Lipid and Carotenoid Production by Rhodotorula glutinis with a Combined Cultivation Mode of Nitrogen, Sulfur, and Aluminium Stress


Abstract
1. Introduction
2. Materials and Methods
2.1. The Microorganism, Media Preparation, and Cultivation
2.2. Effect of Different Al2(SO4)3 Concentration on Growth, and Lipid and Carotenoid Production
2.3. Detection of Dry Cell Weight and Residual Sugar in Culture Media
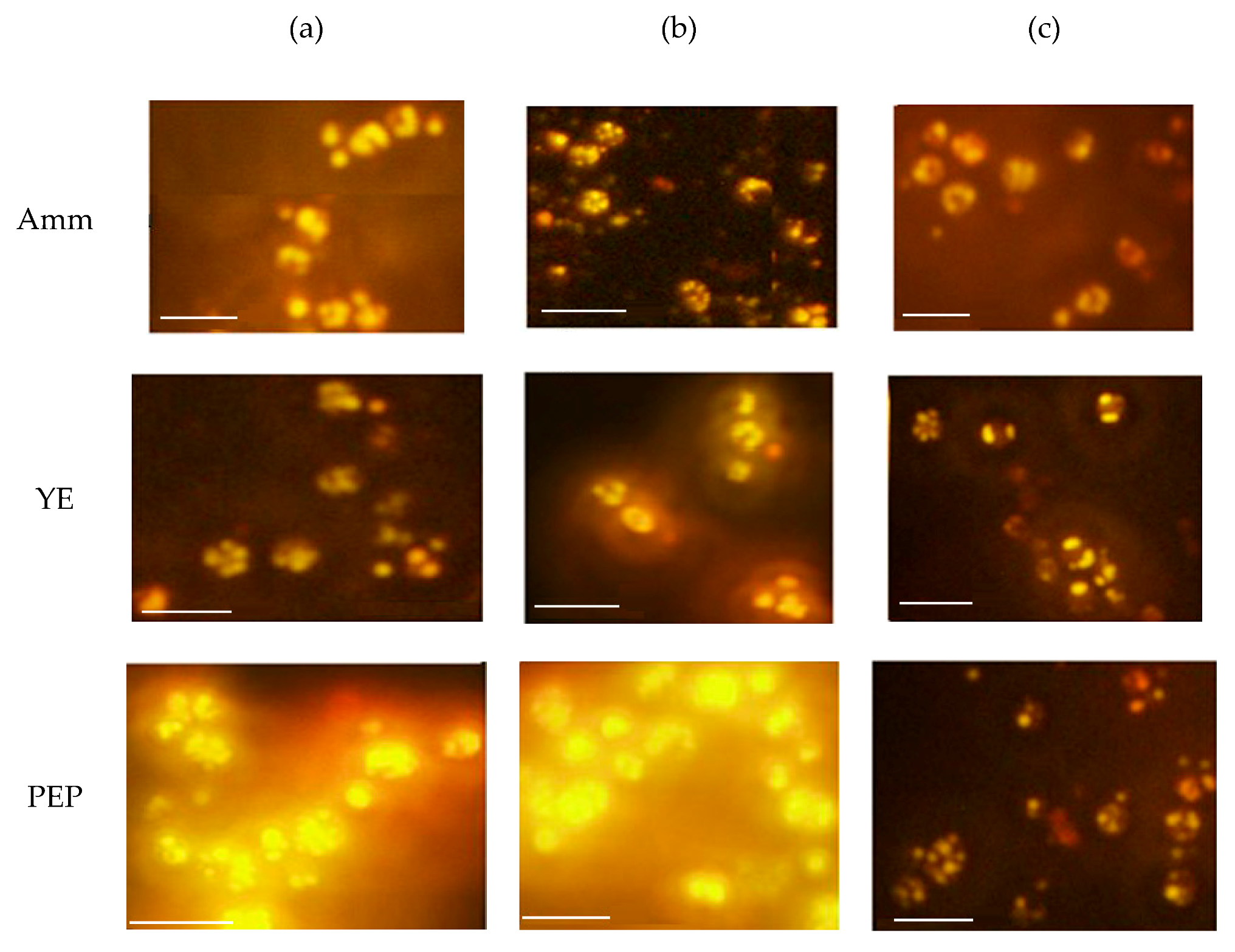
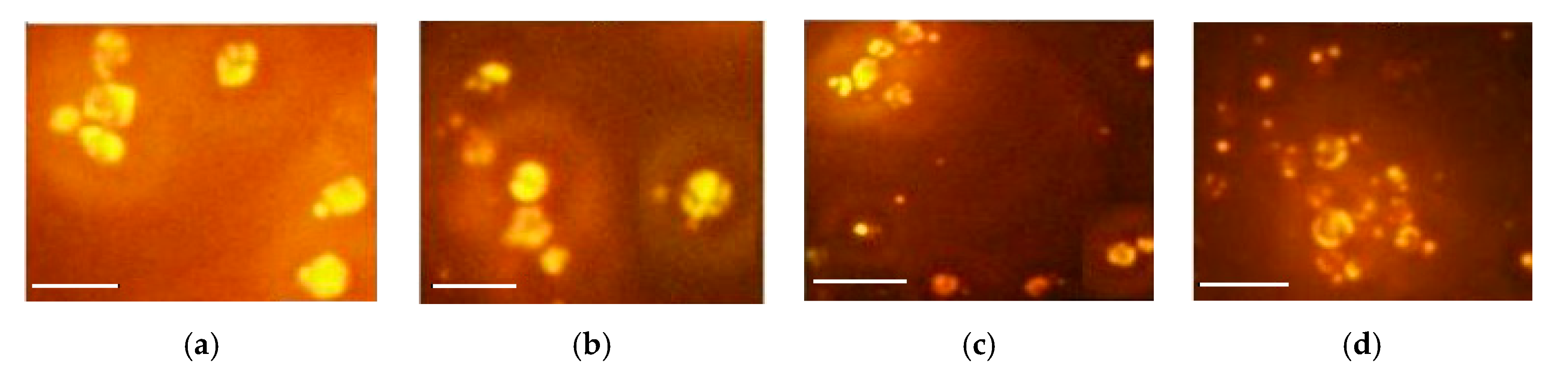
