Evaluation of Some Rhodes Grass (Chloris gayana) Genotypes for Their Salt Tolerance, Biomass Yield and Nutrient Composition



Abstract
1. Introduction
2. Materials and Methods
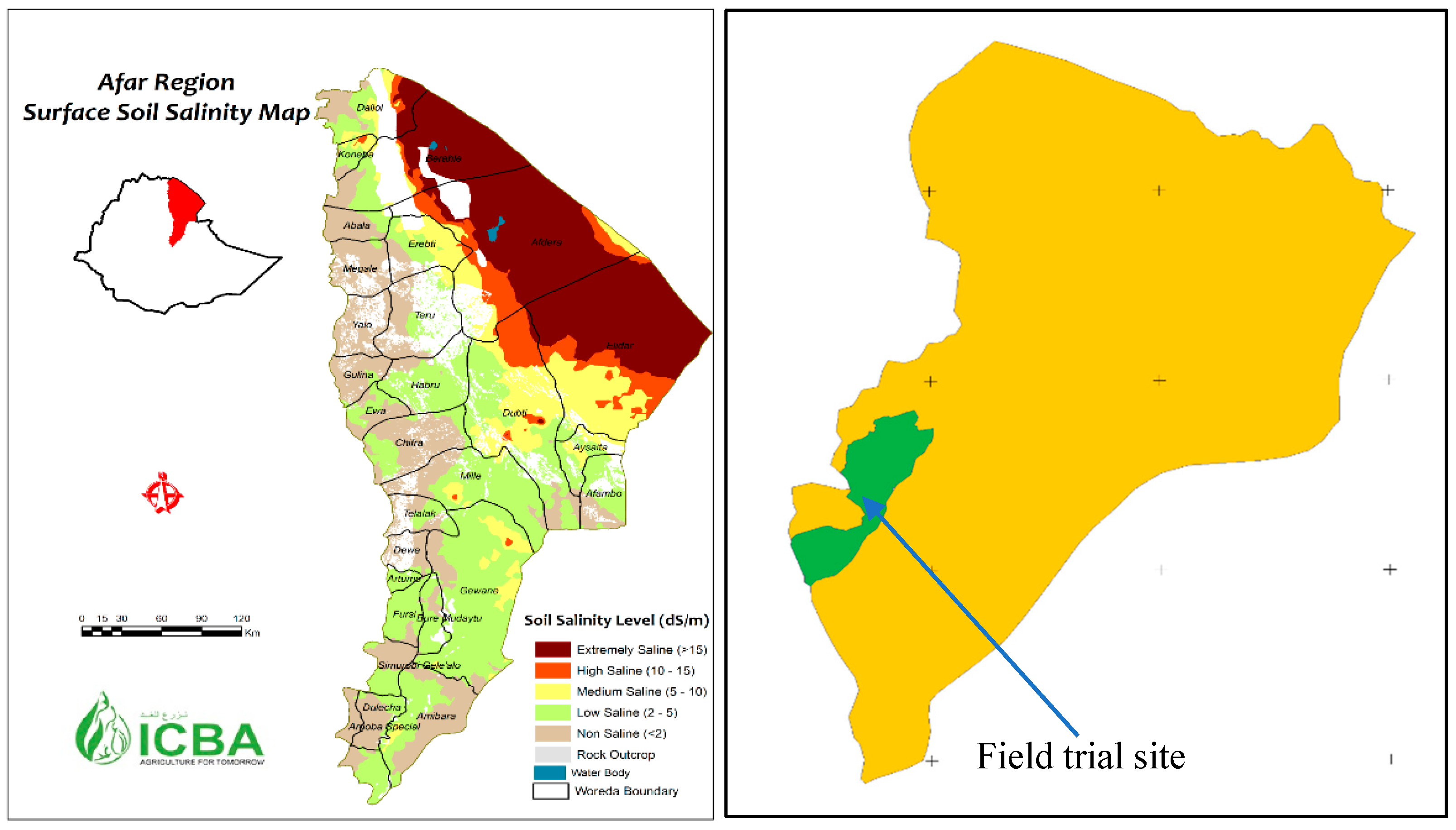
2.1. Study Area
2.2. Experimental Design and Salinity Treatments
2.3. Data Collection and Analysis
3. Results and Discussion
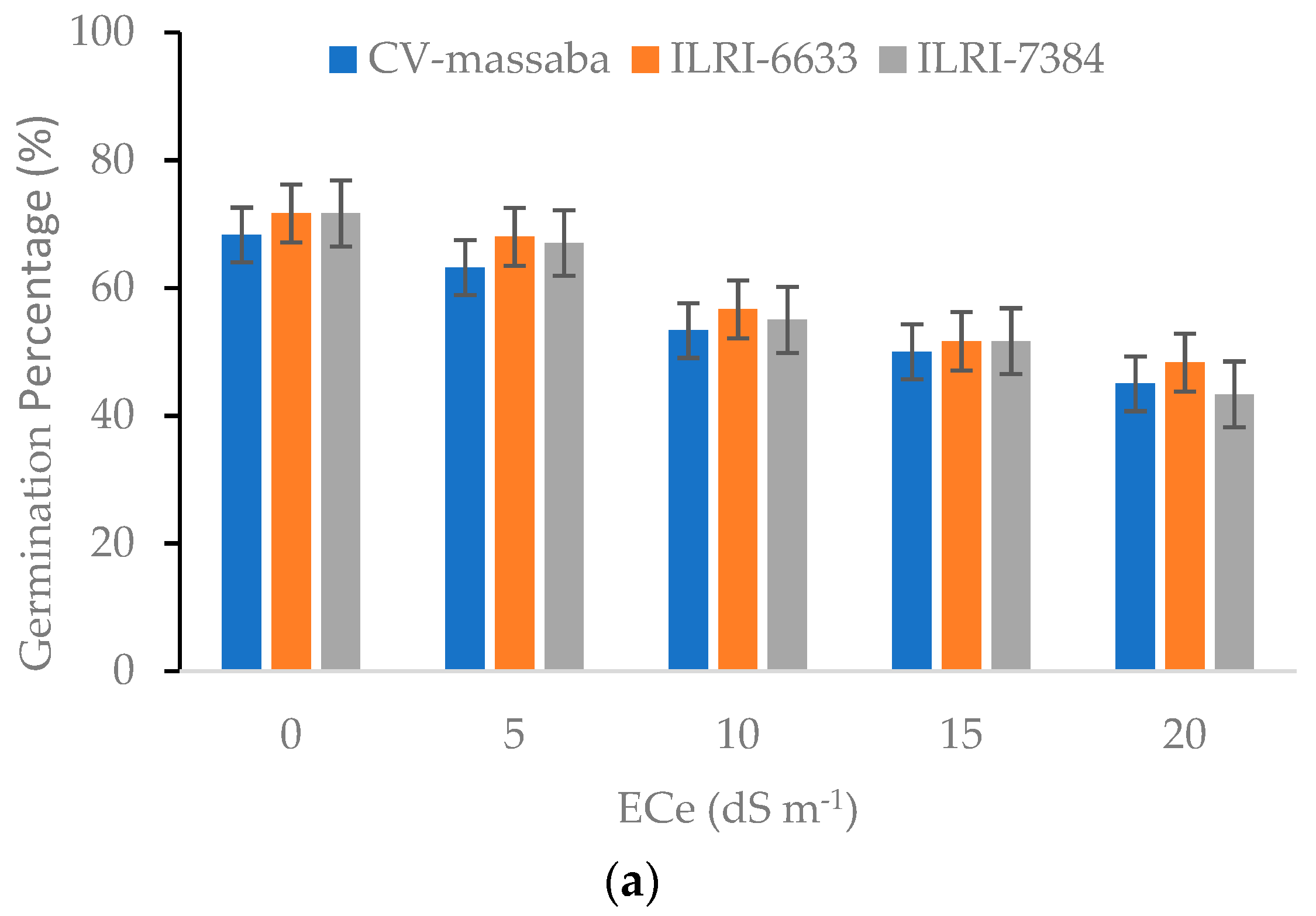
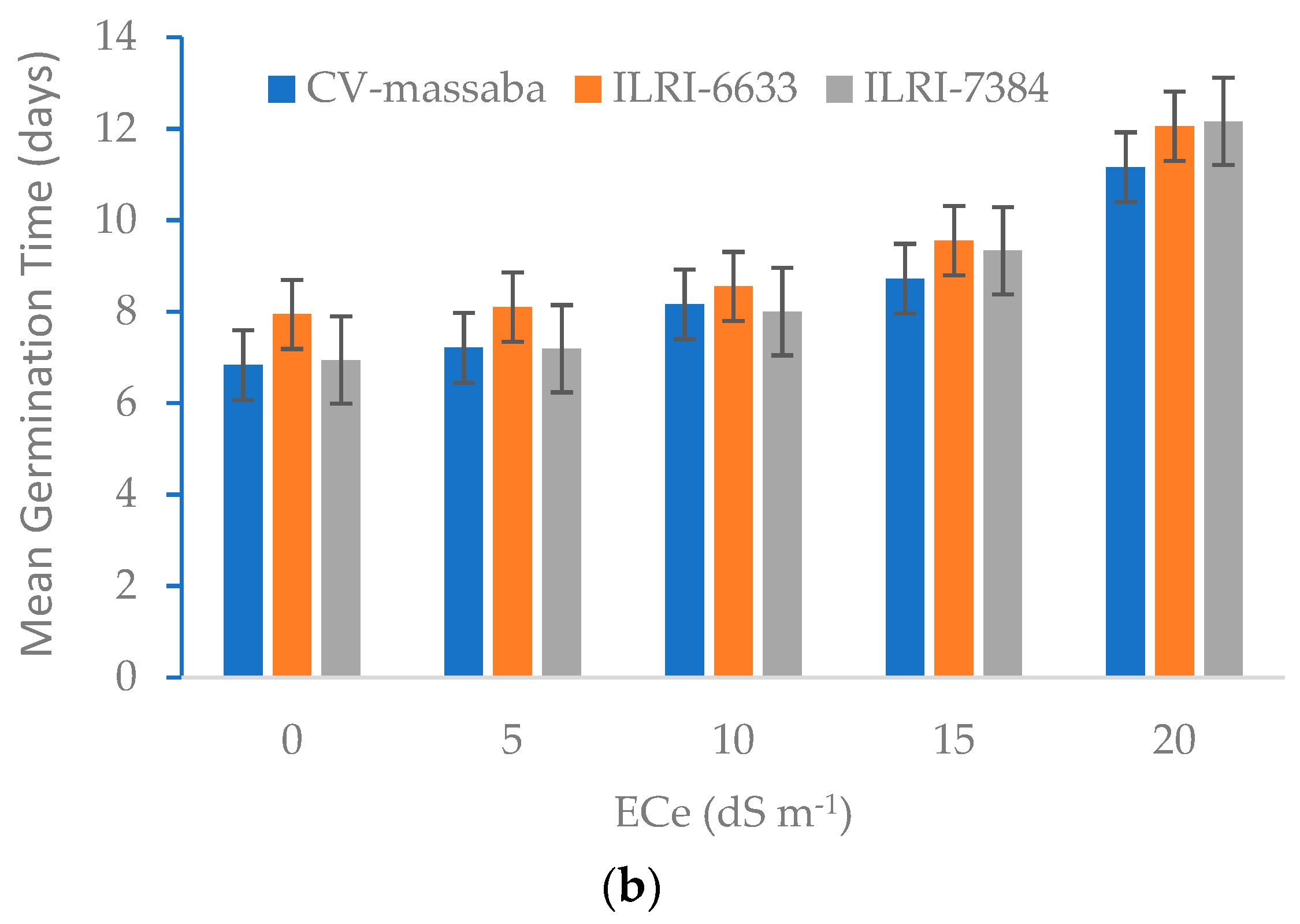
