Lavandula Luisieri and Lavandula Viridis Essential Oils as Upcoming Anti-Protozoal Agents: A Key Focus on Leishmaniasis

 ,
,  ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.1.1. Origin
2.1.2. Essential Oil Preparation
2.1.3. Essential Oils Analysis
2.2. Parasites and Cultures
2.3. Viability Assays
2.4. Transmission and Scanning Electron Microscopy
2.5. Flow Cytometry
2.5.1. Cell Cycle Analysis
2.5.2. Phosphatidylserine Externalization Analysis
2.5.3. Assessment of Mitochondrial Membrane Potential (MMP)
2.6. Cathepsin D Activity Assay
2.7. Cytotoxicity Evaluation in Mammalian Cells
2.8. Statistical Analysis
3. Results
3.1. Analysis of EO
3.2. Evaluation of Leishmanicidal Activity
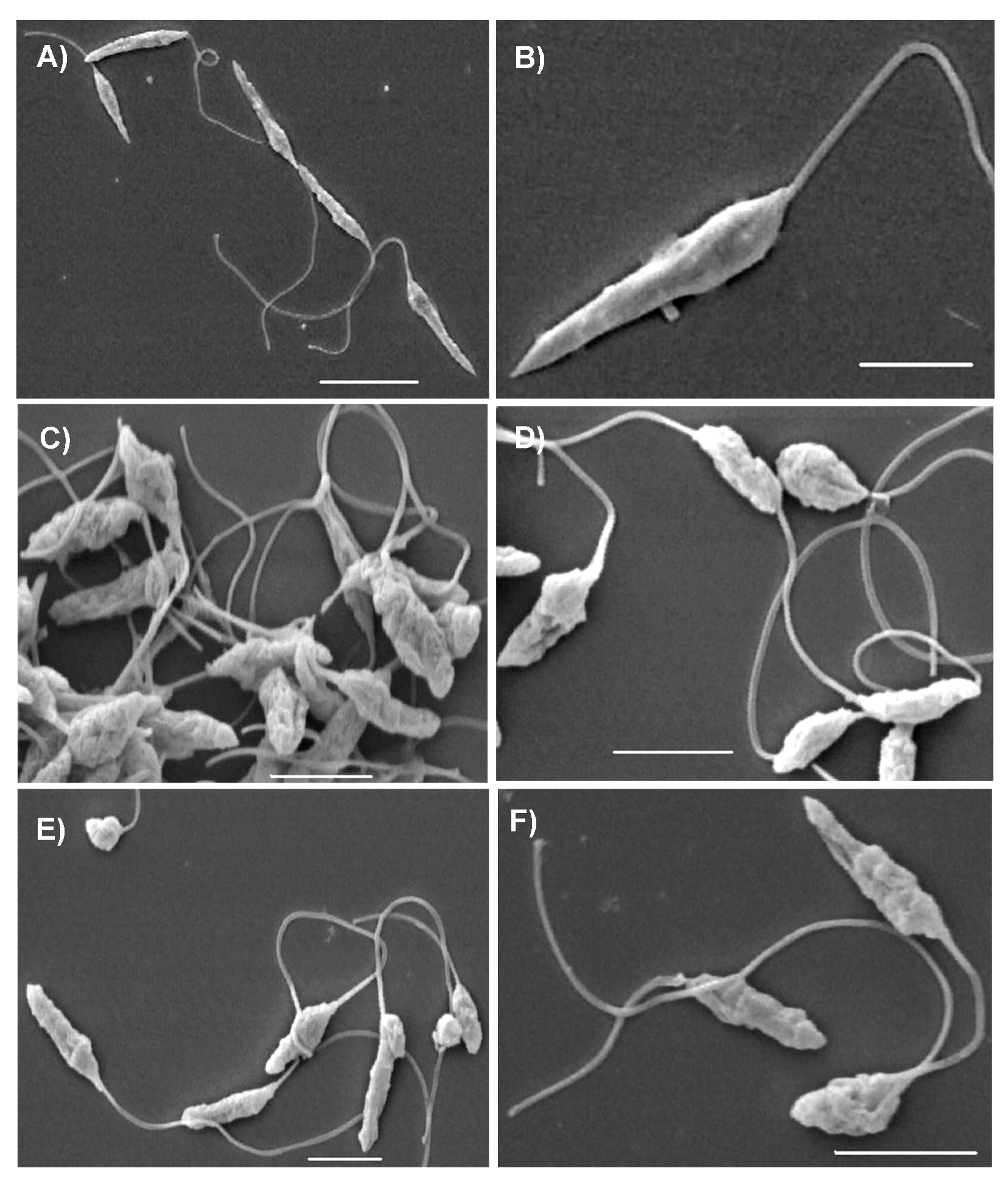
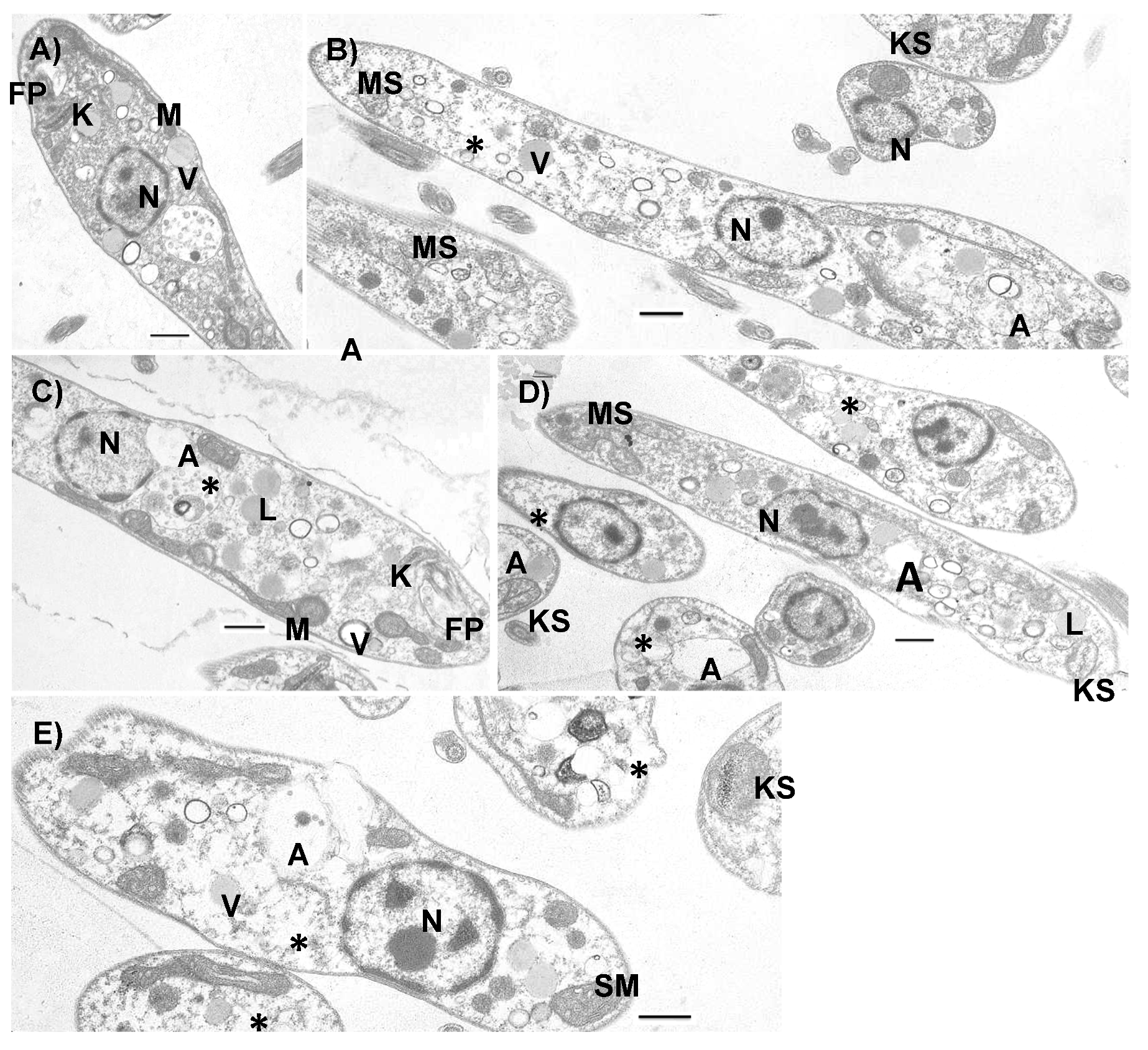
3.3. Ultrastructural Effects
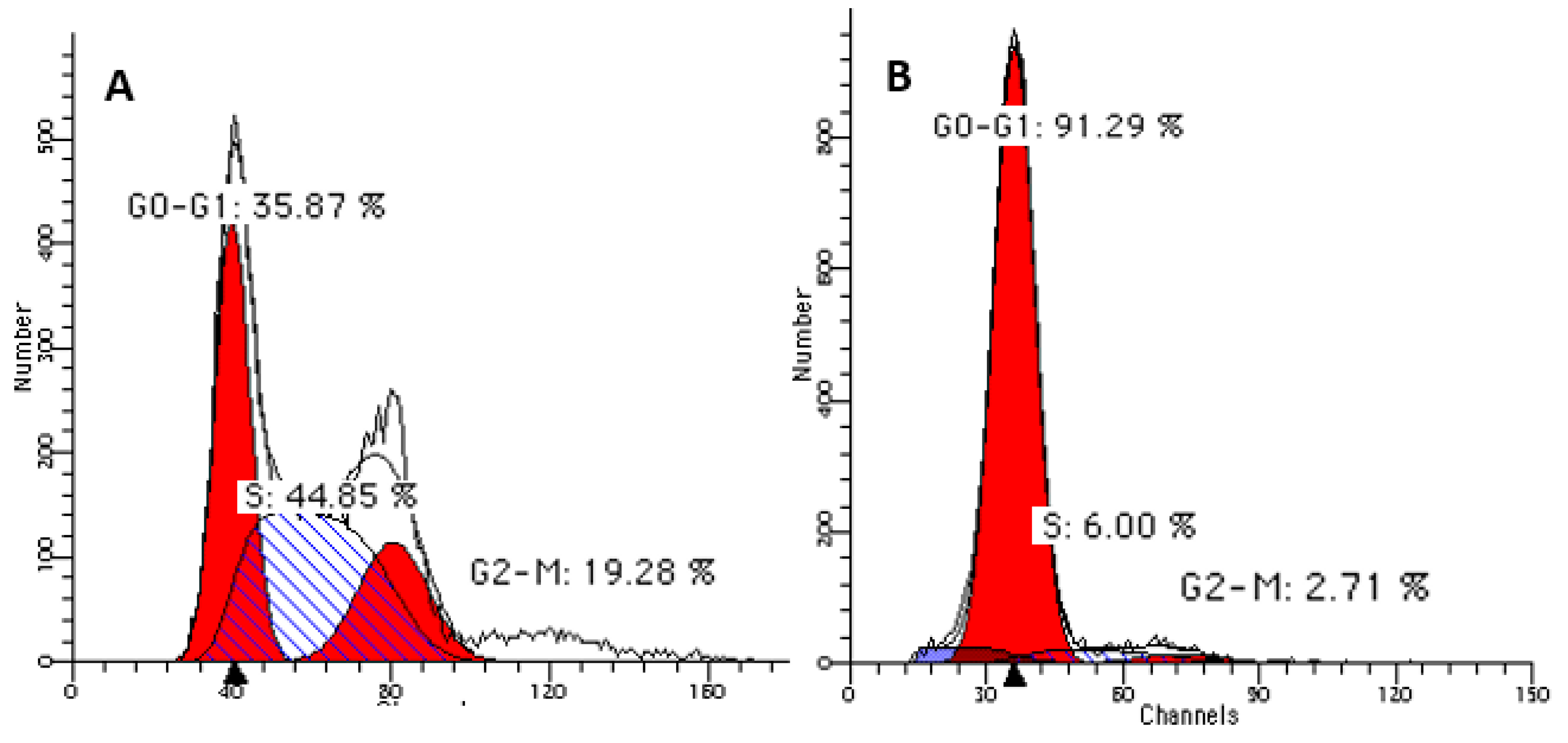
3.4. Cell-cycle Arrest at G(0)/G(1) Phase
3.5. Phosphatidylserine Externalization
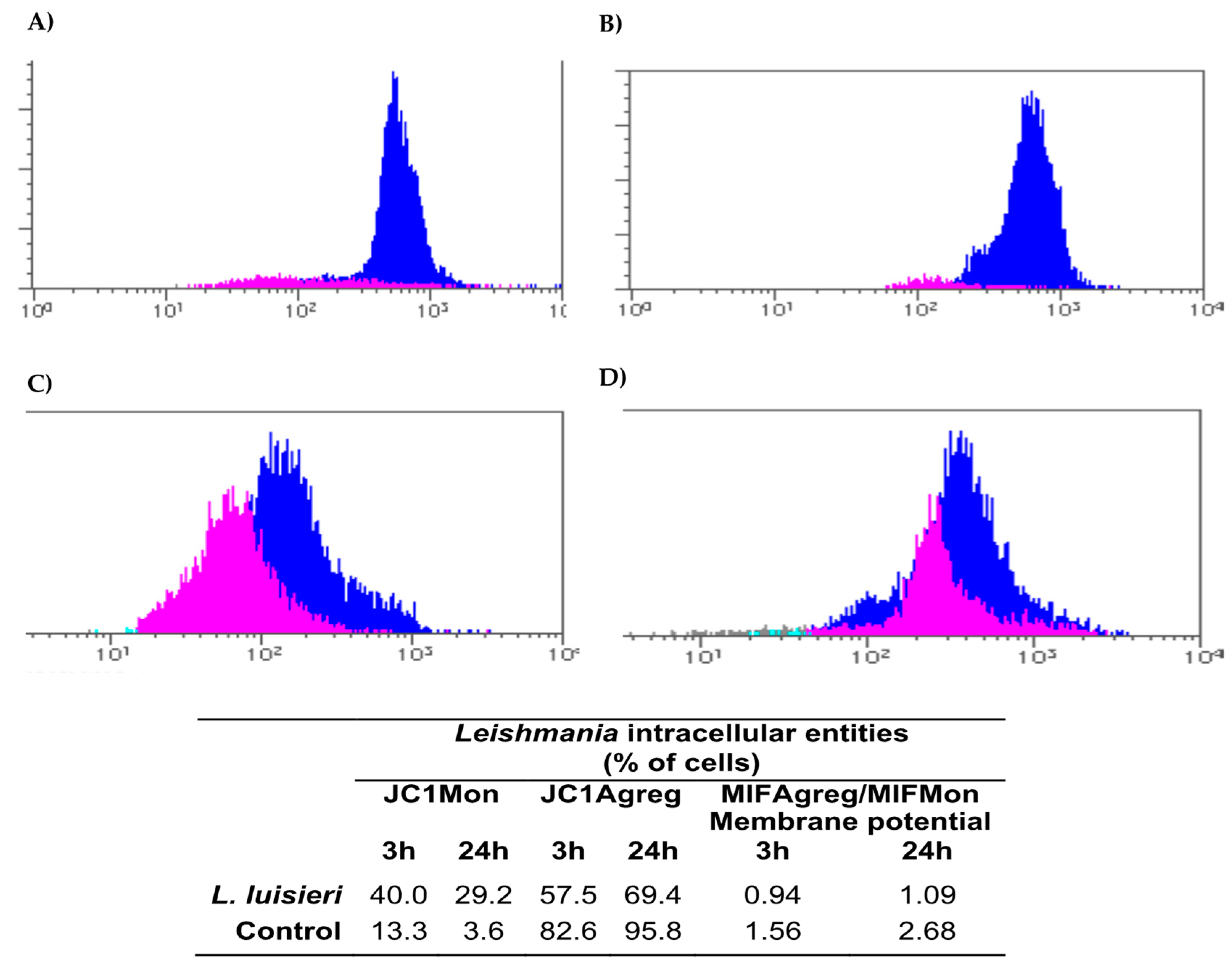
3.6. Mitochondrial Membrane Potential (MMP) Depolarization
3.7. Cathepsin D Activity Assay
3.8. Cytotoxicity Evaluation in Mammalian Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zuzarte, M.; Gonçalves, M.J.; Cavaleiro, C.; Dinis, A.M.; Canhoto, J.M.; Salgueiro, L.R. Chemical composition and antifungal activity of the essential oils of Lavandula viridis L’Hér. Chem. Biodivers 2009, 6, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Zuzarte, M.; Gonçalves, M.J.; Cavaleiro, C.; Canhoto, J.; Vale-Silva, L.; Silva, M.J.; Pinto, E.; Salgueiro, L. Chemical composition and antifungal activity of the essential oils of Lavandula viridis L’Her. J. Med. Microbiol. 2011, 60, 612–618. [Google Scholar] [CrossRef]