2.4. Nile Red Staining and Fluorescence Microscopy of Cultured R. glutinis
2.5. Total Lipid Detection of Cultured R. glutinis
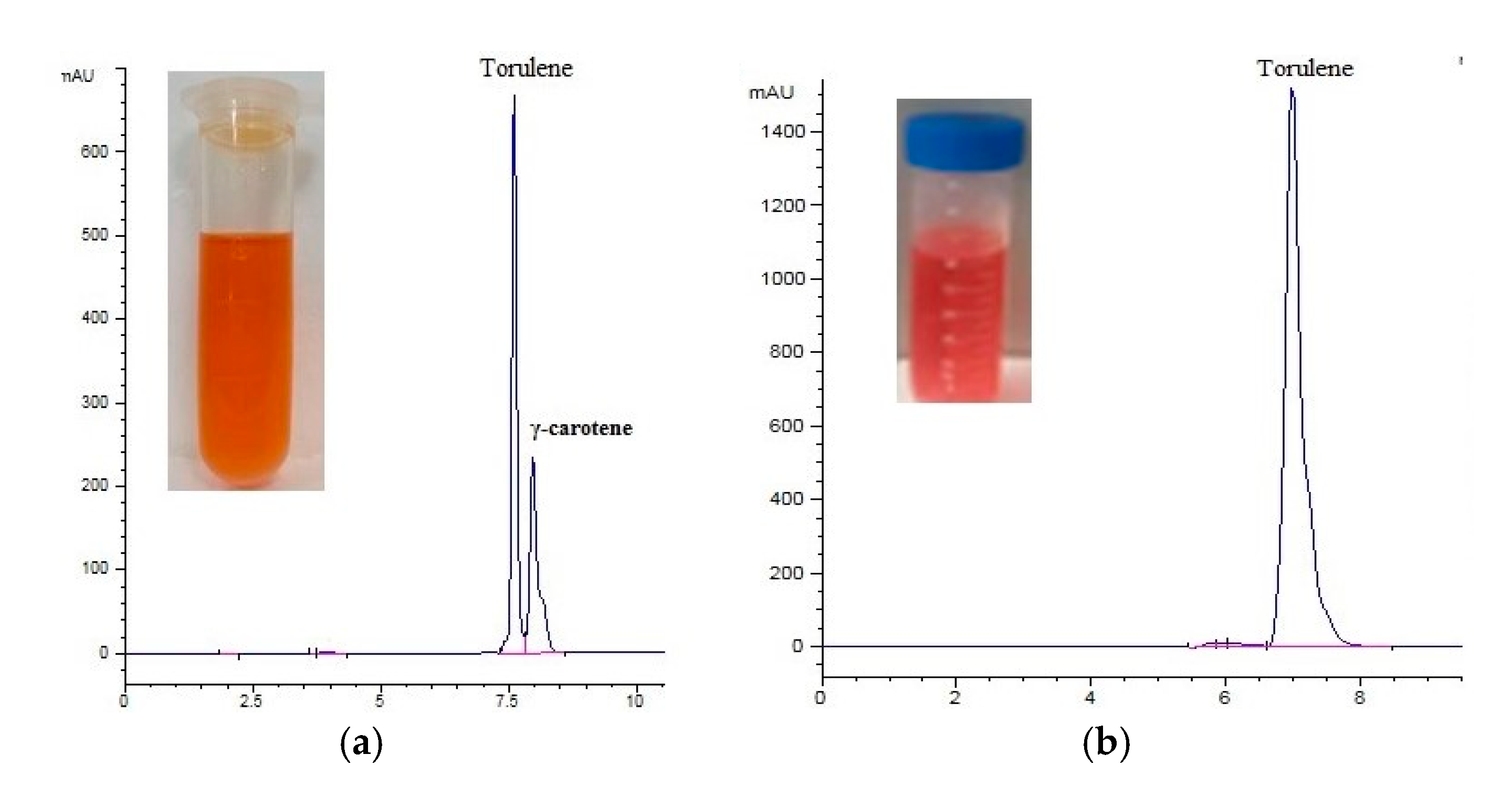
2.6. Extraction and Assay of Carotenoid
2.7. The Kinetics of Lipid and Carotenoid Biosynthesis
2.8. Statistical Analysis
3. Results
3.1. Effect of Different Nitrogen Sources and Different C/N Ratios on Cell Biomass, and Lipid and Carotenoid Production by R. glutinis
3.2. Effect of Different C/S Ratios on Biomass, Lipid, and Carotenoid Production