3.1. Effect of Soil Salinity on Germination Percentage (GP) and Mean Germination Time (MGT)
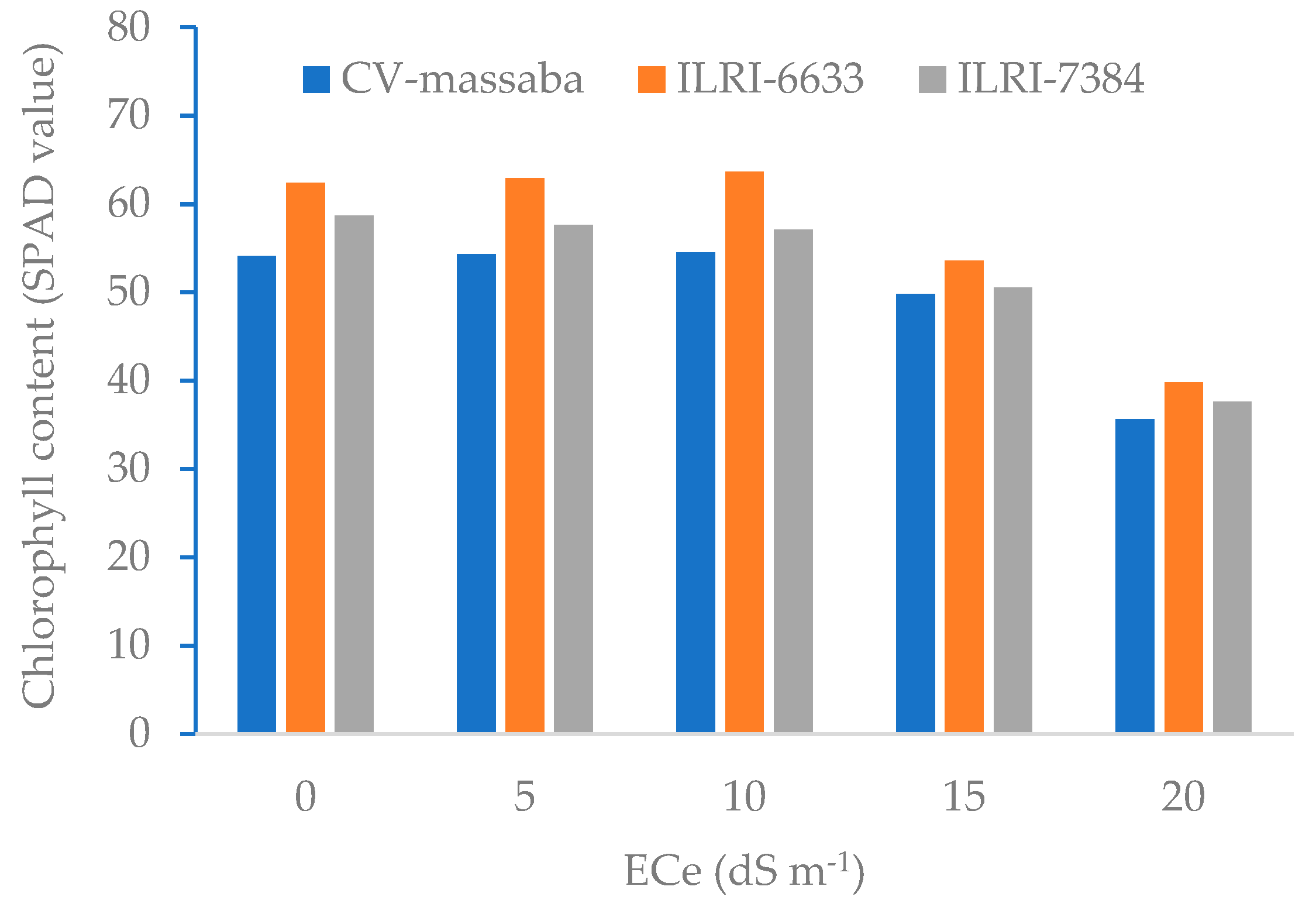
3.2. Plant Height, Number of Tillers and Chlorophyll SPAD Value
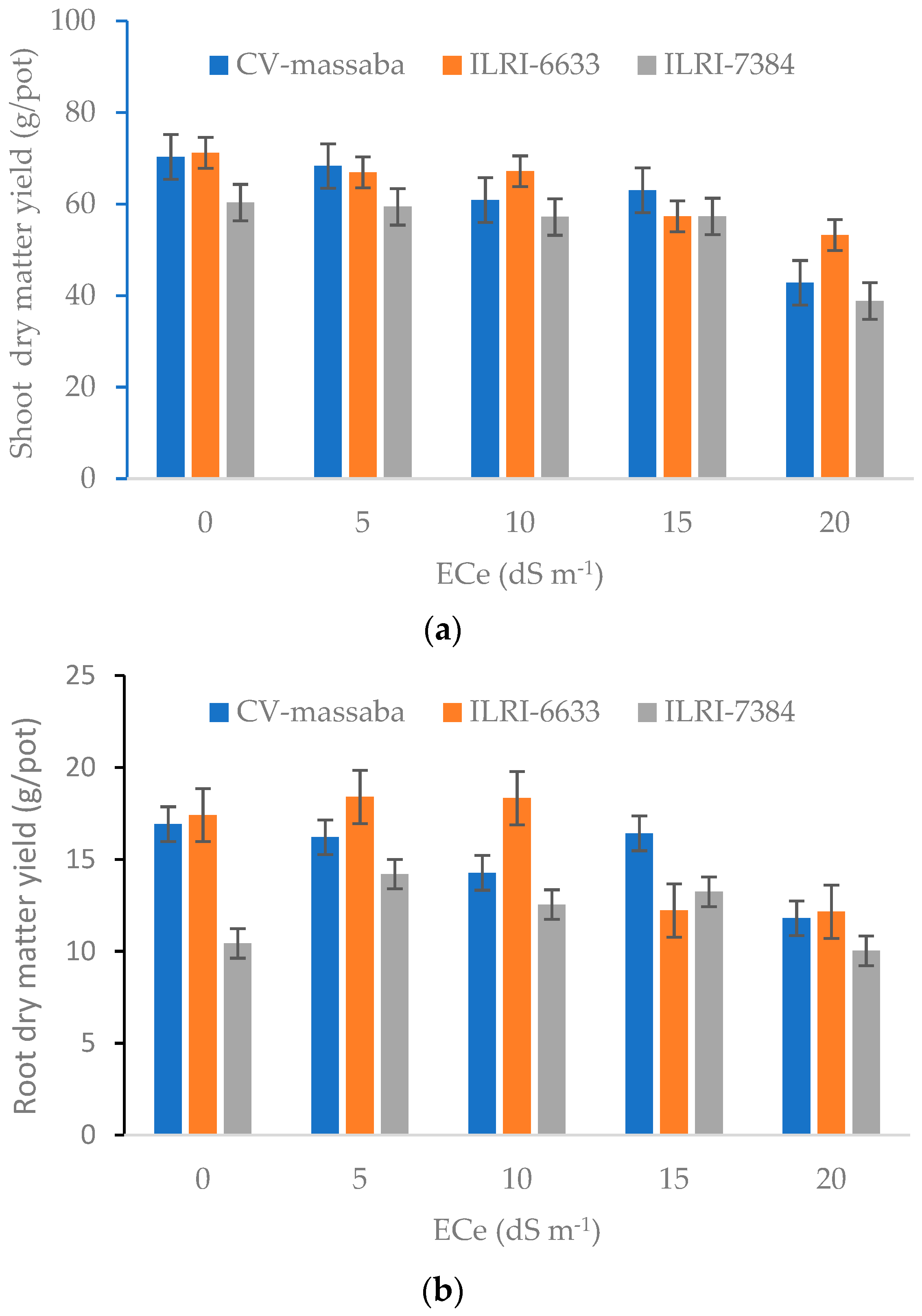
3.3. Root and Shoot Dry Matter
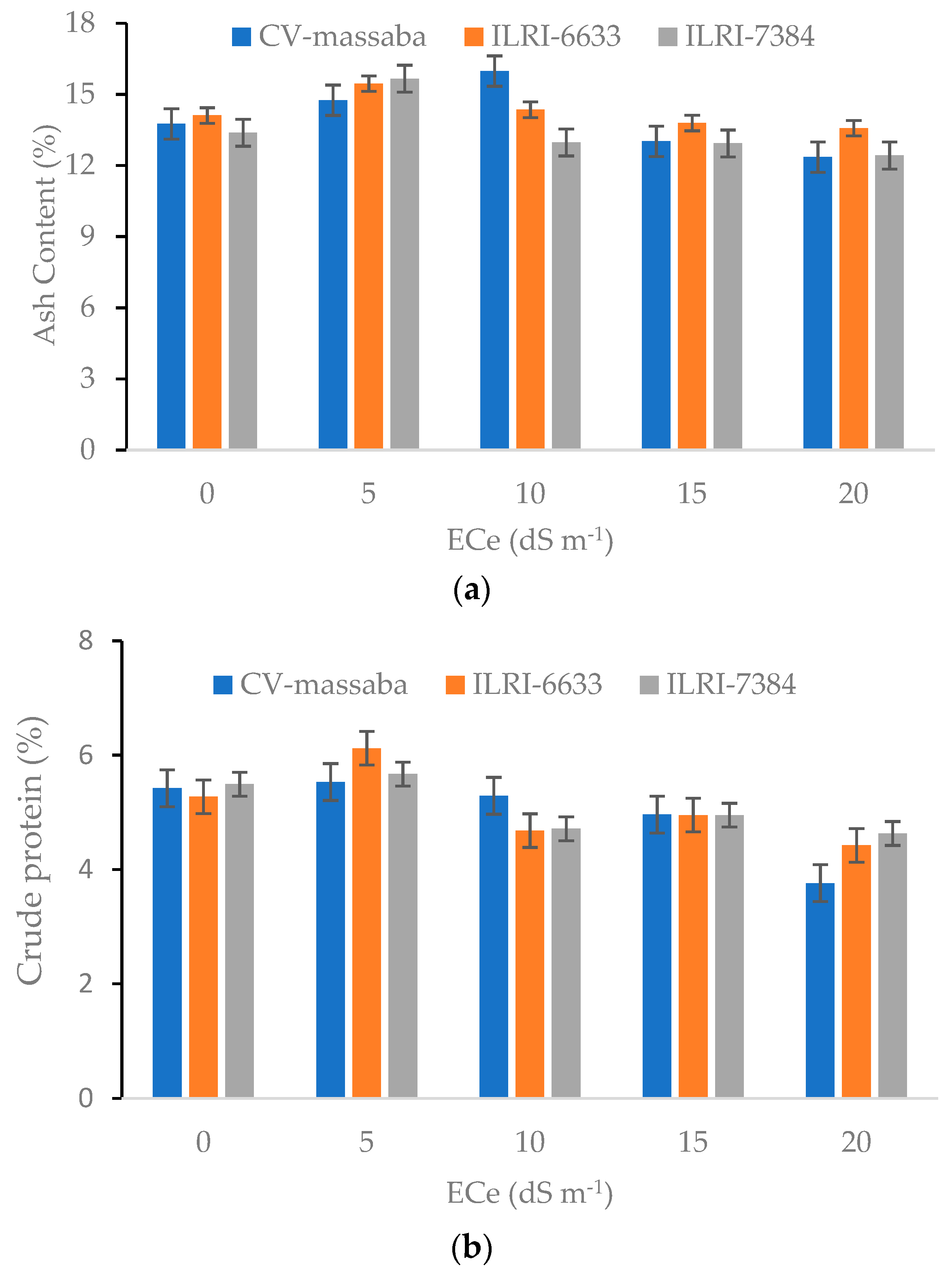
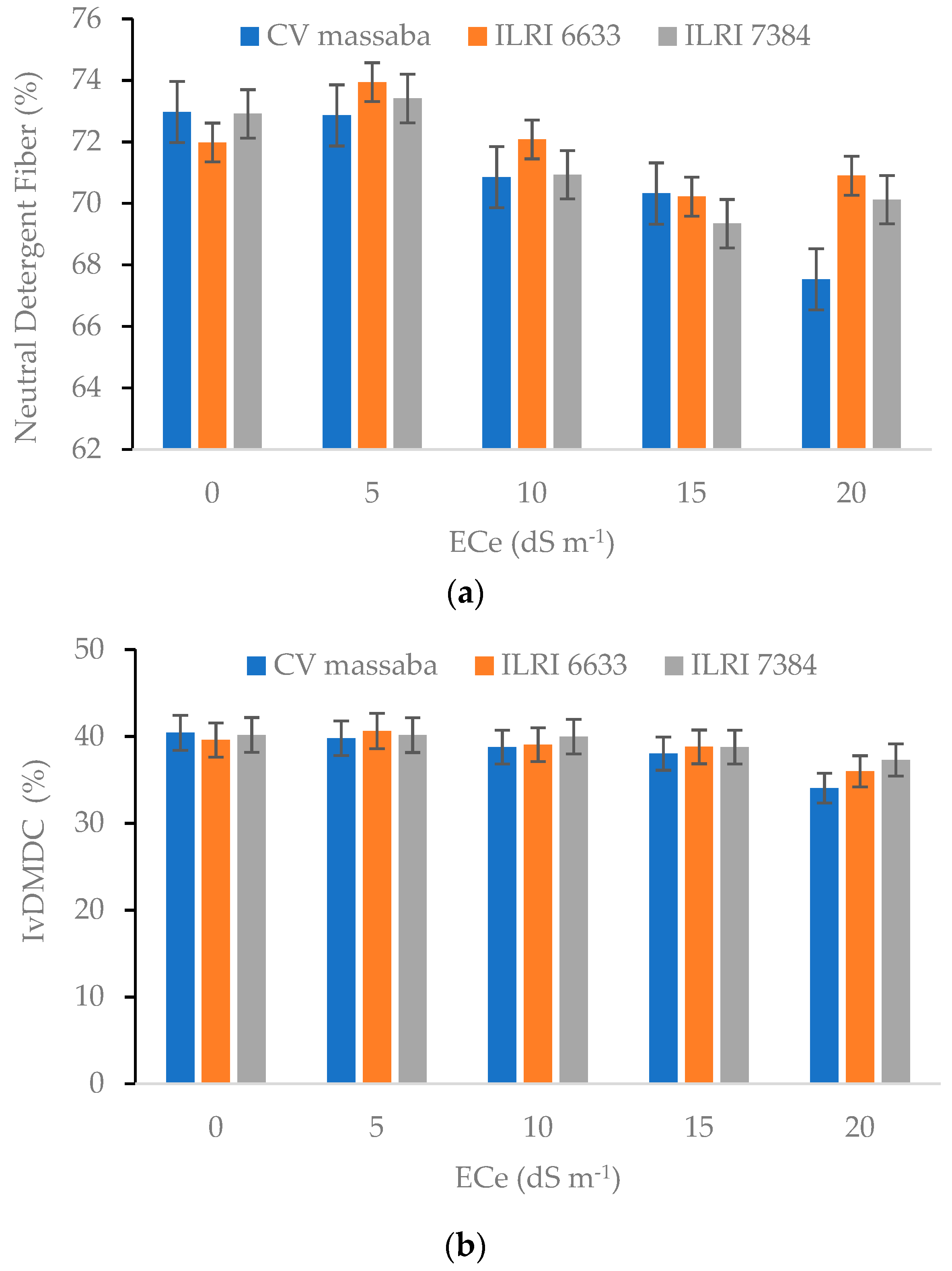
3.4. Nutrient Composition
4. Conclusions and