yamaguchi et al. (2016) [52] |
| 30 mg/kg in high-fat-fed Wistar rats (p.o.) | Antiatherogenic; Antioxidant | Activation of cannabinoid receptor 2 (CB2-R), PPAR-γ, PPAR-α, PGC1-α; Reduction of Vascular cell adhesion molecule-1 (VCAM1) expression | Youssef et al. (2019) [53] |
| 40 mg in Wistar rats (model of bilateral carotid artery occlusion and reperfusion) | Prevention of the effects induced bycarotid occlusion and reperfusion | Activation of endocannabinoid system; Increased PPAR-α expression | Poddighe et al. (2018) [55] |
| 215 and 430 mg/kg/die (p.o.) in rats (model of rheumatoid arthritis) | Reduction of oxidative stress, inflammation and edema | Radical scavenging; Nrf2 induction; Increased glutathione levels; Reduction of the expression of COX-2, cytokines and NF-κB | Ames-Sibin et al. (2018) [56] |
| 30 and 300 mg/kg (p.o.) in C57BL/6J mice fed with methionine- and choline-deficient diet (model of non-alcoholic steatohepatitis, NASH) | Reduction of liver steatosis, inflammation and fibrosis | Reduction of cytokineslevels Transforming growth factor beta (TGF-β), Nox2 and collagen; Increased levels of SOD2 and GPx1 | Arizuka et al. (2017) [58] |
| In the oral cavity for about 10 min | Bactericidal against Streptococcus mutans (tooth decay) | GlucosyltransferaseB (GtfB), GtfC and GtfD Inhibition | Yoo et al. (2018) [59] |
| 0.1–100 µM in culture of mouse bone marrow cells | Stimulates osteoblast mineralization and inhibits adipogenesis and osteoclastogenesis | PPAR-γ activation in pluripotent stem cells; Inhibits TNF-α and NF-κB in osteoclasts | Yamaguchi et al. (2016) [61] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francomano, F.; Caruso, A.; Barbarossa, A.; Fazio, A.; La Torre, C.; Ceramella, J.; Mallamaci, R.; Saturnino, C.; Iacopetta, D.; Sinicropi, M.S. β-Caryophyllene: A Sesquiterpene with Countless Biological Properties. Appl. Sci. 2019, 9, 5420. https://doi.org/10.3390/app9245420
Francomano F, Caruso A, Barbarossa A, Fazio A, La Torre C, Ceramella J, Mallamaci R, Saturnino C, Iacopetta D, Sinicropi MS. β-Caryophyllene: A Sesquiterpene with Countless Biological Properties. Applied Sciences. 2019; 9(24):5420. https://doi.org/10.3390/app9245420
Chicago/Turabian StyleFrancomano, Fabrizio, Anna Caruso, Alexia Barbarossa, Alessia Fazio, Chiara La Torre, Jessica Ceramella, Rosanna Mallamaci, Carmela Saturnino, Domenico Iacopetta, and Maria Stefania Sinicropi. 2019. "β-Caryophyllene: A Sesquiterpene with Countless Biological Properties" Applied Sciences 9, no. 24: 5420. https://doi.org/10.3390/app9245420
APA StyleFrancomano, F., Caruso, A., Barbarossa, A., Fazio, A., La Torre, C., Ceramella, J., Mallamaci, R., Saturnino, C., Iacopetta, D., & Sinicropi, M. S. (2019). β-Caryophyllene: A Sesquiterpene with Countless Biological Properties. Applied Sciences, 9(24), 5420. https://doi.org/10.3390/app9245420