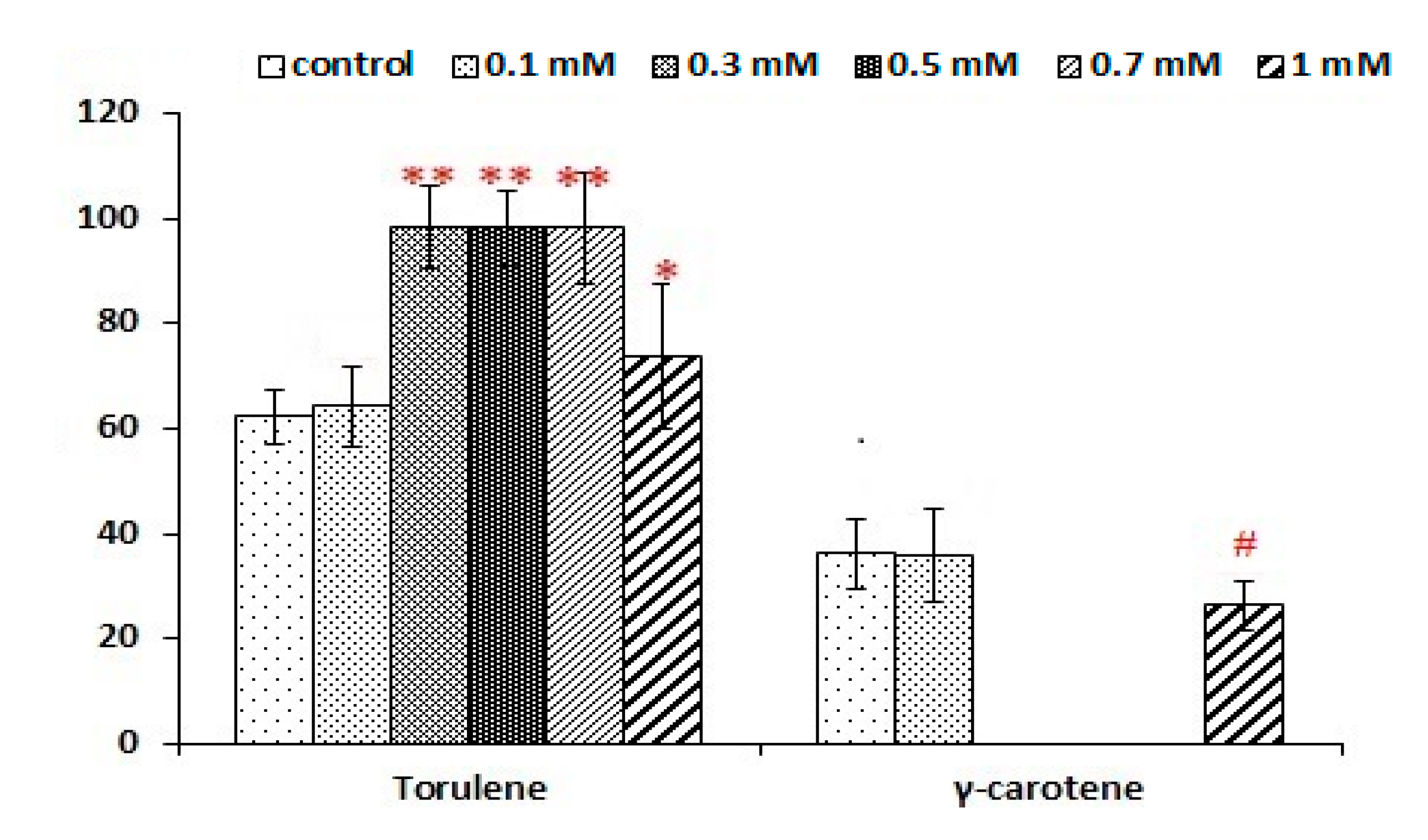
3.3. The Carotenoid Profile from Different C/N, C/S Ratios and Nitrogen Sources Treatments
3.4. Impact of Aluminium Sulfate on Lipogenesis and Carotenoid Production
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- BCC Research. The Global Market for Carotenoids. Available online: https www.bccresearch.com/market-research/food-and-beverage/the-global-market-for-carotenoids.html (accessed on 13 June 2018).