Recommendations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Bhowmik, P.C.; Hossain, M.A.; Rahman, M.M.; Prasad, M.N.V.; Ozturk, M.; Fujita, M. Potential use of halophytes to remediate saline soils. BioMed Res. Int. 2014. [Google Scholar] [CrossRef]
- Ventura, Y.; Sagi, M. Halophyte crop cultivation: The case for Salicornia and sarcocornia. Environ. Exp. Bot. 2013, 92, 144–153. [Google Scholar] [CrossRef]
- Wicke, B.; Smeets, E.; Dornburg, V.; Vashev, B.; Gaiser, T.; Turkenburg, W.; Faaij, A. The global technical and economic potential of bioenergy from salt-affected soils. Energy Environ. Sci. 2011, 8, 2669–2681. [Google Scholar] [CrossRef]
- Qadir, M.; Quillerou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.J. Economics of salt-induced land degradation and restoration. Nat. Res. Forum 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Munns, R. Plant adaptations to salt and water stress: Differences and commonalities. Adv. Bot. Res. 2011, 57, 1–32. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Kidane, G.; Abebe, F.; Heluf, G.; Fentaw, A.; Wondimadegne, C.; Hibstu, A.; Asegid, A.; Messele, F.; Mohammed, B. Report of the National Task Force on Assessment of Salt Affected Soils and Recommendations on Management Options for Sustainable Utilization; Ethiopia, A.A., Ed.; Minister of Agriculture and Rural Development: Addis Ababa, Ethiopia, 2006; 120p.
- Frew, A.; Tena, A.; Fentaw, A. Appraisal and mapping of soil salinity problems in Amibara area of Middle Awash Basin Ethiopia. Int. J. Innov. Sci. Res. 2015, 13, 298–314. [Google Scholar]
- UNICEF (United Nations Children’s Fund). The State of the World’s Children 2016: A Fair Chance for Every Child; UNICEF: New York, NY, USA, 2016; Available online: www.unicef.org/publications/files/UNICEF_SOWC_2016.pdf (accessed on 22 July 2016).