- Zuzarte, M.; Vale-Silva, L.; Gonçalves, M.J.; Cavaleiro, C.; Vaz, S.; Canhoto, J.; Pinto, E.; Salgueiro, L. Antifungal activity of phenolic-rich Lavandula multifida L. Essential oil. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Zuzarte, M.; Gonçalves, M.; Cruz, M.T.; Cavaleiro, C.; Canhoto, J.; Vaz, S.; Pinto, E.; Salgueiro, L. Lavandula luisieri essential oil as a source of antifungal drugs. Food Chem. 2012, 135, 1505–1510. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.; Sousa, M.D.C.; Salgueiro, L.; Cavaleiro, C. Effects of Essential Oils on the Growth of Giardia lamblia Trophozoites. Nat. Prod. Commun. 2010, 5, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.; Dinis, A.M.; Salgueiro, L.; Cavaleiro, C.; Custódio, J.B.A.; Sousa, M.D.C. Anti-Giardia activity of phenolic-rich essential oils: Effects of Thymbra capitata, Origanum virens, Thymus zygis subsp. sylvestris, and Lippia graveolens on trophozoites growth, viability, adherence, and ultrastructure. Parasitol. Res. 2010, 106, 1205–1215. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.; Santoro, G.; Sousa, M.C.; Salgueiro, L.; Cavaleiro, C.; Sousa, M.D.C.R. Activity of essential oils on the growth ofLeishmania infantumpromastigotes. Flavour Fragr. J. 2010, 25, 156–160. [Google Scholar] [CrossRef]
- Machado, M.; Dinis, A.; Salgueiro, L.; Custódio, J.B.; Cavaleiro, C.; Sousa, M.; Sousa, M.D.C.R. Anti-Giardia activity of Syzygium aromaticum essential oil and eugenol: Effects on growth, viability, adherence and ultrastructure. Exp. Parasitol. 2011, 127, 732–739. [Google Scholar] [CrossRef]