- Mannazzu, I.; Landolfo, S.; Lopes, T.; Buzzini, P. Red yeasts and carotenoid production: Outlining a future for non-conventional yeasts of biotechnological interest. World J. Microbiol. Biotechnol. 2015, 31, 1665–1673. [Google Scholar] [CrossRef]
- Mussagy, C.U.; Winterburn, J.; Santos-Ebinuma, V.C.; Pereira, J.F.B. Production and extraction of carotenoids produced by microorganisms. Appl. Microbiol. Biotechnol. 2019, 103, 1095–1114. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Schmidt-Dannert, C. Metabolic engineering towards biotechnological production of carotenoids in microorganisms. Appl. Microbiol. Biotechnol. 2002, 60, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kot, A.M.; Błażejak, S.; Kurcz, A.; Gientka, I.; Kieliszek, M. Rhodotorula glutinis—Potential source of lipids, carotenoids, and enzymes for use in industries. Appl. Microbiol. Biotechnol. 2016, 100, 6103–6117. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Ding, Y.; Gong, Z.; Yang, L.; Zhang, S.; Zhang, C.; Lin, X.; Shen, H.; Zou, H.; Xie, Z.; et al. Dynamics of the lipid droplet proteome of the oleaginous yeast Rhodosporidium Toruloides. Eukaryot. Cell 2015, 14, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Sawangkeaw, R.; Ngamprasertsith, S. A review of lipid-based bio-masses as feedstocks for biofuels production. Renew. Sustain. Energy Rev. 2013, 25, 97–108. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, X.; Tan, T. The effect of amino acids on lipid production and nutrient removal by Rhodotorula glutinis cultivation in starch wastewater. Bioresour. Technol. 2016, 218, 712–717. [Google Scholar] [CrossRef]
- Schneider, T.; Graeff-Hönninger, S.; French, W.T.; Hernandez, R.; Merkt, N.; Claupein, W.; Pham, P. Lipid and carotenoid production by oleaginous red yeast Rhodotorula glutinis cultivated on brewery effluents. Energy 2013, 61, 34–43. [Google Scholar] [CrossRef]
- Tkáčová, J.; Čaplová, J.; Klempová, T.; Čertík, M. Correlation between lipid and carotenoid synthesis in torularhodin-producing Rhodotorula glutinis. Ann. Microbiol. 2017, 67, 541–551. [Google Scholar] [CrossRef]
- Buzzini, P.; Innocenti, M.; Turchetti, B.; Libkind, D.; Broock, M.; Mulinacci, N. Carotenoid profiles of yeasts belonging to the genera Rhodotorula, Rhodosporidium, Sporobolomyces, and Sporidiobolus. Can. J. Microbiol. 2007, 1031, 1024–1031. [Google Scholar] [CrossRef]
- Braunwald, T.; Schwemmlein, L.; Graeff-Hönninger, S.; French, W.T.; Hernandez, R.; Holmes, W.E.; Claupein, W. Effect of different C/N ratios on carotenoid and lipid production by Rhodotorula glutinis. Appl. Microbiol. Biotechnol. 2013, 97, 6581–6588. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhang, N.; Li, B.; Xu, Q.; Song, J.; Wei, N. Increased torulene accumulation in red yeast Sporidiobolus pararoseus NGR as stress response to high salt conditions. Food Chem. 2017, 15, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Buzzini, P.; Martini, A.; Gaetani, M.; Turchetti, B.; Maria, U.; Davoli, P.; Emilia, R. Optimisation of carotenoid production by Rhodotorula graminis DBVPG 7021 as a function of trace element concentration by means of response surface analysis. Enzyme Microb. Technol. 2005, 36, 687–692. [Google Scholar] [CrossRef]
- Han, M.; Xu, Z.; Du, C.; Qian, H. Effects of nitrogen on the lipid and carotenoid accumulation of oleaginous yeast Sporidiobolus pararoseus. Bioprocess Biosyst. Eng. 2016, 39, 1425–1433. [Google Scholar] [CrossRef]
- Zoz, L.; Carvalho, J.C.; Soccol, V.T.; Casagrande, T.C.; Cardoso, L. Torularhodin and torulene: Bioproduction, properties and prospective applications in food and cosmetics–A review. Braz. Arch. Biol. Technol. 2015, 58, 278–288. [Google Scholar] [CrossRef]
- Kot, A.M.; Błażejak, S.; Gientka, I.; Kieliszek, M.; Bryś, J. Torulene and torularhodin: “new’’ fungal carotenoids for industry? Microbl. Cell Fact. 2018, 100, 6103–6117. [Google Scholar] [CrossRef] [PubMed]
- Marova, I.; Carnecka, M.; Halienova, A.; Breierova, E.; Koci, R. Production of carotenoid-ergosterol-supplemented biomass by red yeast Rhodotorula glutinis grown under external stress. Food Technol. Biotechnol. 2010, 48, 56–61. [Google Scholar]
