
A Simple Predator-Prey Population Model with Rich Dynamics

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Model Formulation
3. Preliminary Results
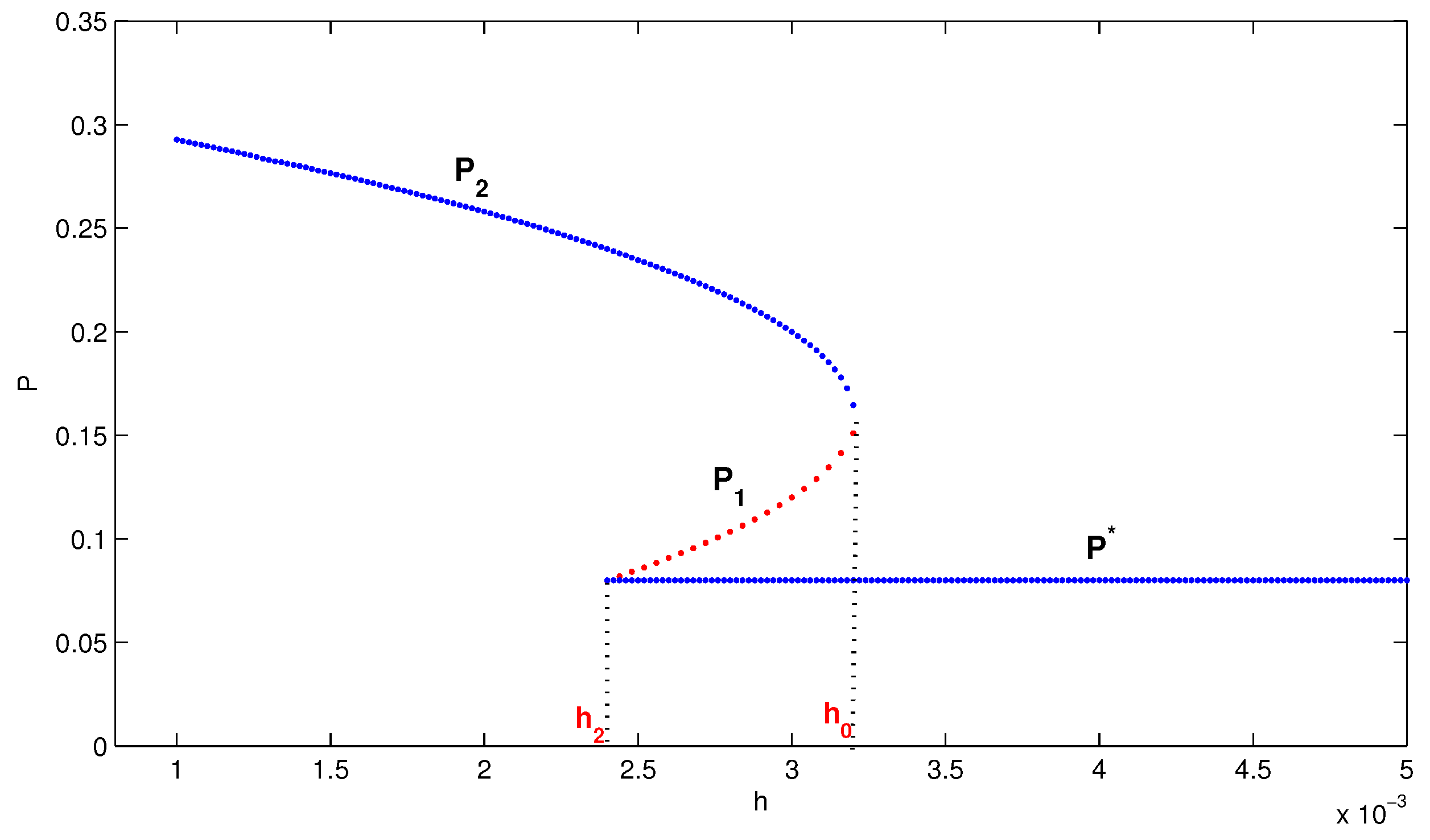
3.1. Existence of Equilibria
- (a)
- No positive equilibrium if
- (b)
- A unique positive equilibrium withif
- (c)
- Two positive equilibria , , whereandif and
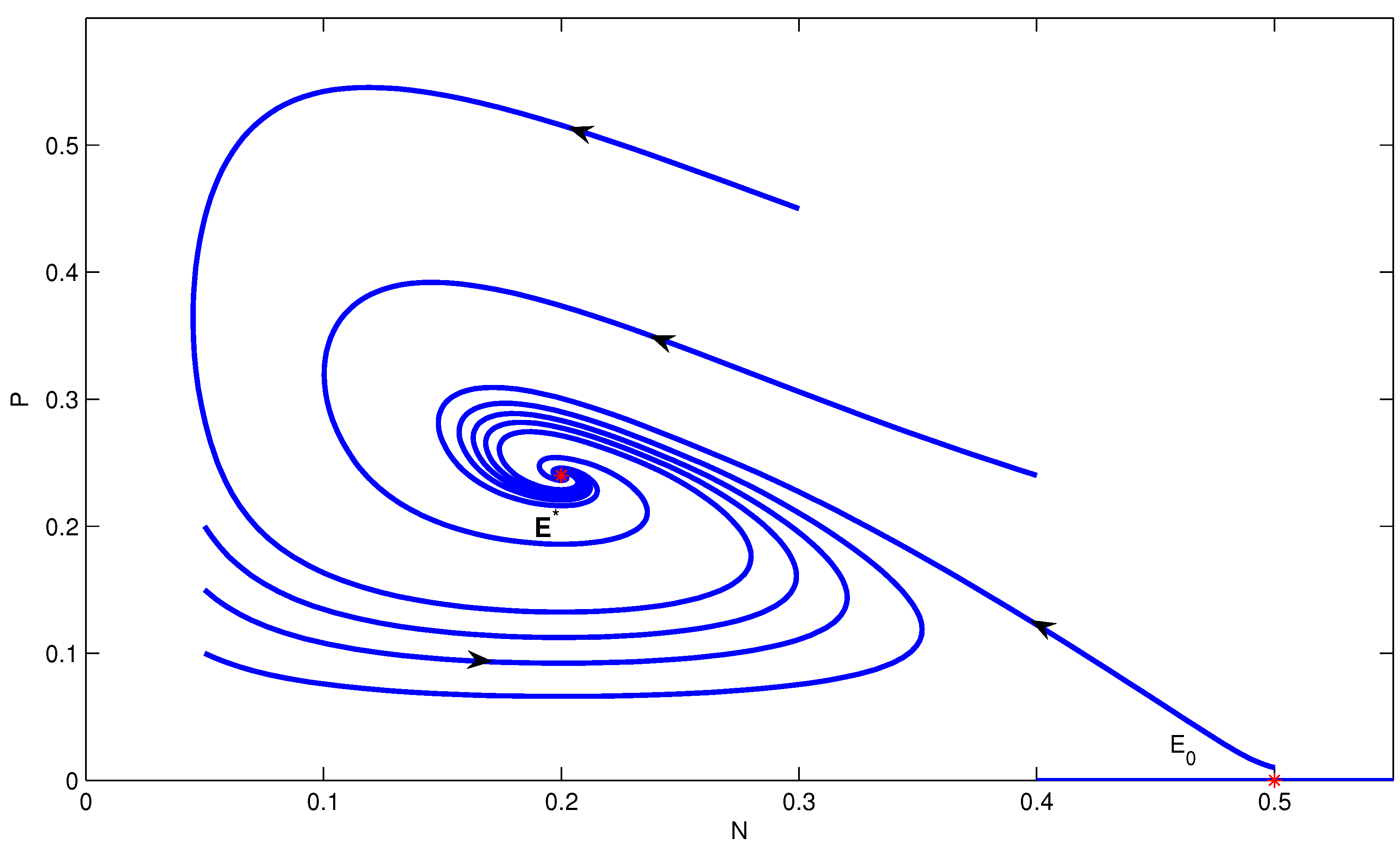
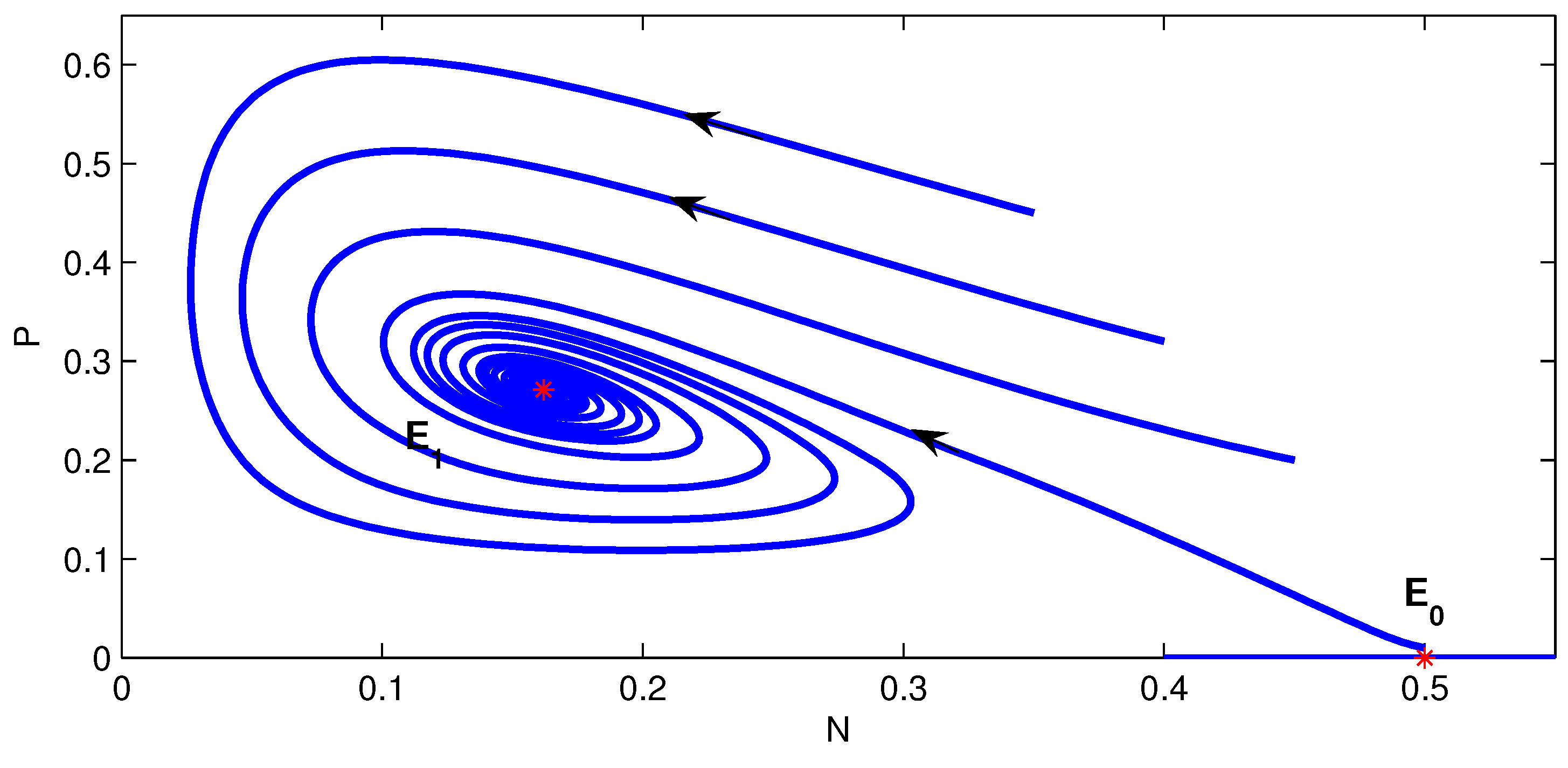
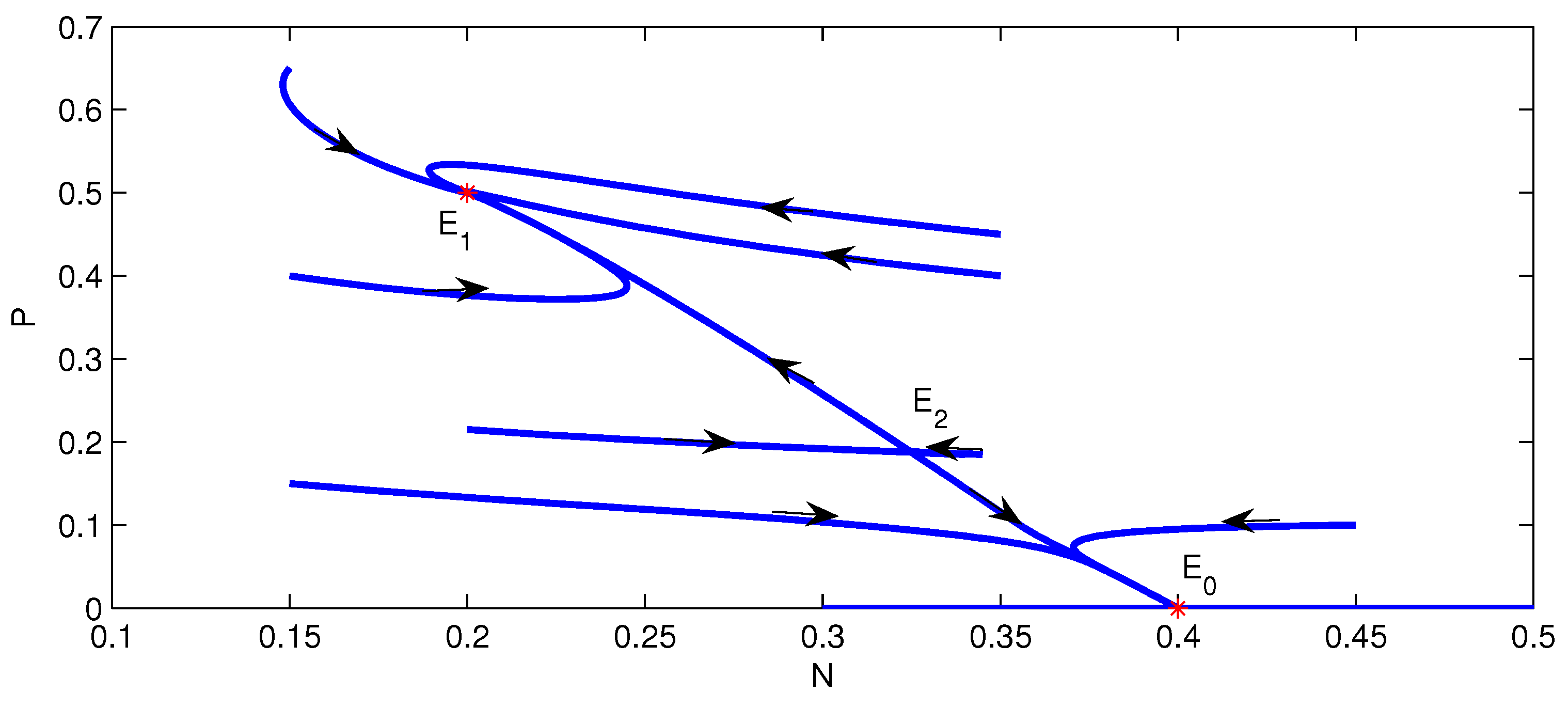
3.2. Stability of Equilibria
- (a)
- If , then .
- (b)
- If , and , then .
- (c)
- If , and , then .
- (a)
- ;
- (b)
- and ;
- (c)
- and and ,
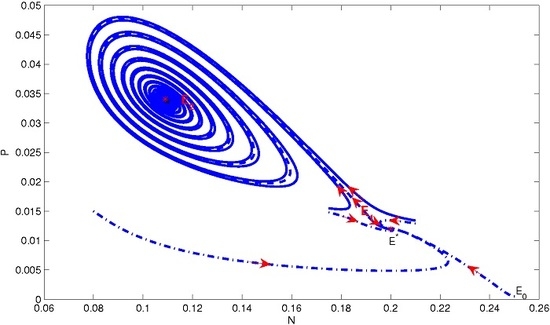
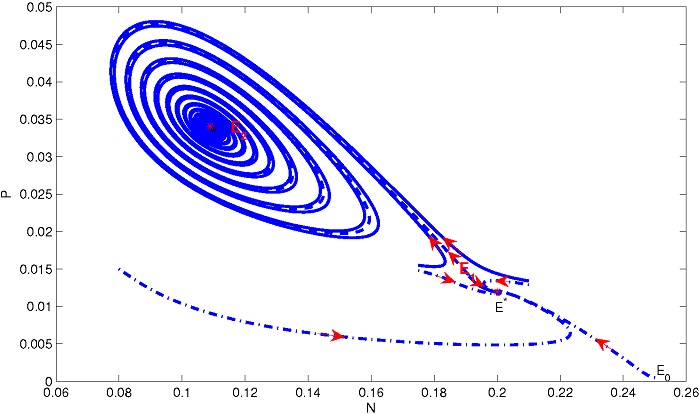
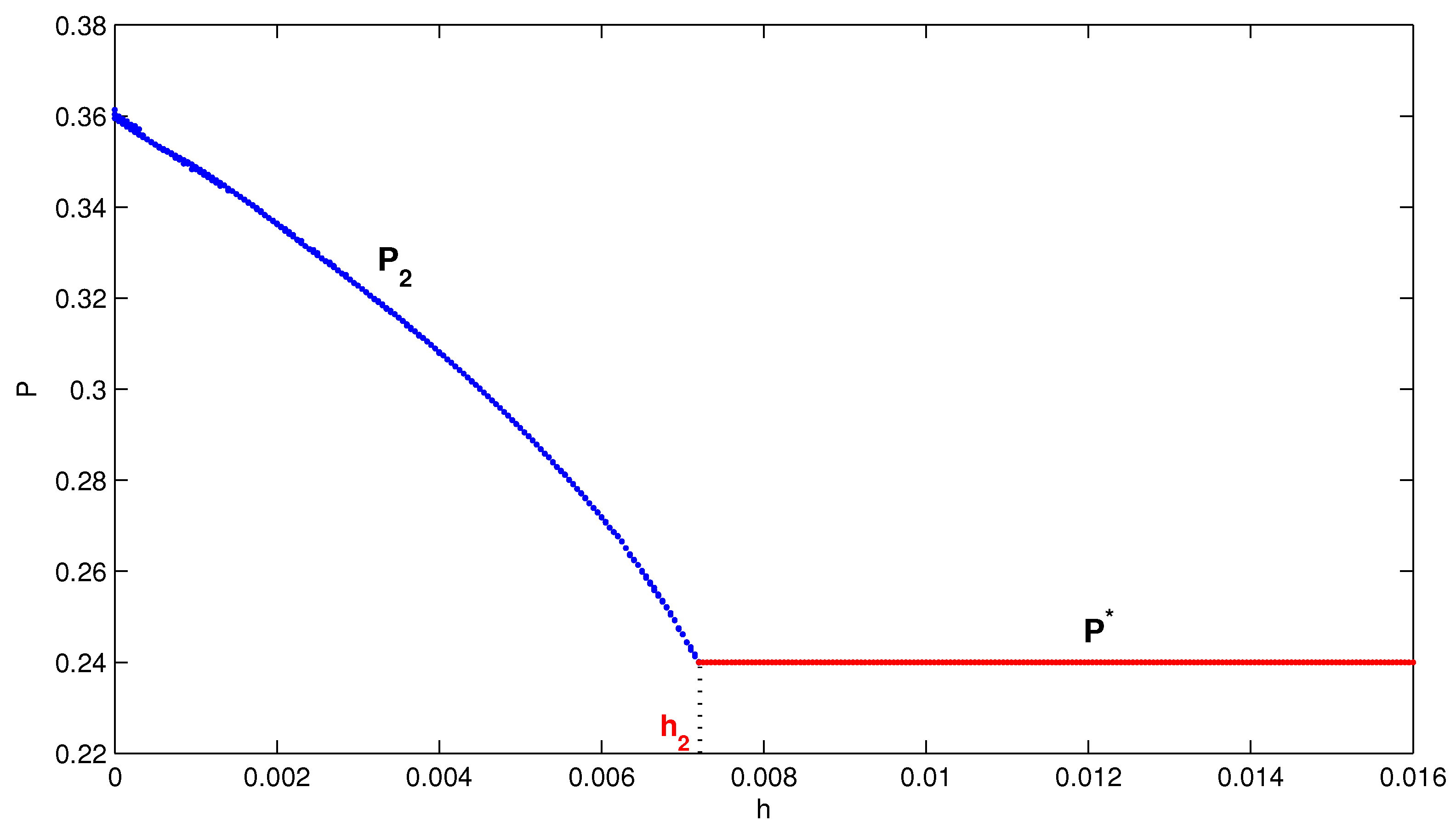
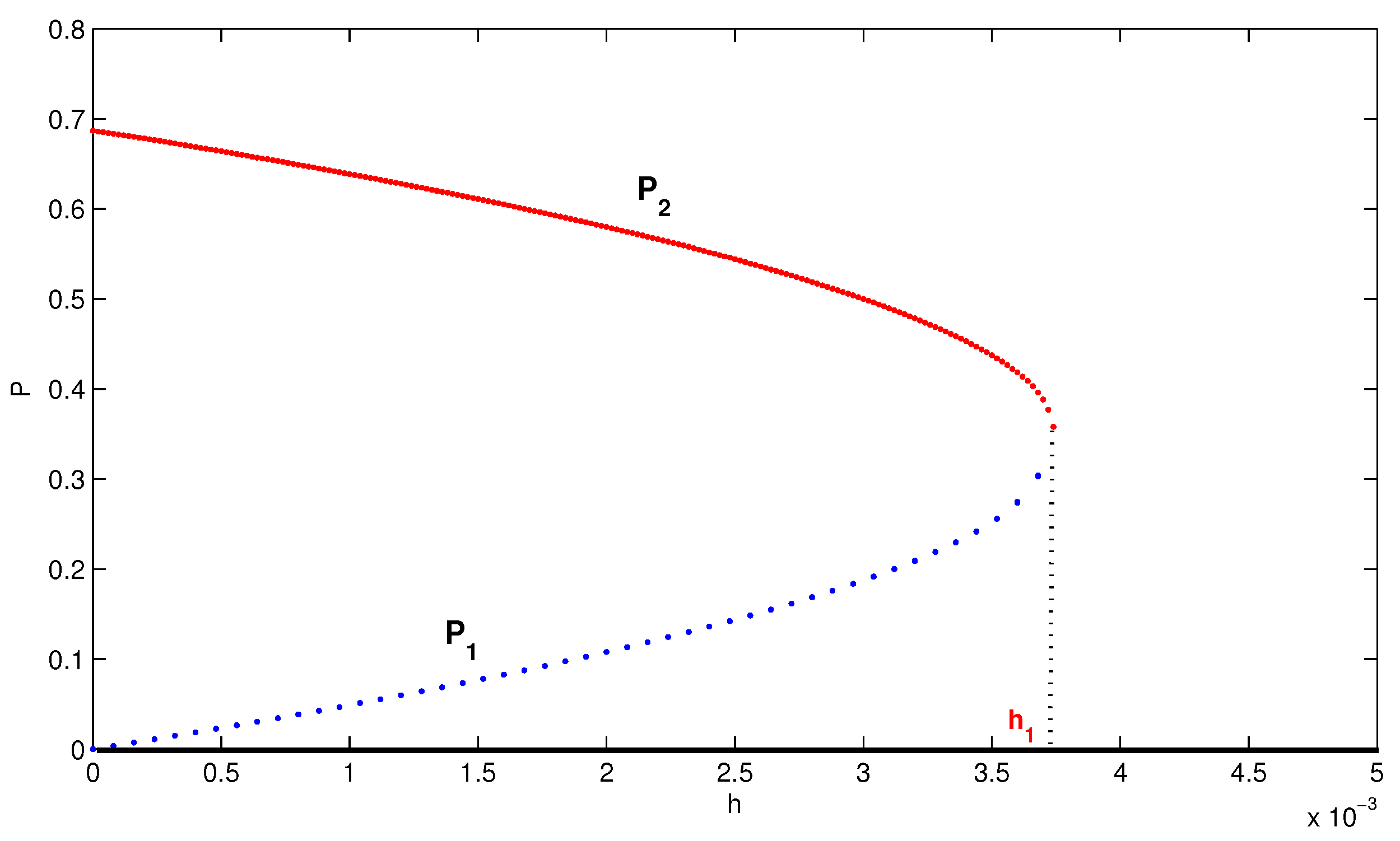
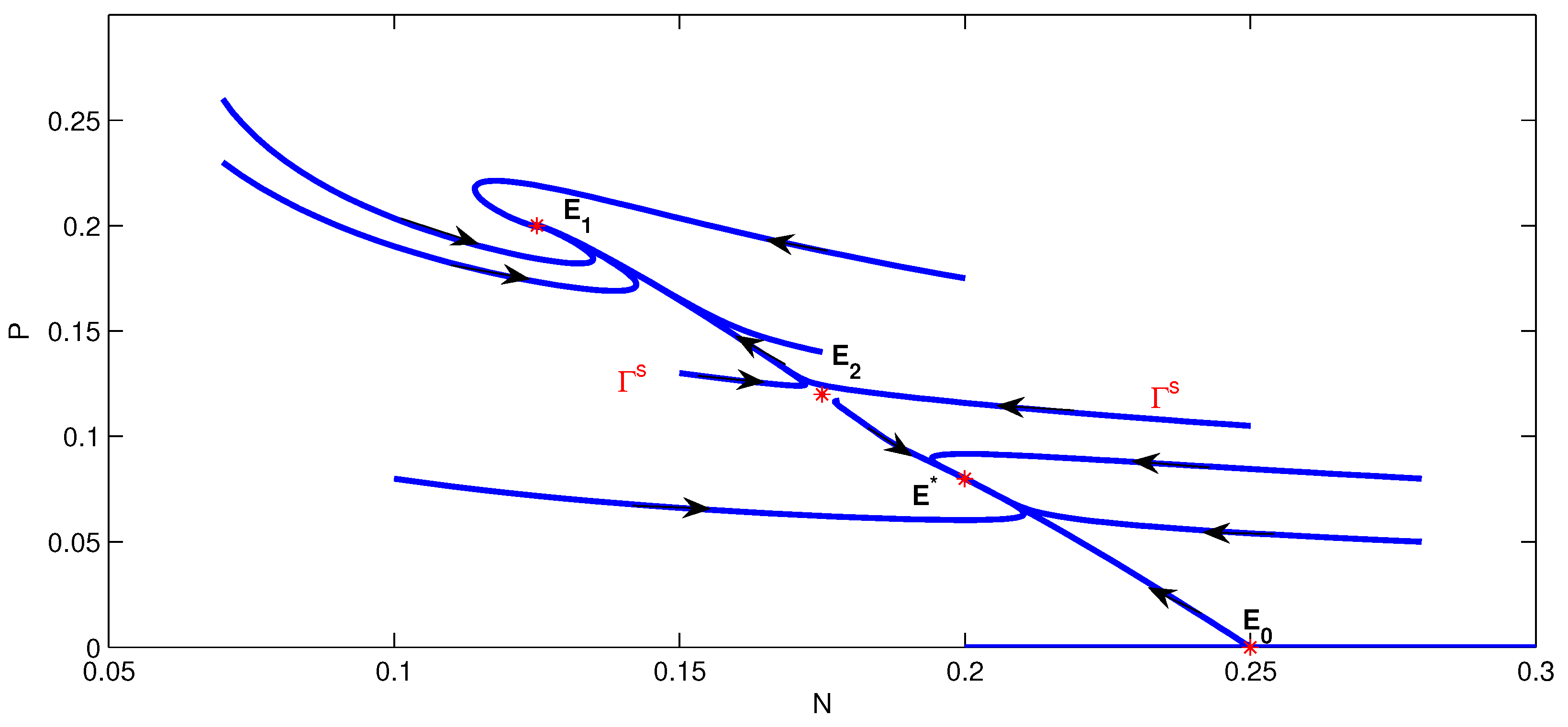
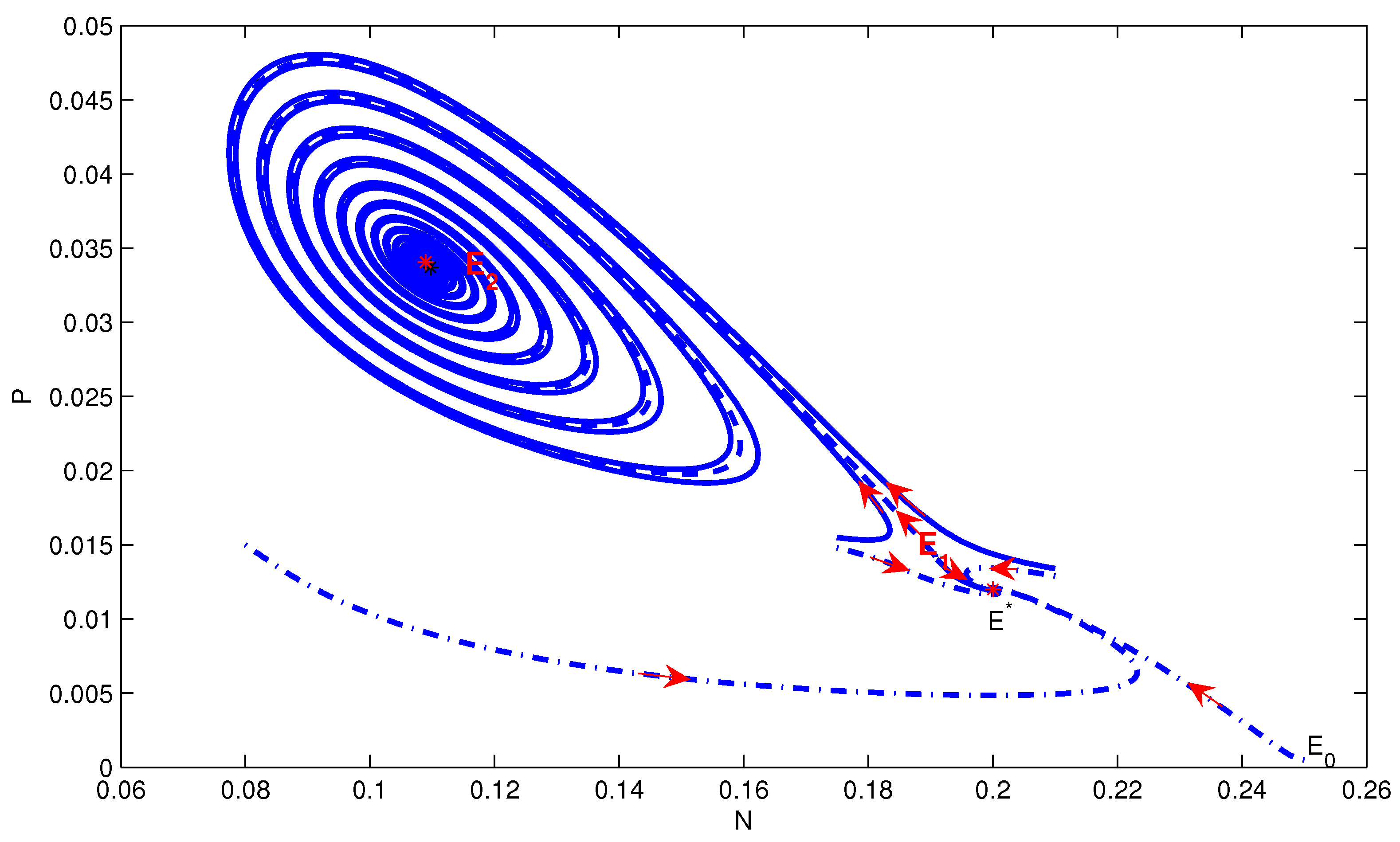
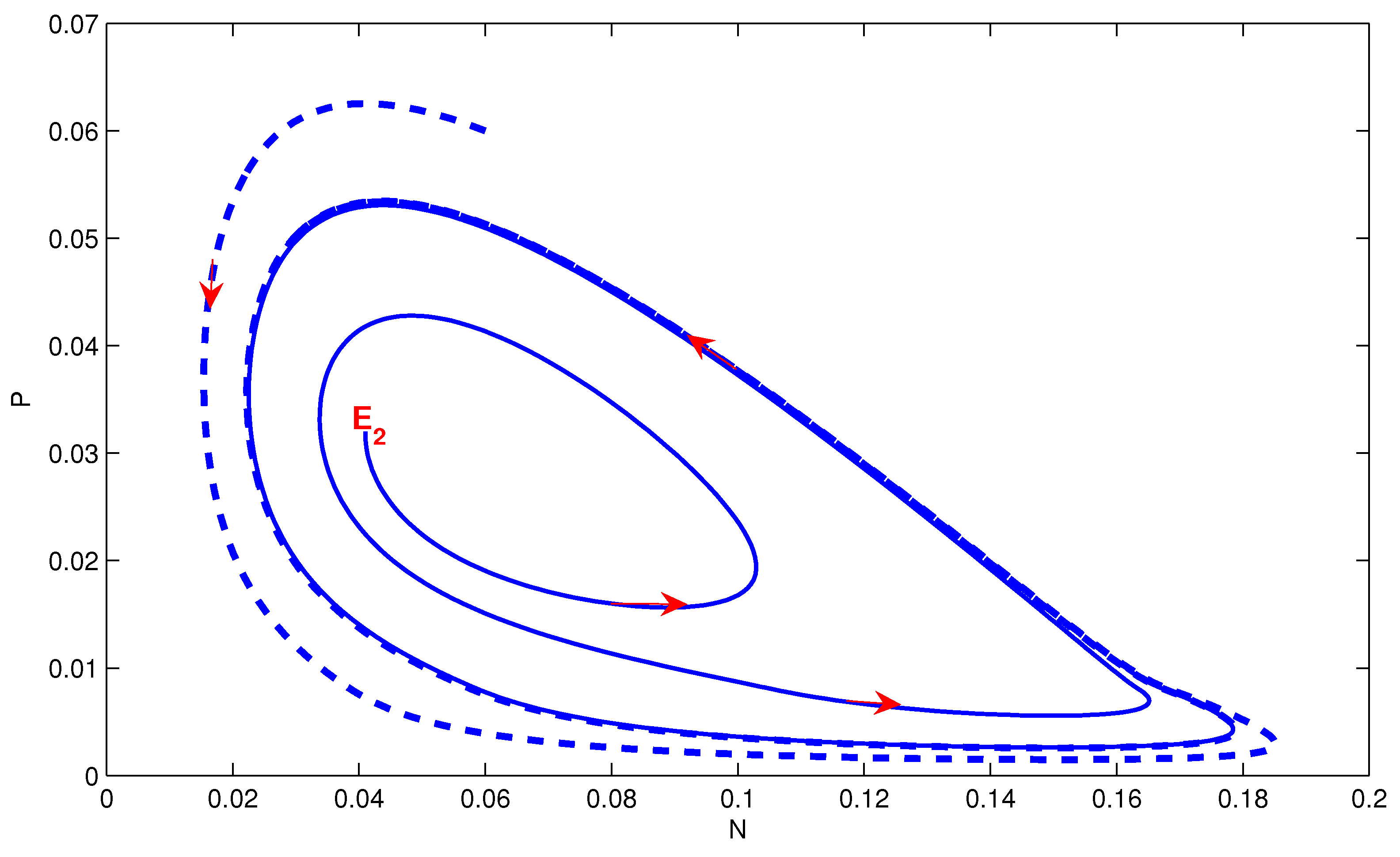
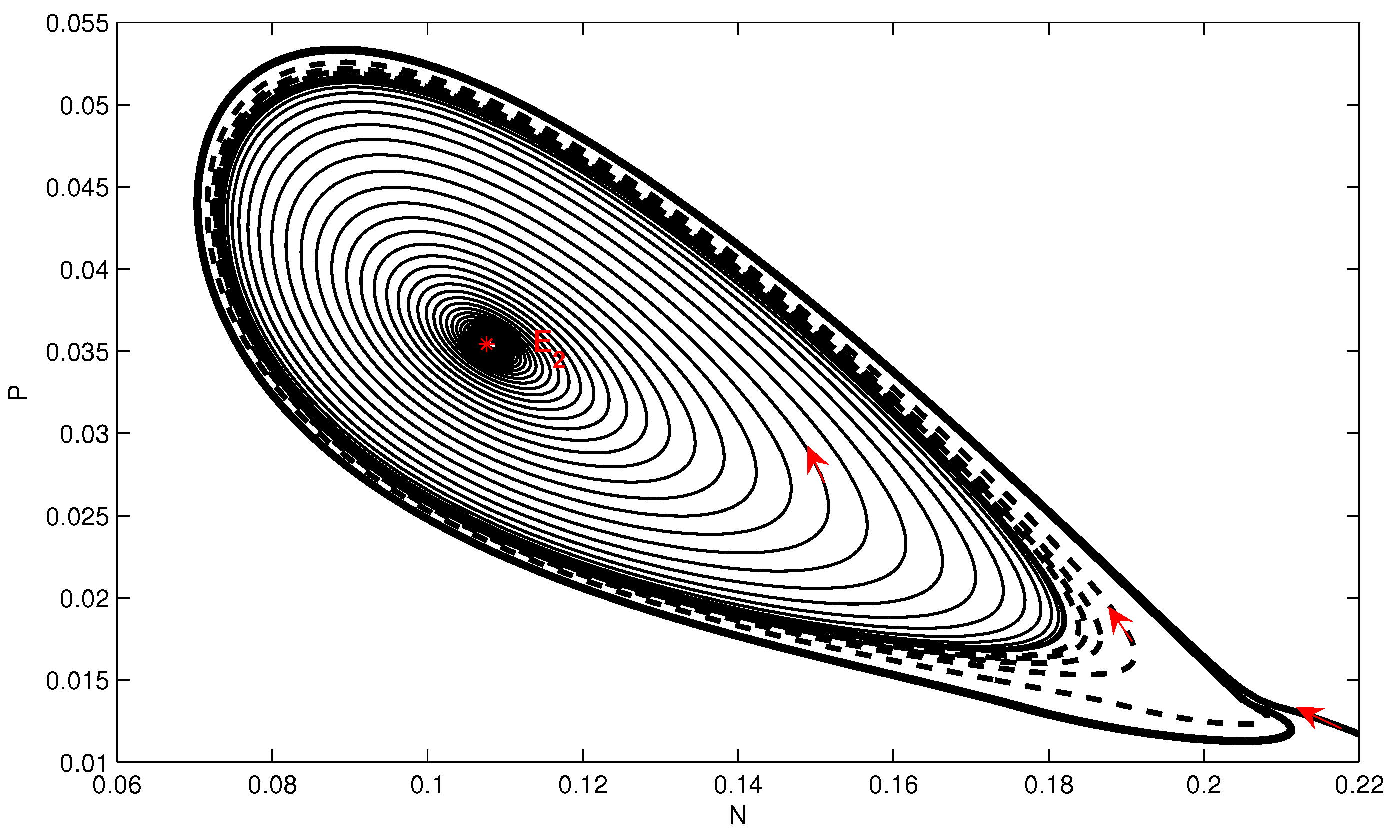
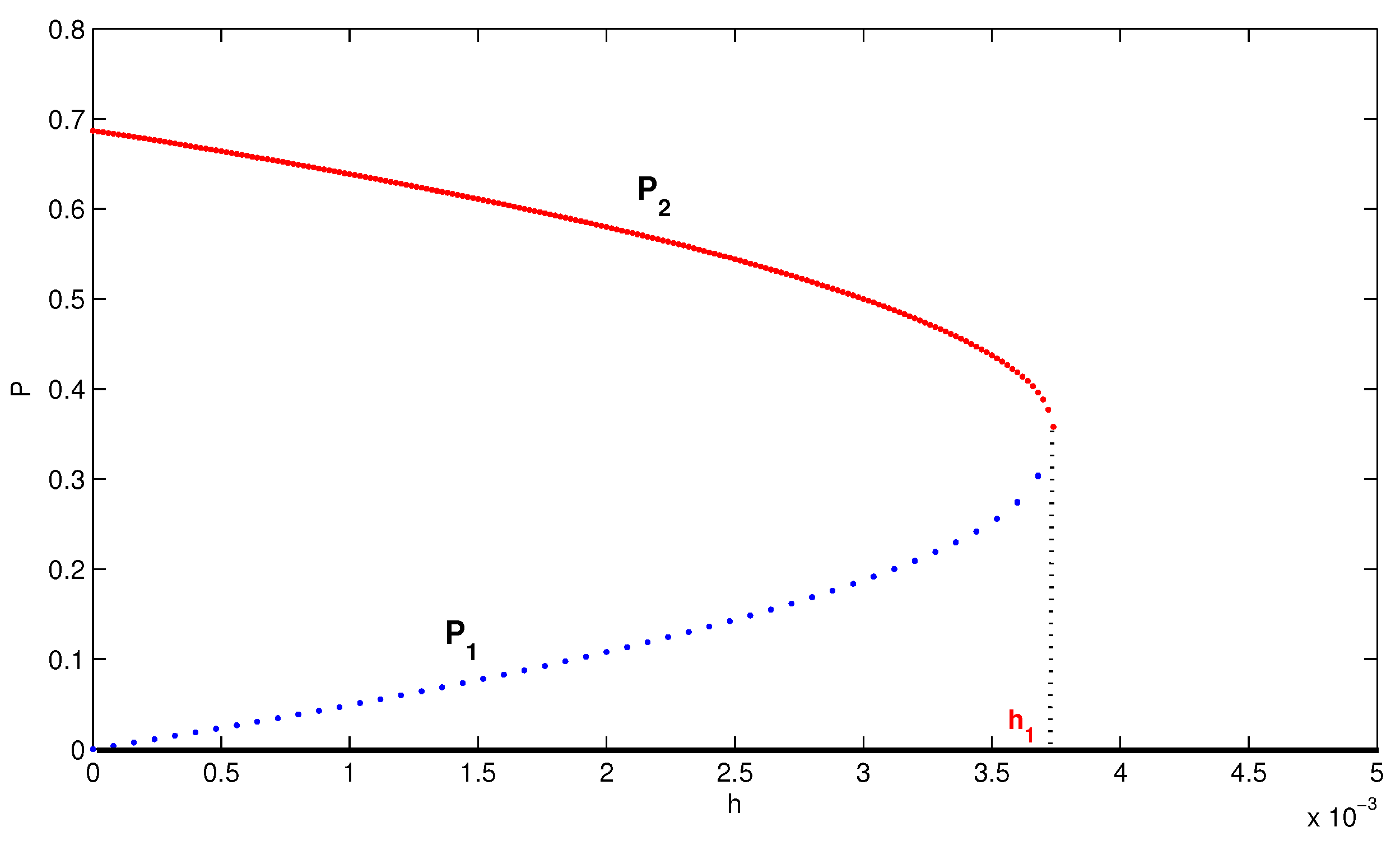
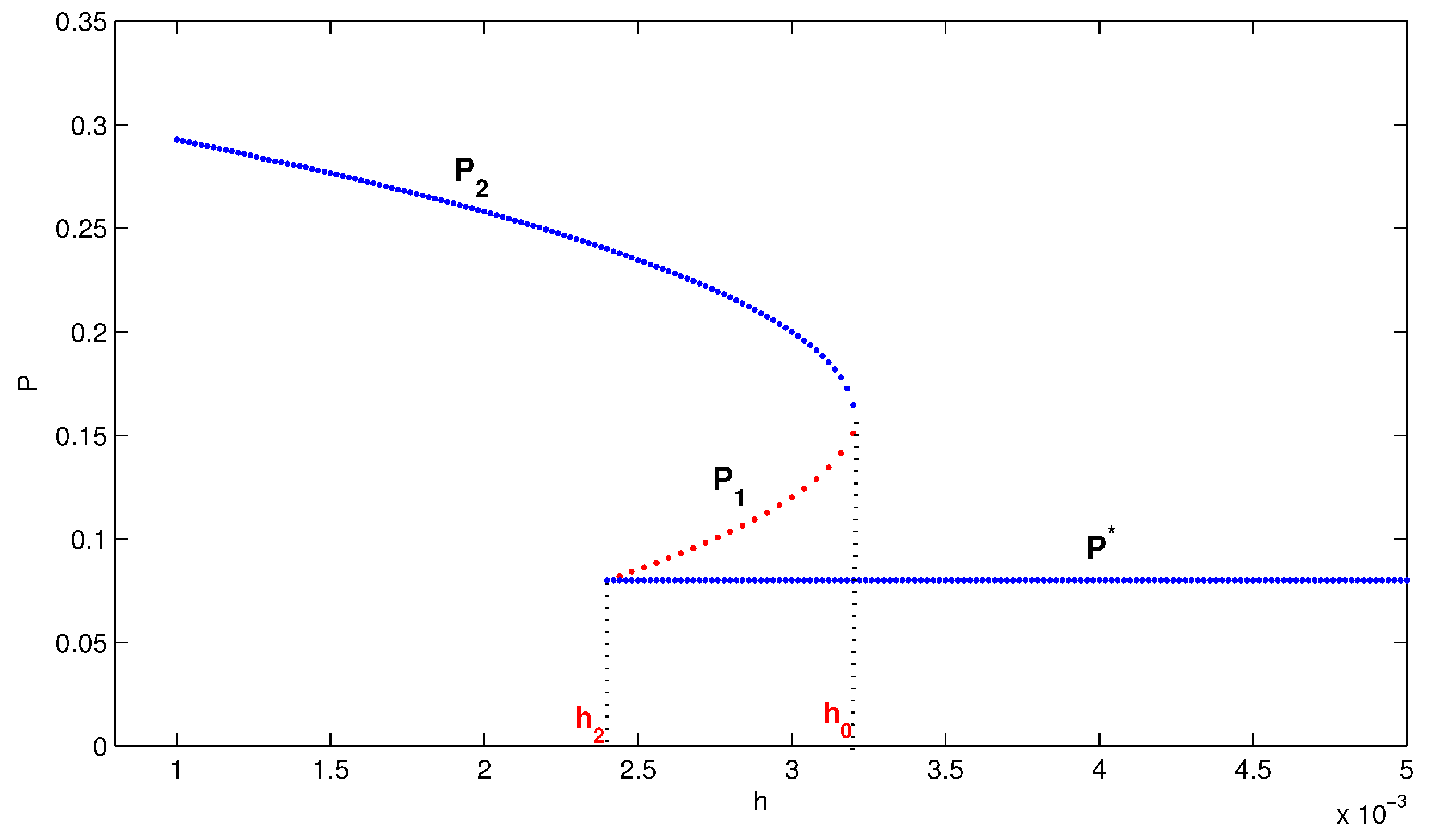
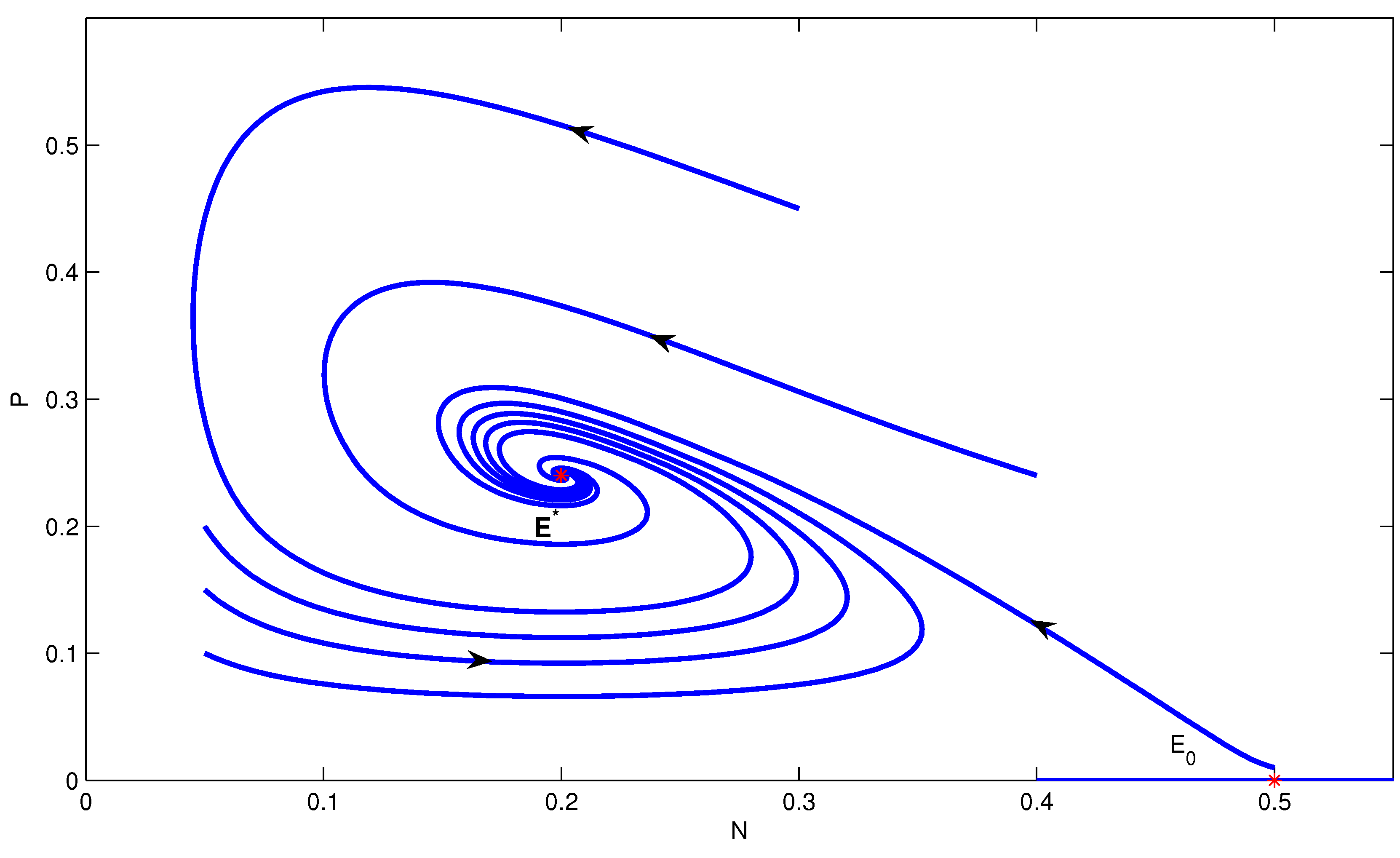