- Machado, M.; Pires, P.; Dinis, A.M.; Santos-Rosa, M.; Alves, V.; Salgueiro, L.; Cavaleiro, C.; Sousa, M.C. Monoterpenic aldehydes as potential anti-Leishmania agents: Activity of Cymbopogon citratus and citral on L. infantum, L. tropica and L. major. Exp. Parasitol. 2012, 130, 223–231. [Google Scholar] [CrossRef]
- Machado, M.; Dinis, A.; Santos-Rosa, M.; Alves, V.; Salgueiro, L.; Cavaleiro, C.; Sousa, M.; Sousa, M.D.C.R. Activity of Thymus capitellatus volatile extract, 1,8-cineole and borneol against Leishmania species. Veter Parasitol. 2014, 200, 39–49. [Google Scholar] [CrossRef]
- Wiley, R. NIST 05 MS Spectra, 8th ed.; Revision 2005 D.06.00. s.l.; NIST Mass Spectral Library: Gaithersburg, MD, USA, 2007. [Google Scholar]
- Joulain, D.; Konig, W.A. The atlas of spectral data of sesquiterpene hydrocarbons. J. Nat. Prod. 1998, 62, 1212–1213. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; pp. 1–804. [Google Scholar]
- Monzote, L.; Montalvo, A.M.; Scull, R.; Miranda, M.; Abreu, J. Combined effect of the essential oil from Chenopodium ambrosioides and antileishmanial drugs on promastigotes of Leishmania amazonensis. Rev. Inst. Med. Trop. São Paulo 2007, 49, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Natera, S.; Machuca, C.; Padrón-Nieves, M.; Romero, A.; Díaz, E.; Ponte-Sucre, A. Leishmania spp.: Proficiency of drug-resistant parasites. Int. J. Antimicrob. Agents 2007, 29, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Croft, S.L.; Sundar, S.; Fairlamb, A.H. Drug resistance in Leishmaniasis. Clin. Microbiol. Rev. 2006, 19, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils–A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Simões, M.; Bennett, R.N.; Rosa, E.A.S. Understanding antimicrobial activities of phytochemicals against multidrug resistant bacteria and biofilms. Nat. Prod. Rep. 2009, 26, 746. [Google Scholar] [CrossRef] [PubMed]
- Sen, N.; Das, B.B.; Ganguly, A.; Mukherjee, T.; Tripathi, G.; Bandyopadhyay, S.; Rakshit, S.; Sen, T.; Majumder, H.K. Camptothecin induced mitochondrial dysfunction leading to programmed cell death in unicellular hemoflagellate Leishmania donovani. Cell Death Differ. 2004, 11, 924–936. [Google Scholar] [CrossRef]
- Pedroso, R.B.; Ueda-Nakamura, T.; Filho, B.P.D.; Cortez, D.A.G.; Cortez, L.E.R.; Morgado-Diaz, J.A.; Nakamura, C.V. Biological activities of essential oil obtained from Cymbopogon citratus on Crithidia deanei. Acta Protozool. 2006, 45, 231–240. [Google Scholar]
- Santoro, G.F.; Cardoso, M.G.; Guimarães, L.G.L.; Freire, J.M.; Soares, M.J.; Soares, M. Anti-proliferative effect of the essential oil of Cymbopogon citratus (DC) Stapf (lemongrass) on intracellular amastigotes, bloodstream trypomastigotes and culture epimastigotes of Trypanosoma cruzi (Protozoa: Kinetoplastida). Parasitology 2007, 134, 1649–1656. [Google Scholar] [CrossRef]