- Bhosale, P.B.; Gadre, R.V. Production of β-carotene by a mutant of Rhodotorula glutinis. Appl. Microbiol. Biotechnol. 2001, 55, 423–427. [Google Scholar] [CrossRef]
- Dias, C.; Sousa, S.; Caldeira, J.; Reis, A.; Lopes da Silva, T. New dual-stage pH control fed-batch cultivation strategy for the improvement of lipids and carotenoids production by the red yeast Rhodosporidium toruloides NCYC 921. Bioresour. Technol. 2015, 189, 309–318. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, X.; Tan, T. Lipid and carotenoid production by Rhodotorula glutinis under irradiation/high-temperature and dark/low-temperature cultivation. Bioresour. Technol. 2014, 157, 149–153. [Google Scholar] [CrossRef]
- Kot, A.M.; Błażejak, S.; Kieliszek, M.; Gientka, I.; Bryś, J. Simultaneous Production of Lipids and Carotenoids by the Red Yeast Rhodotorula from Waste Glycerol Fraction and Potato Wastewater. Appl. Biochem. Biotechnol. 2019, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Somashekar, D.; Joseph, R. Inverse relationship between carotenoid and lipid formation in Rhodotorula gracilis according to the C/N ratio of the growth medium. World J. Microbiol. Biotechnol. 2000, 16, 491–493. [Google Scholar] [CrossRef]
- Ratledge, C.; Wynn, J.P. The biochemistry and molecular biology of lipid accumulation in oleaginous microorganisms. Adv. Appl. Microbiol. 2002, 51, 1–51. [Google Scholar] [PubMed]
- Zhu, Z.; Zhang, S.; Liu, H.; Shen, H.; Lin, X.; Yang, F.; Zhou, Y.J.; Jin, G.; Ye, M.; Zou, H.; et al. A multi-omic map of the lipid-producing yeast Rhodosporidium toruloides. Nat. Commun. 2012, 3, 1112. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhao, X.; Shen, H.; Wang, Q.; Zhao, Z.K. Microbial lipid production by Rhodosporidium toruloides under sulfate-limited conditions. Bioresour. Technol. 2011, 102, 1803–1807. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Morris, H.; Cronin, M.T. Metals, toxicity and oxidative stress. Curr. Med. Chem. 2005, 12, 1161–1208. [Google Scholar] [CrossRef]
- Porwol, T.; Ehleben, W.; Zierold, K.; Fandrey, J.; Acker, H. The influence of nickel and cobalt on putative members of the oxygen-sensing pathway of erythropoietin-producing HepG2 cells. Eur. J. Biochem. 1998, 256, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Rainbow, P.; Black, W. Cadmium, zinc and the uptake of calcium by two crabs, Carcinus maenasandEriocheir sinensis. Aquat. Toxicol. 2005, 72, 45–65. [Google Scholar] [CrossRef]
- Qiu, J.; Xie, Z.; Wang, W. Effects of calcium on the uptake and elimination of cadmium and zinc in Asiatic clams. Arch. Environ. Contam. Toxicol. 2005, 48, 278–287. [Google Scholar] [CrossRef]
- Belcastro, M.; Marino, T.; Russo, N.; Toscano, M. Interaction of cysteine with Cu2+ and group IIb (Zn2+,Cd2+,Hg2+) metal cations: A theoretical study. J. Mass Spectrom. 2005, 40, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Stohs, S.J. The role of free radicals in toxicity and disease. J. Basic Clin. Physiol. Pharmacol. 1995, 6, 205–228. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Shi, X. Intracellular signal transduction of cells in response to carcinogenic metals. Crit. Rev. Oncol. Hematol. 2002, 42, 105–121. [Google Scholar] [CrossRef]
- Dandrea, T.; Hellmold, H.; Jonsson, C.; Zhivotovsky, B.; Hofer, T.; Wärngård, L.; Cotgreave, I. The transcriptosomal response of human A549 lung cells to a hydrogen peroxide-generating system: Relationship to DNA damage, cell cycle arrest, and caspase activation. Free Radic. Biol. Med. 2004, 36, 881–896. [Google Scholar] [CrossRef]
- Beyersmann, D.; Hartwig, A. Carcinogenic metal compounds: Recent insight into molecular and cellular mechanisms. Arch. Toxicol. 2008, 82, 493–512. [Google Scholar] [CrossRef]
- Kochian, L.V. Cellular mechanisms of aluminium toxicity and resistance in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 237–260. [Google Scholar] [CrossRef]