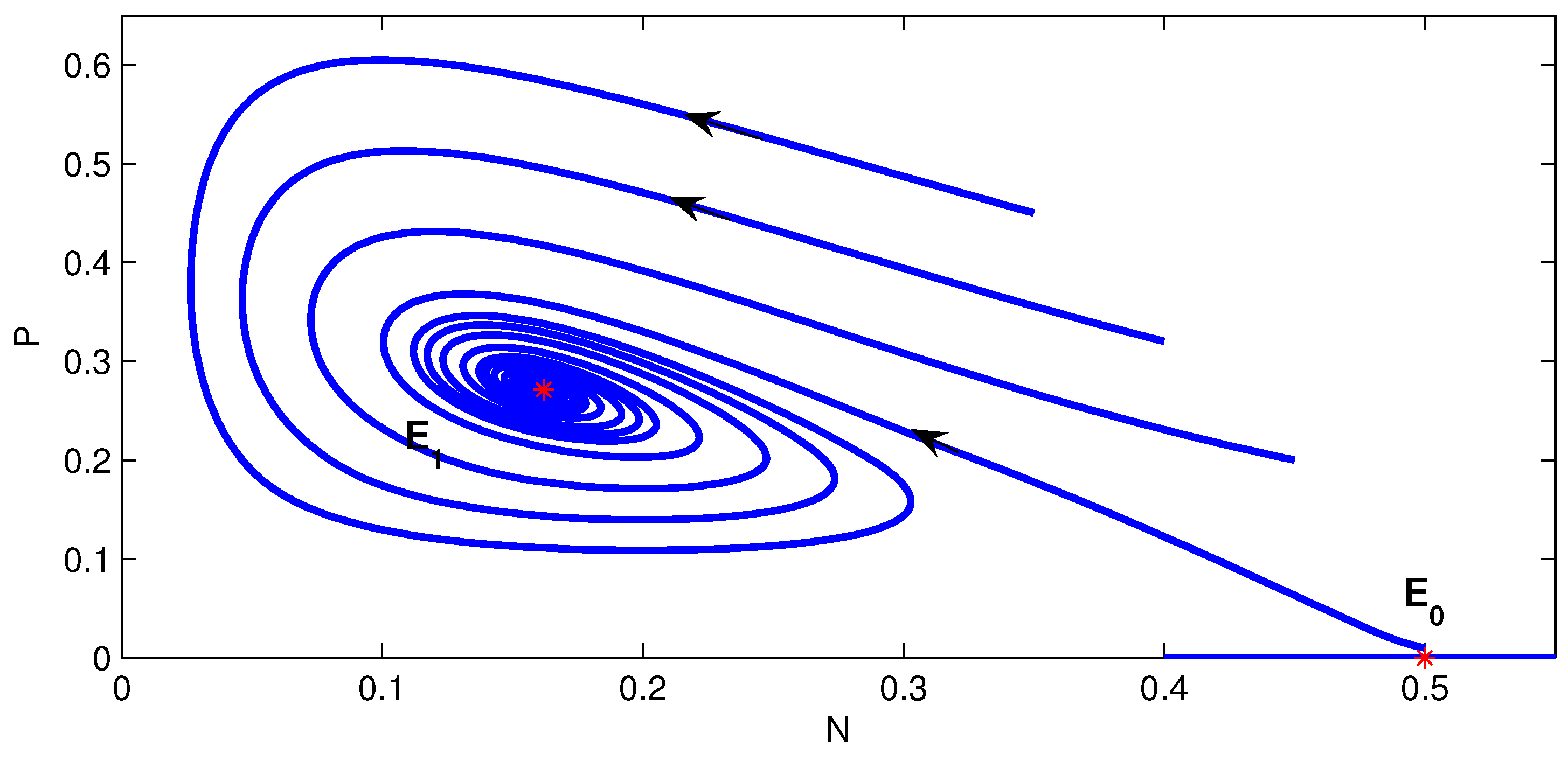
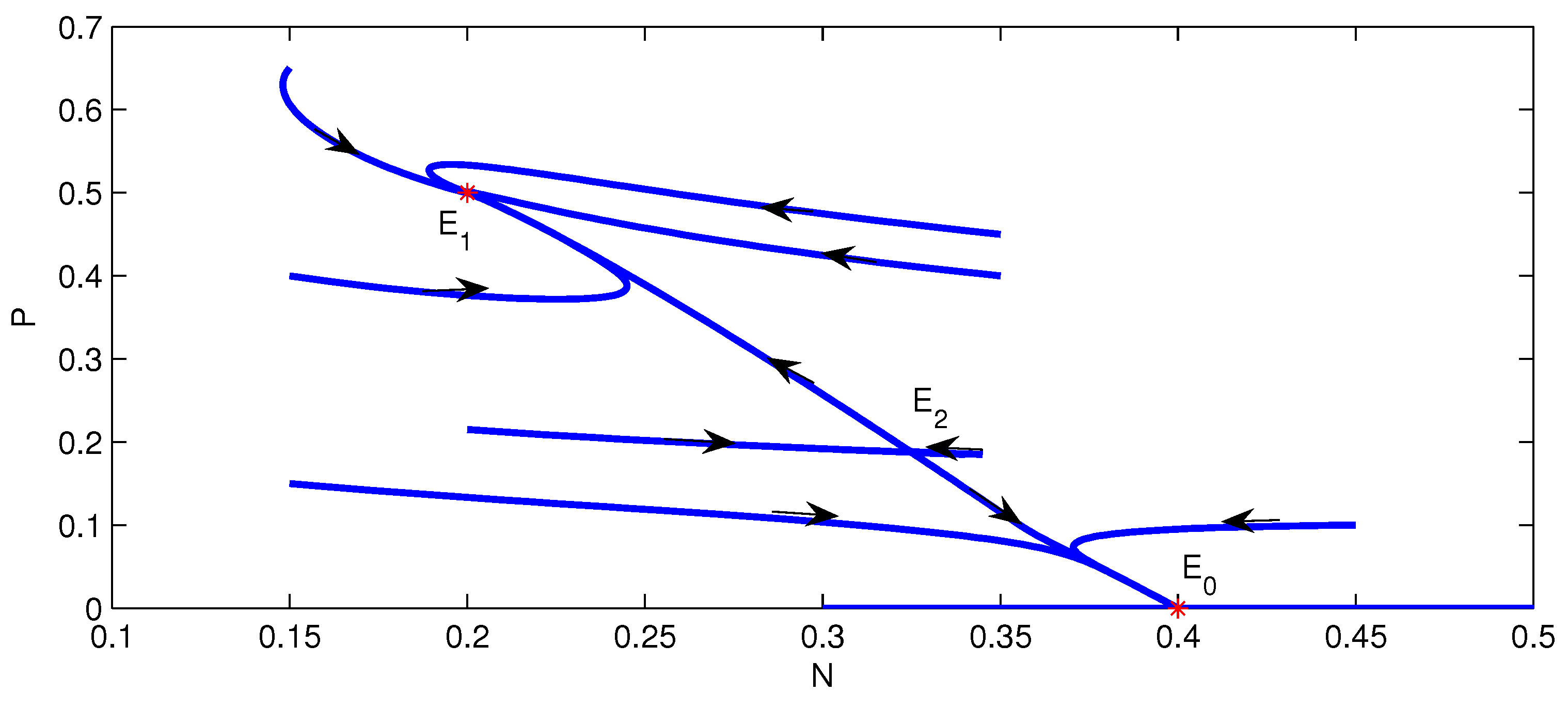
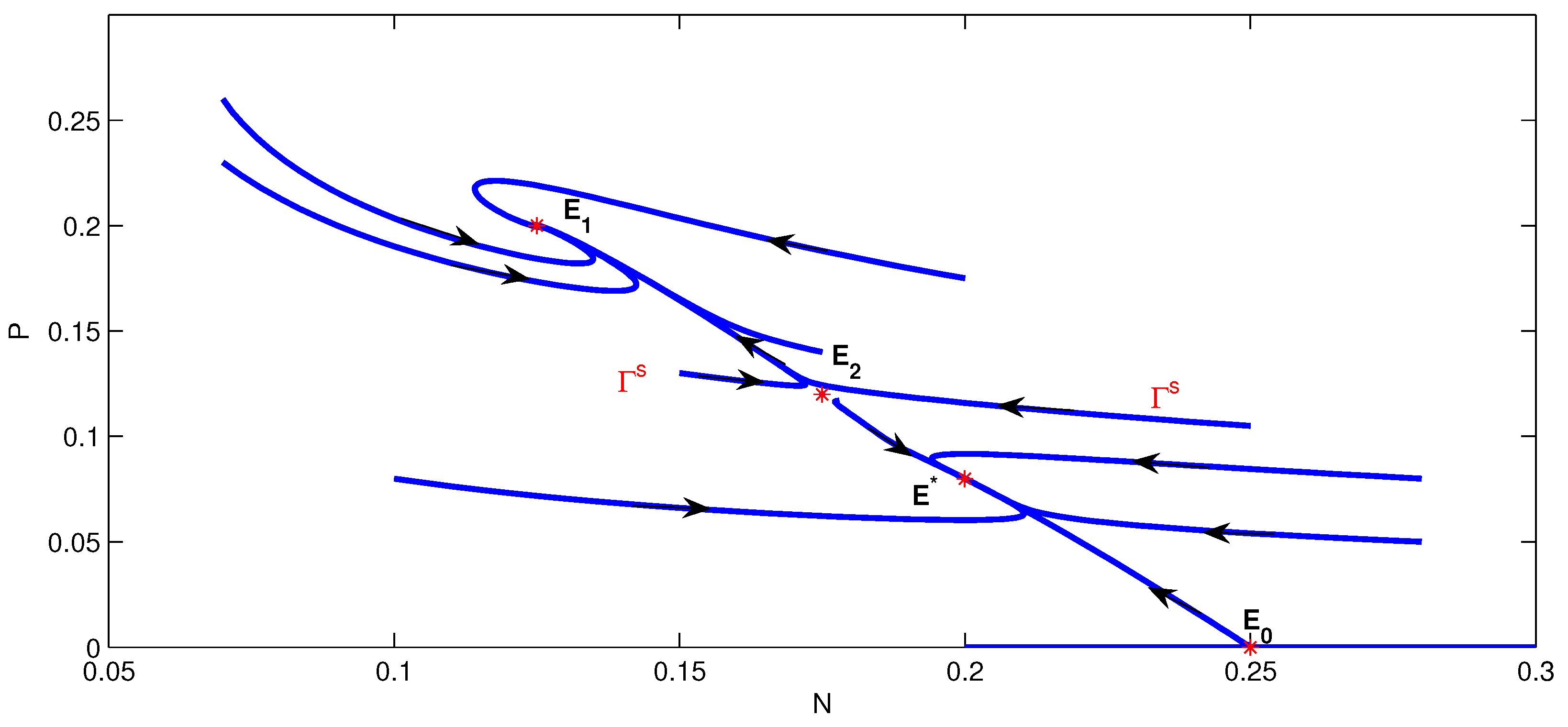
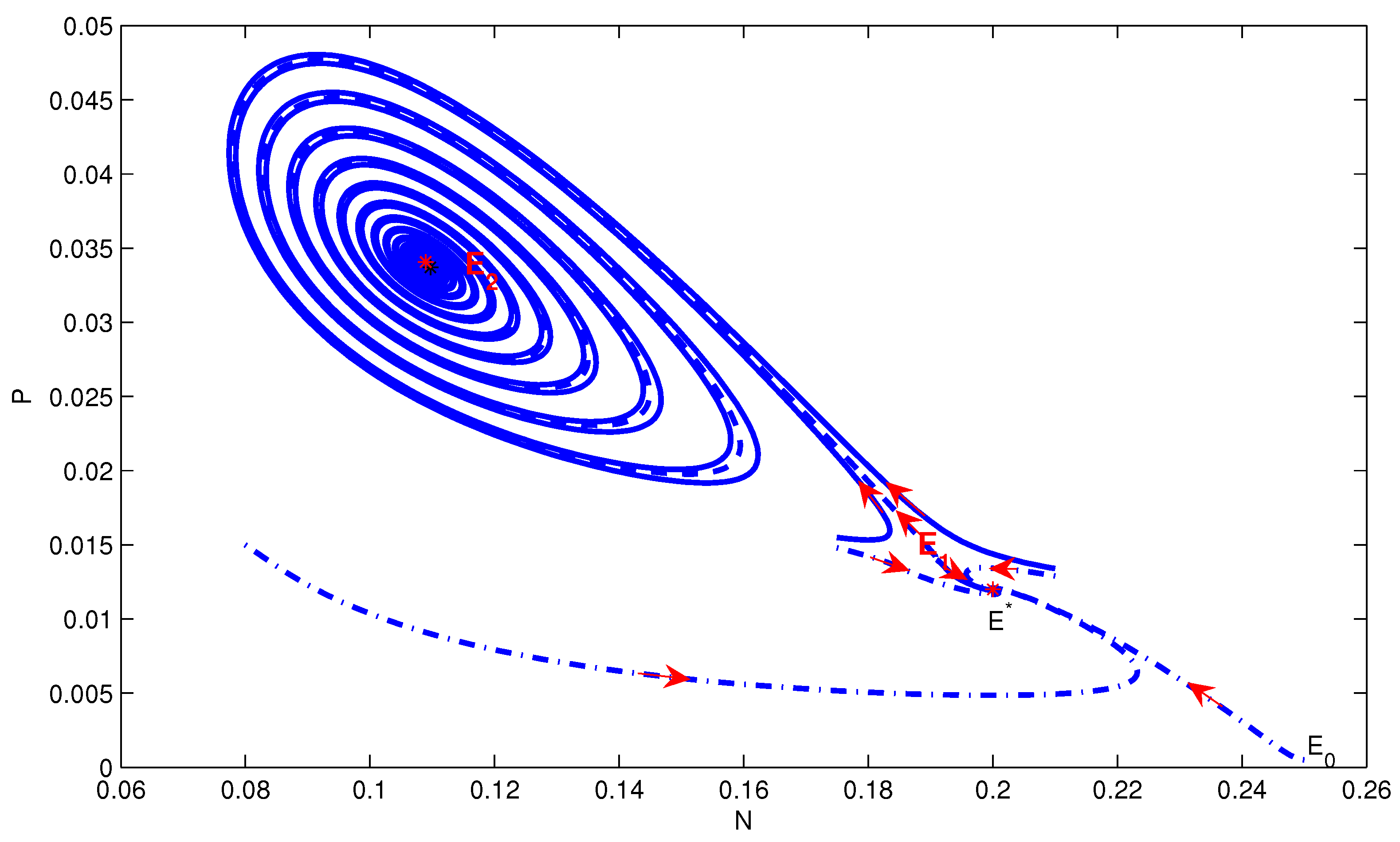
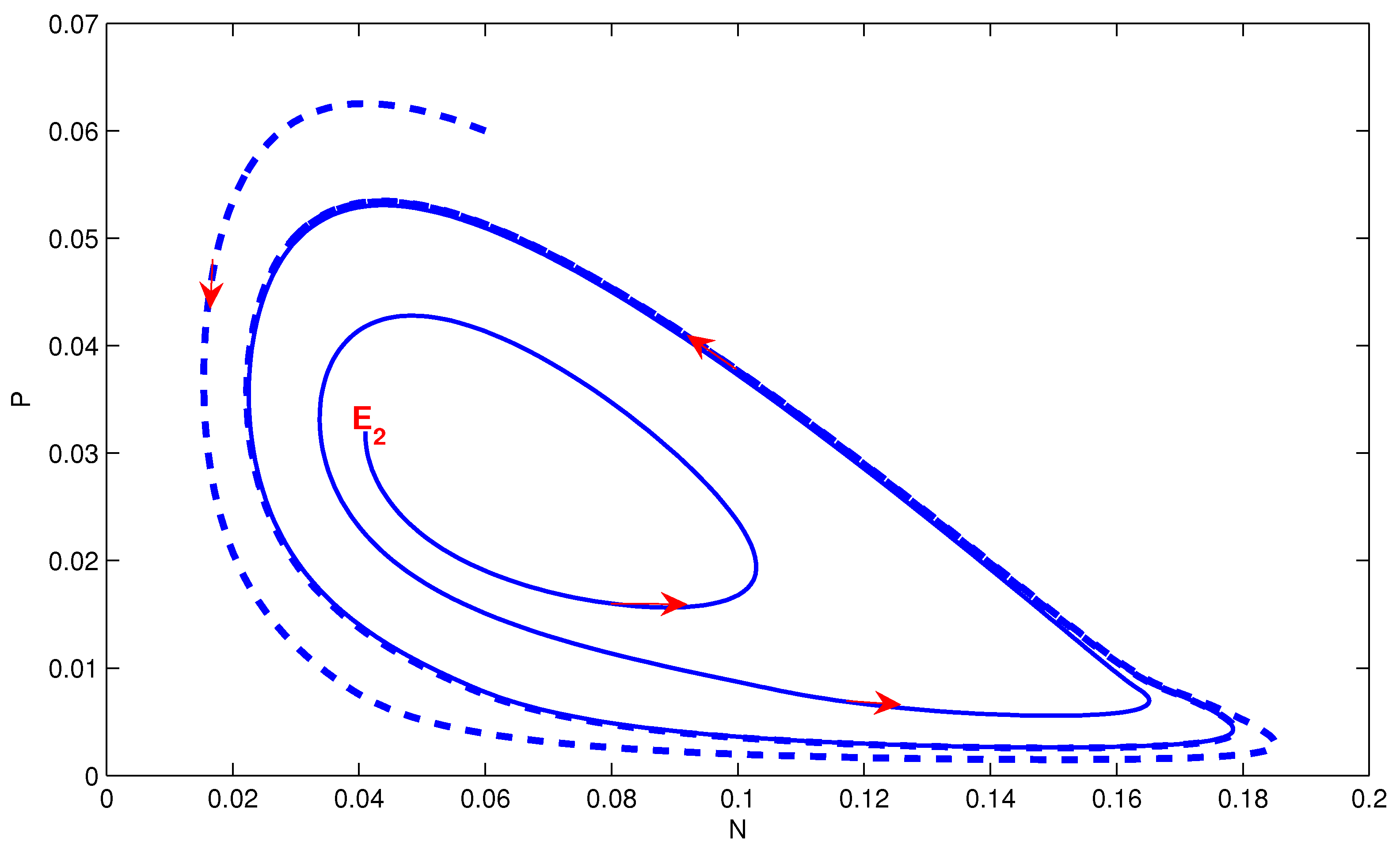
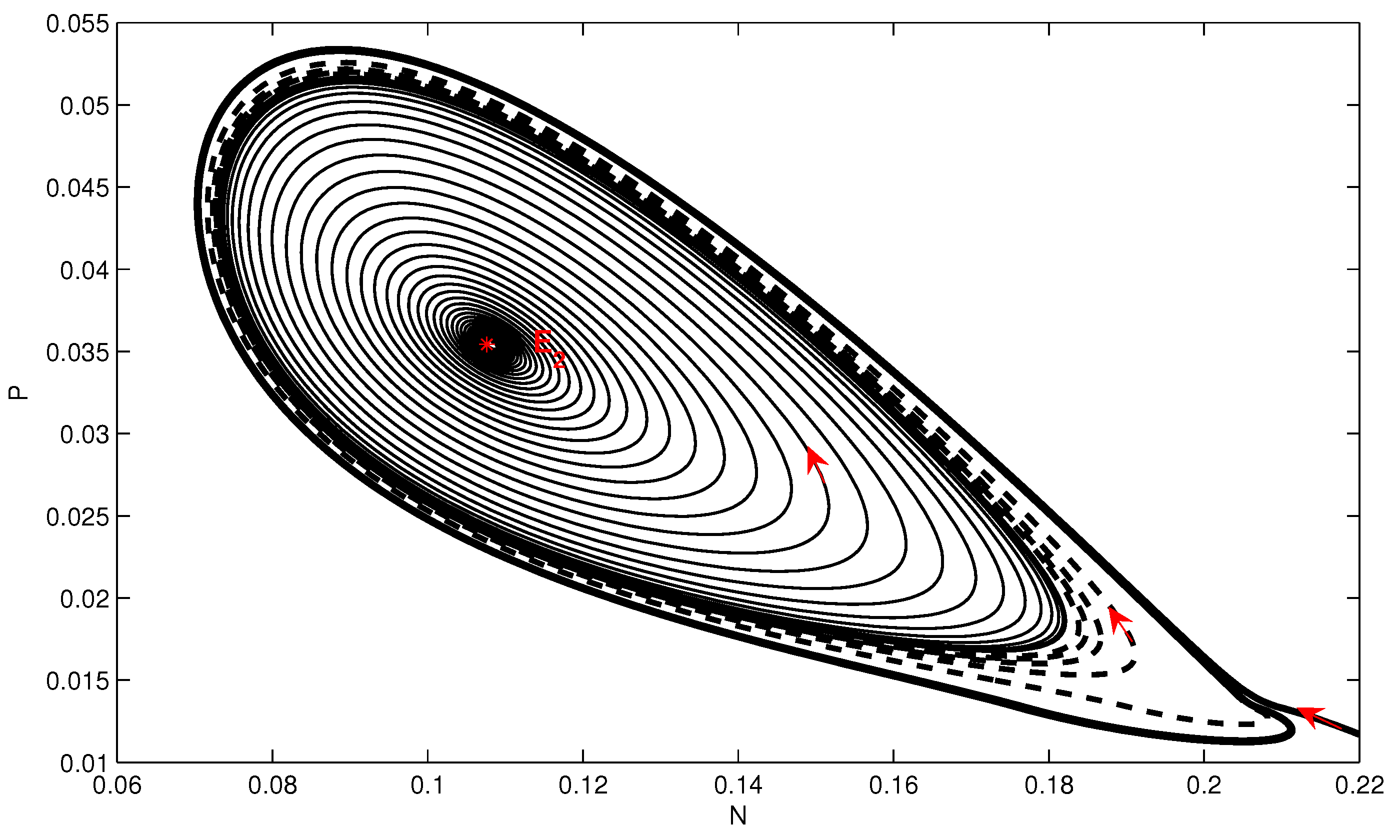
4. Numerical Simulation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Murdoch, W.W.; Briggs, C.J.; Nisbet, R.M. Consumer-Resource Dynamics; Princeton University Press: Princeton, NJ, USA, 2003. [Google Scholar]
- Seo, G.; DeAngelis, D.L. A predator-prey model with a Holling type I functional response including a predator mutual interference. J. Nonlinear Sci. 2011, 21, 811–833. [Google Scholar] [CrossRef]
- Turchin, P. Complex Population Dynamics: A Theoretical/empirical Synthesis; Princeton University Press: Princeton, NJ, USA, 2003. [Google Scholar]
- Gutierrez, A.P. Applied Population Ecology: A Supply-Demand Approach; John Wiley and Sons: New York, NY, USA, 1996. [Google Scholar]