- Oliveira, V.C.S.; Moura, D.M.S.; Lopes, J.A.D.; De Andrade, P.P.; Da Silva, N.H.; Figueiredo, R.C.B.Q. Effects of essential oils from Cymbopogon citratus (DC) Stapf., Lippia sidoides Cham., and Ocimum gratissimum L. on growth and ultrastructure of Leishmania chagasi promastigotes. Parasitol. Res. 2009, 104, 1053–1059. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Rodrigues, J.C.F.; Attias, M.; Rodriguez, C.; Urbina, J.A.; De Souza, W.; Souza, W.D. Ultrastructural and Biochemical Alterations Induced by 22,26-Azasterol, a Δ24(25)-Sterol Methyltransferase Inhibitor, on Promastigote and Amastigote Forms of Leishmania amazonensis. Antimicrob. Agents Chemother. 2002, 46, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Lazardi, K.; Urbina, J.A.; De Souza, W. Ultrastructural alterations induced by two ergosterol biosynthesis inhibitors, ketoconazole and terbinafine, on epimastigotes and amastigotes of Trypanosoma (Schizotrypanum) cruzi. Antimicrob. Agents Chemother. 1990, 34, 2097–2105. [Google Scholar] [CrossRef] [PubMed]
- Lorente, S.O.; Rodrigues, J.C.F.; Jimenez, C.J.; Joyce-Menekse, M.; Rodrigues, C.; Croft, S.L.; Yardley, V.; De Luca-Fradley, K.; Ruiz-Pérez, L.M.; Urbina, J.; et al. Novel Azasterols as Potential Agents for Treatment of Leishmaniasis and Trypanosomiasis. Antimicrob. Agents Chemother. 2004, 48, 2937–2950. [Google Scholar] [CrossRef] [PubMed]
- Brenzan, M.A.; Nakamura, C.V.; Filho, B.P.D.; Ueda-Nakamura, T.; Young, M.C.M.; Cortez, D.A.G.; Young, M.C.M. Antileishmanial activity of crude extract and coumarin from Calophyllum brasiliense leaves against Leishmania amazonensis. Parasitol. Res. 2007, 101, 715–722. [Google Scholar] [CrossRef]