- Cakmak, I.; Horst, W. Effect of aluminium on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiol. Plant. 1991, 83, 463–468. [Google Scholar] [CrossRef]
- Gutteridge, J.; Quinlan, G.; Clark, I.; Halliwall, B. Aluminium salts accelerate peroxidation of membrane lipids stimulated by iron salts. Biochim. Biophys. Acta 1985, 835, 441–447. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Kimura, K.; Yamaoka, M.; Kamisaka, Y. Rapid estimation of lipids in oleaginous fungi and yeasts using Nile red fluorescence. J. Microbiol. Methods 2004, 56, 331–338. [Google Scholar] [CrossRef]
- Mishra, S.; Suh, W.; Farooq, W.; Moon, M.; Shrivastav, A.; Park, M.S.; Yang, J. Rapid quantification of microalgal lipids in aqueous medium by a simple colorimetric method. Bioresour. Technol. 2014, 155, 330–333. [Google Scholar] [CrossRef]
- Frengova, G.; Sirnova, E.; Pavlova, K.; Beshkova, D. Formation of Carotenoids by Rhodotorula glutinis in whey ultrafiltrate. Biotechnol. Bioeng. 1994, 44, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Weber, R.W.; Anke, H.; Davoli, P. Simple method for the extraction and reversed-phase high-performance liquid chromatographic analysis of carotenoid pigments from red yeasts (Basidiomycota, Fungi). J. Chromatogr. A 2007, 1145, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Certik, M.; Shimizu, S. Kinetic analysis of oil biosynthesis by an arachidonic acid-producing fungus, Mortierella alpina 1S-4. Appl. Microbiol. Biotechnol. 2000, 54, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Amaretti, A.; Raimondi, S.; Sala, M.; Roncaglia, L.; De Lucia, M.; Leonardi, A.; Rossi, M. Single cell oils ofthe cold-adapted oleaginous yeast Rhodotorula glacialis DBVPG 4785. Microb. Cell Fact. 2010, 9, 73. [Google Scholar] [CrossRef]
- Jiru, T.M.; Groenewald, M.; Pohl, C.; Steyn, L.; Kiggundu, N.; Abate, D. Optimization of cultivation conditions for biotechnological production of lipid by Rhodotorula kratochvilovae(syn, Rhodosporidium kratochvilovae) SY89 for biodiesel preparation. 3 Biotech 2017, 7, 145. [Google Scholar] [CrossRef]
- Zhu, L.; Zong, M.; Wu, H. Efficient lipid production with Trichosporon fermentans and its use for biodiesel preparation. Bioresour. Technol. 2008, 99, 7881–7885. [Google Scholar] [CrossRef] [PubMed]
- Libkind, D.; Brizzio, S.; Van Broock, M. Rhodotorula mucilaginosa, a carotenoid producing yeast strain from a Patagonian high-altitude Lake. Folia Microbiol. 2004, 49, 19–25. [Google Scholar] [CrossRef]
- Bhosale, P. Environmental and cultural stimulants in the production of carotenoids from microorganisms. Appl. Microbiol. Biotechnol. 2004, 63, 351–361. [Google Scholar] [CrossRef]
- El-Banna, A.; Abd El-Razek, A.; El-Mahdy, A. Some Factors Affecting the Production of Carotenoids by Rhodotorula glutinis var. glutinis. Food Nutr. Sci. 2012, 3, 64–71. [Google Scholar] [CrossRef]
- Cutzu, R.; Clemente, A.; Reis, A.; Lopes, T. Assessment of β-carotene content, cell physiology and morphology of the yellow yeast Rhodotorula glutinis mutant 400A15 using flow cytometry. J. Ind. Microbiol. Biotechnol. 2013, 40, 865–875. [Google Scholar] [CrossRef]
- Wang, C.; WanG, C.I.; Zhao, X.Q.; Chen, F.; Lan, P.; Shen, R.F. Proteomic analysis of a high aluminum tolerant yeast Rhodotorula taiwanensis RS1 in response to aluminum stress. Biochim. Biophys. Acta 2013, 1834, 1969–1975. [Google Scholar] [CrossRef]
- Ren, H.; Liu, B.; Kong, F.; Zhao, L.; Xie, G.; Ren, N. Enhanced lipid accumulation of green microalga Scenedesmus sp. by metal ions and EDTA addition. Bioresour. Technol. 2014, 169, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Cao, J.; Xing, G.; Yuan, H. Lipid production combined with biosorption and bioaccumulation of cadmium, copper, manganese and zinc by oleaginous microalgae Chlorella minutissima UTEX2341. Bioresour. Technol. 2015, 175, 537–544. [Google Scholar] [CrossRef]
- Kan, G.; Wang, X.; Jiang, J.; Zhang, C.; Chi, M.; Ju, M.; Shi, C. Copper stress response in yeast Rhodotorula mucilaginosaAN5 isolated from sea ice. Antarctic. Microbiol. Open 2018, 8, e657. [Google Scholar] [CrossRef]