- Jacobsen, S.E.; Mujica, A.; Jensen, C.R. The resistance of quinoa (Chenopodium quinoa Wild.) to adverse abiotic factors. Food Rev. Int. 2003, 19, 99–109. [Google Scholar] [CrossRef]
- Rosa, M.; Hilal, M.; González, J.A.; Prado, F.E. Low-temperature effect on enzyme activities involved in sucrose-starch partitioning in salt-stressed and salt-acclimated cotyledons of quinoa (Chenopodium quinoa Willd.) seedlings. Plant Physiol. Biochem. 2009, 47, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Bazile, D.; Baudron, F. The dynamics of the global expansion of quinoa growing in view of its high biodiversity. In State of the Art Report on Quinoa Around the World in 2013; Bazile, D., Bertero, H.D., Nieto, C., Eds.; FAO: Rome, Italy, 2015; pp. 42–55. [Google Scholar]
- Akil, H. Biological Improvement of Lime-Saline-Sodic Soil in Harran Plain. Master’s Thesis, Harran Universdity Institute of Science, Sanliurfa, Turkey, 2008. [Google Scholar]
- Temel, S.; Keskin, B.; Simsek, U.; Ibrahim, H.Y. Performance of Some Forage Grass Species in Halomorphic Soil. Turk. J. Field Crops 2015, 20, 131–141. [Google Scholar] [CrossRef]
- Koc, A.; Tan, M. Role of Forage Crops Against Salinity Problem Arising in the GAP region. In Proceedings of the GAP I. Agricultural Congress, Sanliurfa, Turkey, 26–28 May 1999. [Google Scholar]
- Al-Dakheel, A.J.; Hussain, M.I.; Abdul Rahman, A.Q. Impact of irrigation water salinity on agronomical and quality attributes of Cenchrus ciliaris L. accessions. Agric. Water Manag. 2015, 159, 148–154. [Google Scholar] [CrossRef]
- Hassen, A.; Souguir, M.; Hannachi, C. Effect of Salt Stress (NaCl) on Germination and Early Seedling Parameters of Three Pepper Genotypes (Capsicum annuum L.). J. Stress Physiol. Biochem. 2014, 10, 15–25. [Google Scholar]
- Hailay, T.; Tadele, G.S.; Tekalign, M. Assessment of salinity/sodicity problems in Abaya State Farm, Southern Rift Valley of Ethiopia. Ethiop. J. Nat. Res. 2000, 2, 151–163. [Google Scholar]
- Ashraf, M.; Foolad, M.R. Pre-sowing seed treatment-a shotgun approach to improve germination growth and crop yield under saline and none-saline conditions. Adv. Agron. 2005, 88, 223–271. [Google Scholar]
- Ellis, R.A.; Roberts, E.H. The quantification of ageing and survival in orthodox seeds. Seed Sci. Technol. 1981, 9, 373–409. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Kitcherside, M.A.; Glen, E.F.; Webster, A.J.F. FibreCap: An improved methodfor the rapid analysis of fibre in feeding stuffs. Anim. Feed Sci. Technol. 2000, 86, 125–132. [Google Scholar] [CrossRef]
- Gomez, A.; Gomez, H. Statistical Analysis for Agricultural Research; John Willy and Sons Inc.: Hoboken, NJ, USA, 1984; pp. 120–155. [Google Scholar]
- Horst, G.L.; Dunning, N.B. Germination and seedling growth of perennial ryegrasses in soluble salts. J. Am. Soc. Hort. Sci. 1989, 114, 338–342. [Google Scholar]
- Iqbal, N.M.Y.; Ashraf, F.; Javed, V.; Martinez, K. Nitrate reduction and nutrient accumulation in wheat (Triticum aestivum L.) grown in soil salinization with four different salts. J. Plant Nutr. 2006, 29, 409–421. [Google Scholar] [CrossRef]
- Azhar, M.F.; McNeilly, T. Variability for salt tolerance in Sorghumn bicolor (L.) Moench under hydroponic conditions. J. Agron. Crop Sci. 1987, 159, 269–277. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Vicente, O.; Boscaiu, M.; Naranjo, M.A.; Estrelles, E.; Belles, J.M.; Soriana, P. Responses to salt stress in the halophyte Plantago crassifolia (Plantaginaceae). J. Arid Environ. 2004, 58, 463–481. [Google Scholar] [CrossRef]
- Almodares, A.; Hadi, M.R.; Dosti, B. Effects of salt stress on germination percentage and seedling growth in sweet sorghum genotypes. J. Biol. Sci. 2007, 7, 1492–1495. [Google Scholar]
- Al-Dakheel, A.J.; Hussain, M.I. Genotypic Variation for Salinity Tolerance in Cenchrus ciliaris L. Front. Plant Sci. 2016, 7, 1090. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Shirazi, M.U.; Khan, M.A.; Mujtaba, S.M.; Islam, E.; Mumtaz, S.; Shereen, A.; Ansari, R.U.; Ashraf, M.Y. Role of proline, K+/Na+ ratio and chlorophyll content in salt tolerance of wheat (Triticum aestivum L.). Pak. J. Bot. 2009, 41, 633–638. [Google Scholar]