- Rosa, M.D.S.S.; Bizzo, H.R.; Rodrigues, I.D.A.; Soares, R.M.A.; Souto-Padrón, T.; Alviano, C.S.; Lopes, A.H.C.S.; Mendonça-Filho, R.R. Antileishmanial Activity of a Linalool-Rich Essential Oil from Croton cajucara. Antimicrob. Agents Chemother. 2003, 47, 1895–1901. [Google Scholar] [CrossRef]
- Ueda-Nakamura, T.; Mendonça-Filho, R.R.; Morgado-Díaz, J.A.; Maza, P.K.; Filho, B.P.D.; Cortez, D.A.G.; Alviano, D.S.; Rosa, M.D.S.S.; Lopes, A.H.C.; Alviano, C.S.; et al. Antileishmanial activity of Eugenol-rich essential oil from Ocimum gratissimum. Parasitol. Int. 2006, 55, 99–105. [Google Scholar] [CrossRef]
- Arnoult, D.; Akarid, K.; Grodet, A.; Petit, P.X.; Estaquier, J.; Ameisen, J.C. On the evolution of programmed cell death: Apoptosis of the unicellular eukaryote Leishmania major involves cysteine proteinase activation and mitochondrion permeabilization. Cell Death Differ. 2002, 9, 65–81. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Lavandula luisieri (%) | Lavandula viridis (%) |
|---|---|---|
| Monoterpene hydrocarbons | 4.7 | 17.3 |
| Oxygen containing monoterpenes | 75.7 | 58.3 |
| Sesquiterpene hydrocarbons | 2.4 | 18.6 |
| Oxygen containing sesquiterpenes | 3.0 | - |
| Others | 1.2 | 1.9 |
| Total identified | (49) 86.9 | (38) 96.1 |
| Major compounds (>2.0%) | α-Pinene (2.3) 1,8-cineole (18.9) Linalool (3.1) Necrodane derivatives (36.0) Lavandulyl acetate (7.2) | α-Pinene (9.2) Camphene (2.7) 1,8-cineole (29.7) Linalool (9.0) Camphor (10.0) Borneol (2.7) Z-α-bisabolene (6.3) Selina-3,7(11)-diene (6.6) |
| L. infantum IC50 μg/mL (CI) * | L. tropica IC50 μg/mL (CI) * | L. major IC50 μg/mL (CI) * | |