- Seo, G.; Kot, M. A comparison of two predator-prey models with Hollingąŕs type I functional response. Math. Biosci. 2008, 212, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Cushing, J.M.; Zou, Y. The net reproductive value and stability in matrix population models. Nat. Resour. Model. 1994, 8, 297–333. [Google Scholar]
- Cushing, J.M. An Introduction to Structured Population Dynamics; SIAM: Philadelphia, PA, USA, 1998. [Google Scholar]
- Kar, T.K. Stability analysis of a prey-predator model incorporating a prey refuge. Commun. Nonlinear Sci. Numer. Simul. 2005, 10, 681–691. [Google Scholar] [CrossRef]
- Chakraborty, K.; Haldar, S.; Kar, T.K. Global stability and bifurcation analysis of a delay induced prey-predator system with stage structure. Nonlinear Dyn. 2013, 73, 1307–1325. [Google Scholar] [CrossRef]
- Liu, S.Q.; Beretta, E. A stage-structured predator-prey model of Beddington-Deangelis type. SIAM J. Appl. Math. 2006, 66, 1101–1129. [Google Scholar] [CrossRef]
- Qu, Y.; Wei, J.J. Bifurcation analysis in a time-delay model for preyĺCpredator growth with stage-structure. Nonlinear Dyn. 2007, 49, 285–294. [Google Scholar] [CrossRef]