- Gong, G.; Liu, L.; Zhang, X.; Tan, T. Multi-omics metabolism analysis on irradiation-induced oxidative stress to Rhodotorula glutinis. Appl. Microbiol. Biotechnol. 2019, 103, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Maldonade, I.; Rodriguez-Amaya, D.; Scamparini, A. Carotenoids of yeasts isolated from the Brazilian ecosystem. Food Chem. 2008, 107, 145–150. [Google Scholar] [CrossRef]
- Kot, A.M.; Błażejak, S.; Kurcz, A.; Bryś, J.; Gientka, I.; Bzducha-Wróbel, A.; Maliszewska, M.; Reczek, L. Effect of initial pH of medium with potato wastewater and glycerol on protein, lipid and carotenoid biosynthesis by Rhodotorula glutinis. Electron. J. Biotechnol. 2017, 27, 25–31. [Google Scholar] [CrossRef]
- Saenge, C.; Cheirslip, B.; Suksaroge, T.T.; Bourtoom, T. Efficient concomitant production of lipids and carotenoids by oleaginous red yeast Rhodotorula glutinis cultured in palm oil mill effluent and application of lipids for biodiesel production. Biotechnol. Bioprocess. Eng. 2011, 16, 23–33. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C/N | N-Sources | g/L | C/S | C/S − C/N |
|---|---|---|---|---|
| 146 | * PEP | 1.5 | 240 | 94 |
| * YE | 1.7 | 240 | 94 | |
| (NH4)2SO4 | 0.9 | 133 | −13 | |
| 94 | PEP | 2.4 | 240 | 146 |
| YE | 2.7 | 240 | 146 | |
| (NH4)2SO4 | 1.4 | 116 | 22 | |
| 70 | PEP | 3.2 | 240 | 170 |
| YE | 3.6 | 240 | 170 | |
| (NH4)2SO4 | 1.9 | 89 | 19 |
| C/N | MgSO4 (g/L) | C/S | C/S − C/N |
|---|---|---|---|
| 146 | 0.5 | 245 | 108 |
| 1 | 166 | 20 | |
| 1.5 | 123 | −23 | |
| 2 | 98 | −48 |
| C/N | N | DCW (g/L) | TL (g/L) | TP (mg/L) | DCW-Y (%) | TL-Y (%) | TP-Y | L-RP (%) | Car-RP | Car-SR | L-SR (g/L·d) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 d | 6 d | 3 d | 6 d | 3 d | 6 d | 3 d | 6 d | 3 d | 6 d | 3 d | 6 d | 3 d | 6 d | 3 d | 6 d | 3 d | 6 d | 3 d | 6 d | ||
| 146 | PEP | 6.23 ± 0.25 | 9.3 ± 0.1 | 2.49 ± 0.52 | 5.04 ± 0.19 | 0.31 ± 0.16 | 0.36 ± 0.00 | 46.2 | 23.6 | 18.5 | 19.6 | 23.2 | 1.8 | 40.1 | 54.2 | 50.2 | 38.8 | 104 | 16.2 | 0.83 | 0.85 |
| YE | 6.62 ± 0.56 | 10.43 ± 0.49 | 2.47 ± 0.15 | 5.2 ± 0.18 | 0.44 ± 0.07 | 0.64 ± 0.03 | 46.7 | 24.9 | 17.5 | 17.9 | 31.3 | 14.5 | 37.4 | 50.0 | 67.0 | 61.6 | 148 | 66.3 | 0.82 | 0.92 | |
| AMM | 6.22 ± 0.17 | 10.17 ± 0.47 | 2.07 ± 0.84 | 4.81 ± 0.405 | 0.53 ± 0.04 | 0.79 ± 0.03 | 44.3 | 23.0 | 14.7 | 16 | 37.4 | 15.3 | 33.3 | 47.4 | 84.6 | 77.5 | 175 | 87.5 | 0.69 | 0.91 | |
| 96 | PEP | 7.77 ± 0.34 | 9.63 ± 0.25 | 2.50 ± 0.35 | 4.96 ± 0.37 | 0.48 ± 0.04 | 0.50 ± 0.03 | 47.5 | 21.3 | 15.3 | 28.0 | 27.8 | 2.0 | 32.2 | 51.5 | 58.5 | 52.4 | 152 | 16.7 | 0.83 | 0.82 |
| YE | 7.98 ± 0.23 | 11.68 ± 0.58 | 2.50 ± 0.18 | 5.4 ± 0.61 | 0.56 ± 0.03 | 0.79 ± 0.07 | 47.8 | 24.0 | 14.7 | 19.3 | 34.2 | 14.5 | 40.0 | 46.6 | 70.7 | 67.5 | 188 | 74.5 | 0.82 | 0.99 | |
| AMM | 6.90 ± 0.21 | 10.78 ± 0.11 | 1.90 ± 0.43 | 4.31 ± 0.57 | 0.56 ± 0.08 | 0.85 ± 0.10 | 46 | 20.5 | 12.6 | 12.8 | 37.0 | 15.4 | 27.5 | 40..0 | 80.5 | 78.6 | 185 | 96.9 | 0.63 | 0.8 | |
| 70 | PEP | 8.02 ± 0.18 | 10.14 ± 0.31 | 2.33 ± 0.51 | 4.75 ± 0.37 | 0.48 ± 0.06 | 0.53 ± 0.12 | 48.5 | 19.5 | 14.1 | 22.3 | 29.1 | 1.7 | 29.1 | 46.9 | 60.0 | 52.3 | 160 | 16.2 | 0.78 | 0.81 |
| YE | 8.32 ± 0.14 | 12.2 ± 0.2 | 2.20 ± 0.17 | 4.98 ± 0.14 | 0.57 ± 0.04 | 0.85 ± 0.25 | 50.0 | 24.7 | 13.3 | 17.5 | 33.7 | 17.9 | 26.7 | 40.8 | 68.4 | 69.7 | 190 | 94.1 | 0.74 | 0.92 | |
| AMM | 7.23 ± 0.47 | 11.09 ± 0.38 | 1.62 ± 0.3 | 3.87 ± 0.65 | 0.58 ± 0.02 | 0.88 ± 0.02 | 45.1 | 19.2 | 10.1 | 11.2 | 35.8 | 15.2 | 22.5 | 34.9 | 79.5 | 79.4 | 192 | 102 | 0.54 | 0.75 | |
| T | C/S | DCW (g/L) | DCW-Y (%) | TL (g/L) | TL-Y (%) | TL-RP (%) | TL-SR (g/L·d) | TP (mg/L) | TP-Y | Car-RP | Car-SR (µg/L·d) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 d | 254 | 6.49 ± 0.38 a | 48.7 | 2.5 ± 0.1 | 18.64 | 38.3 | 0.83 | 0.457 ± 0.05 | 34.4 | 70.7 | 152.9 |
| 166 | 6.79 ± 0.4 | 48.1 | 2.4 ± 0.5 | 17.28 | 35.4 | 0.81 | 0.565 ± 0.1 | 40.3 | 83.7 | 188.5 | |
| 123 | 7.26 ± 0.4 a | 48.9 | 2.4 ± 0.32 | 16.34 | 33.4 | 0.81 | 0.624 ± 0.01 d | 42.03 | 85.9 | 208.1 | |
| 98 | 7.11 ± 0.16 | 41.8 | 2.2 ± 0.22 | 13.2 | 31.6 | 0.75 | 0.612 ± 0.05 d | 35.9 | 86 | 203.86 | |