|---|---|---|---|
| L. luisieri | 63 (52–77) | 38 (33–45) | 31 (25–38) |
| L. viridis | 263 (248–279) | >400 | >400 |
| α-pinene | 161 (149–175) | >400 | >400 |
| 1,8-cineole | >400 | >400 | >400 |
| Borneol | >400 | >400 | >400 |
| Linalool | >400 | >400 | >400 |
| Leishmania Promastigotes (% of Cells) 24 h | |||
|---|---|---|---|
| Anexine | PI | Anexine/PI | |
| L. luisieri | 17.5 | 4 | 10 |
| Control | 3.3 | 1.1 | 4.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machado, M.; Martins, N.; Salgueiro, L.; Cavaleiro, C.; Sousa, M.C. Lavandula Luisieri and Lavandula Viridis Essential Oils as Upcoming Anti-Protozoal Agents: A Key Focus on Leishmaniasis. Appl. Sci. 2019, 9, 3056. https://doi.org/10.3390/app9153056
Machado M, Martins N, Salgueiro L, Cavaleiro C, Sousa MC. Lavandula Luisieri and Lavandula Viridis Essential Oils as Upcoming Anti-Protozoal Agents: A Key Focus on Leishmaniasis. Applied Sciences. 2019; 9(15):3056. https://doi.org/10.3390/app9153056
Chicago/Turabian StyleMachado, Marisa, Natália Martins, Lígia Salgueiro, Carlos Cavaleiro, and Maria C. Sousa. 2019. "Lavandula Luisieri and Lavandula Viridis Essential Oils as Upcoming Anti-Protozoal Agents: A Key Focus on Leishmaniasis" Applied Sciences 9, no. 15: 3056. https://doi.org/10.3390/app9153056
APA StyleMachado, M., Martins, N., Salgueiro, L., Cavaleiro, C., & Sousa, M. C. (2019). Lavandula Luisieri and Lavandula Viridis Essential Oils as Upcoming Anti-Protozoal Agents: A Key Focus on Leishmaniasis. Applied Sciences, 9(15), 3056. https://doi.org/10.3390/app9153056