- Zhang, X.A.; Chen, L.S.; Neumann, A.U. The stage-structured predator-prey model and optimal harvesting policy. Math. Biosci. 2000, 168, 201–210. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Q.L. Dynamic behavior in a delayed stage-structured population model with stochastic fluctuation and harvesting. Nonlinear Dyn. 2011, 66, 231–245. [Google Scholar] [CrossRef]
- Lai, X.H.; Liu, S.Q.; Lin, R.Z. Rich dynamical behaviours for predator-prey model with weak Allee effect. Appl. Anal. 2010, 89, 1271–1292. [Google Scholar] [CrossRef]
- Lin, R.Z.; Liu, S.Q.; Lai, X.H. Bifurcations of a predator-prey system with weak Allee effects. J. Korean Math. Soc. 2013, 50, 695–713. [Google Scholar] [CrossRef]
- Xiao, D.M.; Jennings, L.S. Bifurcations of a ratio-dependent predator-prey system with constant rate harvesting. SIAM J. Appl. Math. 2005, 65, 737–753. [Google Scholar] [CrossRef]
- Xiao, D.M.; Li, W.X.; Han, M.A. Dynamics in a ratio-dependent predator-prey model with predator harvesting. J. Math. Anal. Appl. 2006, 324, 14–29. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Q.L.; Zhang, X. Dynamical behavior of a class of prey-predator system with impulsive state feedback control and Beddington-DeAngelis functional response. Nonlinear Dyn. 2012, 70, 1511–1522. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, S.Q. Global stability for a predator-prey model with dispersal among patches. Abstr. Appl. Anal. 2014, 2014, 176493. [Google Scholar] [CrossRef]
- Martin, A.; Ruan, S.G. Predator-prey models with delay and prey harvesting. J. Math. Biol. 2001, 43, 247–267. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.J.; Chen, L.S. Periodic solution and heteroclinic bifurcation in a predatorĺCprey system with Allee effect and impulsive harvesting. Nonlinear Dyn. 2014, 76, 1109–1117. [Google Scholar] [CrossRef]
- Azar, C.; Holmberg, J.; Lindgren, K. Stability analysis of harvesting in a predator-prey model. J. Theor. Biol. 1995, 174, 13–19. [Google Scholar] [CrossRef]
- Brauer, F.; Soudack, A.C. Stability regions and transition phenomena for harvested predator-prey systems. J. Math. Biol. 1979, 7, 319–337. [Google Scholar] [CrossRef]