- Harinasut, P.; Srisunak, S.; Pitukchaisopol, S.; Charoensataporn, R. Mechanisms of adaptation to increasing salinity of mulberry: Proline content and ascorbate peroxidase activity in leaves of multiple shoots. Sci. Asia 2000, 26, 207–211. [Google Scholar] [CrossRef]
- McConnell, J.S.; Francis, P.B.; Stark, C.R.; Glover, R.E. Plant responses of ultra-narrow row cotton to nitrogen fertilization. J. Plant Nutr. 2008, 31, 1005–1017. [Google Scholar] [CrossRef]
- Kumari, P.; Pahuja, S.K.; Sheoran, R.S.; Satyawan, A.; Joshi, U.N. Effect of Varying Levels of Salinity on Growth, Yield and Quality of Forage Sorghum Genotypes. Forage Res. 2017, 43, 64–66. [Google Scholar]
- Robinson, P.H.; Grattan, S.R.; Getachew, G.; Grieve, C.M.; Poss, J.A.; Suarez, D.L.; Benes, S.E. Biomass accumulation and potential nutritive value of some forages irrigated with saline-sodic drainage water. Anim. Feed Sci. Technol. 2004, 111, 175–189. [Google Scholar] [CrossRef]
- Masters, D.G.; Benesand, S.E.; Norman, H.C. Biosaline Agriculture for Forage and Livestock Production. Agric. Ecosyst. Environ. 2007, 119, 234–248. [Google Scholar] [CrossRef]
- Qadir, M.; Khan, Z.H.; Majeed, A.; Yaqoob, S.; Khan, R.A.; Anjum, K. Effect of salinity on forage production of range grasses. Pak. J. Sci. 2008, 60, 59–63. [Google Scholar]
- Kandil, A.A.; Sharif, A.E.; Abido, W.A.E.; İbrahim, M.M. Effect of salinity on seed germination and seedling characters of some forage sorghum genotypes. Int. J. Agric. Sci. 2012, 4, 306–311. [Google Scholar]
- Suyama, H.; Benes, S.B.; Robinson, P.H.; Grattan, S.R.; Grieve, C.M.; Getachew, G. Forage yield and quality under irrigation with saline-sodic drainage water: Greenhouse evaluation. Agric. Water Manag. 2007, 88, 159–173. [Google Scholar] [CrossRef]
- Ashenafi, W.; Bobe, B. Studies on Soil Physical Properties of Salt Affected Soil in Amibara Area, Central Rift Valley of Ethiopia. Int. J. Agric. Sci. Nat. Resour. 2016, 3, 8–17. [Google Scholar]
- Sima, N.A.; Ahmad, S.T.; Pessarakli, M. Comparative Study of Different Salts (Sodium Chloride, Sodium Sulfate, Potassium Chloride, and Potassium Sulfate) on Growth of Forage Species. J. Plant Nutr. 2013, 36, 214–230. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Genotypes | Salinity Level (dS m−1) | LSD (p ≤ 0.05) | CV (%) | ||||
|---|---|---|---|---|---|---|---|---|
| 0 | 5 | 10 | 15 | 20 | ||||
| Plant height (cm) | CV-massaba | 101.3 | 100.3 | 97.0 | 76.3 | 63.8 | 7.3 | 9.0 |
| ILRI-6633 | 120.8 | 117.8 | 103.6 | 78.3 | 68.9 | |||
| ILRI-7384 | 93.0 | 92.5 | 81.4 | 60.3 | 52.0 | |||
| Total tillers (#) | CV-massaba | 6.9 | 6.8 | 6.3 | 5.2 | 5.5 | 1.1 | 17.6 |
| ILRI-6633 | 5.4 | 5.3 | 5.0 | 4.7 | 3.1 | |||
| ILRI-7384 | 9.4 | 9.3 | 8.4 | 8.0 | 5.9 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daba, A.W.; Qureshi, A.S.; Nisaren, B.N. Evaluation of Some Rhodes Grass (Chloris gayana) Genotypes for Their Salt Tolerance, Biomass Yield and Nutrient Composition. Appl. Sci. 2019, 9, 143. https://doi.org/10.3390/app9010143
Daba AW, Qureshi AS, Nisaren BN. Evaluation of Some Rhodes Grass (Chloris gayana) Genotypes for Their Salt Tolerance, Biomass Yield and Nutrient Composition. Applied Sciences. 2019; 9(1):143. https://doi.org/10.3390/app9010143
Chicago/Turabian StyleDaba, Ashenafi Worku, Asad Sarwar Qureshi, and Bethel Nekir Nisaren. 2019. "Evaluation of Some Rhodes Grass (Chloris gayana) Genotypes for Their Salt Tolerance, Biomass Yield and Nutrient Composition" Applied Sciences 9, no. 1: 143. https://doi.org/10.3390/app9010143
APA StyleDaba, A. W., Qureshi, A. S., & Nisaren, B. N. (2019). Evaluation of Some Rhodes Grass (Chloris gayana) Genotypes for Their Salt Tolerance, Biomass Yield and Nutrient Composition. Applied Sciences, 9(1), 143. https://doi.org/10.3390/app9010143