| 6 d | 254 | 9.59 ± 0.25 b | 22.4 | 5.1 ± 0.4 c | 19.02 | 53.4 | 0.88 | 0.528 ± 0.3 | 5.026 | 55.1 | 23.2 |
| 166 | 10.45 ± 0.39 | 22.8 | 5.1 ± 0.29 c | 16.55 | 48.85 | 0.89 | 0.807 ± 0.08 e | 14.9 | 77.2 | 80.6 | |
| 123 | 10.74+0.06 b | 25.6 | 4.8 ± 0.18 | 17.7 | 44.98 | 0.8 | 0.865 ± 0.01 e | 17.7 | 80.5 | 80.4 | |
| 98 | 10.91 ± 0.56 | 22.9 | 4.5 ± 0.18 | 13.84 | 41.68 | 0.77 | 0.875 ± 0.01 e | 15.9 | 80.2 | 87.89 |
| YE-146 | YE-94 | YE-70 | PEP-146 | PEP-94 | PEP-70 | Amm-146 | Amm-94 | Amm-70 | C/S-254 | C/S-166 | C/S-123 | C/S-98 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Torh | 3.3 ± 3.3 | 6.7 ± 2.8 | 5.3 ± 4.9 | 6.2 ± 2.1 | 12.05 ± 3.96 | 3.6 ± 1.9 | 10.5 ± 0.49 | 7.1 ± 6.8 | 0.78 ± 5 | 0 | 0 | 0 | 0 |
| Toru | 52.5 ± 6.1 | 52.4 ± 3.0 | 37.5 ± 10 | 57.3 ± 7.4 | 55.4 ± 9.7 | 50.5 ± 3.5 | 62 ± 8.5 | 61.1 ± 5.4 | 57.4 ± 6.9 | 50.1 ± 0.71 | 53.4 ± 5.9 | 54.7 ± 3.8 | 54.8 ± 1 |
| γ-car | 44.2 ± 4.2 | 38.6 ± 2.5 | 52.3 ± 8.8 | 33.6 ± 3.6 | 28.3 ± 3.6 | 44.4 ± 5.6 | 11.1 ± 4 | 24 ± 10 | 32.1 ± 3.1 | 21.4 ± 2.3 | 33.6 ± 4.5 | 40.3 ± 4.3 | 41.5 ± 3.2 |
| β- car | 0 | 0 | 0 | 0 | 0 | 0 | 5.4 ± 4.8 | 7.6 ± 2.9 | 8.9 ± 4.1 | 12.2 ± 4.2 | 0 | 0 | 0 |
| Stress Condition | Yeast strains | TP (mg/L) | Torulene (mg/L) | Car-RP (µgP/gDCW) | C/N | NS | Carbon Source | C/S | Ref |
|---|---|---|---|---|---|---|---|---|---|
| None | R.glutinis | 0.881 | 0.261 | 13 | 3SC | YE, ME, &PEP | Glucose & maltose | NC | [57] |
| R. graminis | 0.594 | 0.186 | 70 | ||||||
| R. mucilaginosa-137 | 0.59 | 0.352 | 69 | ||||||
| R. mucilaginosa-108 | 0.562 | 0.307 | 68 | ||||||
| R. mucilaginosa-135 | 0.545 | 0.356 | 76 | ||||||
| R. minuta | 0.168 | – | 33 | ||||||
| Sporobolomyces roseus | 0.237 | 0.071 | 72 | ||||||
| None | R. glutinis DBVPG6081 | 0.631 | 0.136 | 112.2 | 57 | YE | Glucose | 319SC | [11] |
| None | R. glutinis DBVPG7021 | 1.5 | 0.213 | 162.7 | |||||
| None | R. sp. nov CRUB1032 | 1.2 | 0.95 | 138.6 | |||||
| None | Sporidiobolus CRUB1044 | 1.99 | 1.9 | 184 | |||||
| None | Rhod. toruloides DBVPG6739 | 1.311 | 0.68 | 122.6 | |||||
| None | R. glutinis NCIM3353 | 2.2 | 1.16 | 120 | 10 | YE | Glucose | – | |
| None | R. glutinis var.glutinis | 2.4 | 1.4 | 333 | 10 | AMM | Glucose | 199SC | [19,50] |
| None | 2.3 | 2 | 273 | 10 | PEP | ||||
| 0.1 g/L MnSO4 | 1.64 | 1.3 | 304 | 10 | AMM | ||||
| 0.1 g/L ZnSO4 | 2.81 | 1.6 | 638 | 10 | AMM | ||||
| None | R. glutinis JMT 21978 | 18 | 3.5sc | 1600 | 50 | YE | Glucose | – | |
| None | R. glutinis ATCC 15125) | 1.2 | – | 120 | 120 | YE:Amm | Glucose | 919SC | [10] |
| None | Sporidiobolus pararoseus NGR | 1.636 | 0.251 | – | 3.6 | YE:PEP | Glucose | – | [12] |
| 0.75 M NaCl | 3.952 | 2.1 | – | 3.6 | – | ||||
| Unoptimized | R. glutinis AS 2.703 | 0.789 | 0.498 | 77.5 | 146 | Amm | Glucose | 133 | This study |
| Optimized media | 0.95 | 0.589 | 89.05 | 120 | |||||
| Optimized media + 0.7 mM Al2(SO4)3 | 2.2 | 2.16 | 212.9 | 120 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elfeky, N.; Elmahmoudy, M.; Zhang, Y.; Guo, J.; Bao, Y. Lipid and Carotenoid Production by Rhodotorula glutinis with a Combined Cultivation Mode of Nitrogen, Sulfur, and Aluminium Stress. Appl. Sci. 2019, 9, 2444. https://doi.org/10.3390/app9122444
Elfeky N, Elmahmoudy M, Zhang Y, Guo J, Bao Y. Lipid and Carotenoid Production by Rhodotorula glutinis with a Combined Cultivation Mode of Nitrogen, Sulfur, and Aluminium Stress. Applied Sciences. 2019; 9(12):2444. https://doi.org/10.3390/app9122444
Chicago/Turabian StyleElfeky, Nora, Mostafa Elmahmoudy, Yue Zhang, JianLi Guo, and Yongming Bao. 2019. "Lipid and Carotenoid Production by Rhodotorula glutinis with a Combined Cultivation Mode of Nitrogen, Sulfur, and Aluminium Stress" Applied Sciences 9, no. 12: 2444. https://doi.org/10.3390/app9122444
APA StyleElfeky, N., Elmahmoudy, M., Zhang, Y., Guo, J., & Bao, Y. (2019). Lipid and Carotenoid Production by Rhodotorula glutinis with a Combined Cultivation Mode of Nitrogen, Sulfur, and Aluminium Stress. Applied Sciences, 9(12), 2444. https://doi.org/10.3390/app9122444