- Brauer, F.; Soudack, A.C. Stability regions in predator-prey systems with constant-rate prey harvesting. J. Math. Biol. 1979, 8, 55–71. [Google Scholar] [CrossRef]
- Kar, T.K. Selective harvesting in a prey-predator fishery with time delay. Math. Comput. Model. 2003, 38, 449–458. [Google Scholar] [CrossRef]
- Lenzini, P.; Rebaza, J. Nonconstant predator harvesting on ratio-dependent predator-prey models. Appl. Math. Sci. 2010, 4, 791–803. [Google Scholar]
- Negi, K.; Gakkhar, S. Dynamics in a Beddington-DeAngelis prey-predator system with impulsive harvesting. Ecol. Model. 2007, 206, 421–430. [Google Scholar] [CrossRef]
- Xiao, M.; Cao, J.D. Hopf bifurcation and non-hyperbolic equilibrium in a ratio-dependent predator-prey model with linear harvesting rate: Analysis and computation. Math. Comput. Model. 2009, 50, 360–379. [Google Scholar] [CrossRef]
- Beddington, J.R.; Cooke, J.K. Harvesting from a prey-predator complex. Ecol. Model. 1982, 14, 155–177. [Google Scholar] [CrossRef]
- Hu, Z.X.; Ma, W.B.; Ruan, S.G. Analysis of an SIR epidemic model with nonlinear incidence rate and treatment. Math. Biosci. 2012, 238, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.D. Backward bifurcation of an epidemic model with treatment. Math. Biosci. 2006, 201, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, X.N. Backward bifurcation and global dynamics of an SIS epidemic model with general incidence rate and treatment. Nonlinear Anal. Real World Appl. 2009, 10, 565–575. [Google Scholar] [CrossRef]
- Cantrell, R.S.; Cosner, C. On the dynamics of predator-prey models with the Beddington—DeAngelis functional response. J. Math. Anal. Appl. 2001, 257, 206–222. [Google Scholar] [CrossRef]
- Lu, Y.; Li, D.; Liu, S.Q. Modeling of hunting strategies of the predators in susceptible and infected prey. Appl. Math. Comput. 2016, 284, 268–285. [Google Scholar] [CrossRef]
- Hale, J.; Waltman, P. Persistence in infinite-dimensional systems. SIAM J. Math. Anal. 1989, 20, 388–395. [Google Scholar] [CrossRef]
- McCluskey, C. Lyapunov Functions for Tuberculosis Models with Fast and Slow Progression. Math. Biol. Eng. 2006, 3, 603–614. [Google Scholar] [CrossRef]
- Hale, J.; Lunel, S.V. Introduction to Functional Differential Equations; Springer-Verlag: New York, NY, USA, 1993. [Google Scholar]
- Freedman, H.I. Deterministic Mathematical Models in Population Ecology; Marcel Dekker: New York, NY, USA, 1980. [Google Scholar]
- Cui, J.; Mu, X.; Wan, H. Saturation recovery leads to multiple endemic equilibria and backward bifurcation. J. Theor. Biol. 2008, 254, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Van den Driessche, P.; Watmough, J. Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 2002, 180, 29–48. [Google Scholar] [CrossRef]
- McQuaid, C.F.; Britton, N.F. Trophic structure, stability, and parasite persistence threshold in food webs. Bull. Math. Biol. 2013, 75, 2196–2207. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, X.N. Backward bifurcation of an epidemic model with saturated treatment function. J. Math. Anal. Appl. 2008, 348, 433–443. [Google Scholar] [CrossRef]
- Zhou, L.H.; Fan, M. Dynamics of an SIR epidemic model with limited medical resources revisited. Nonlinear Anal. Real World Appl. 2012, 13, 312–324. [Google Scholar] [CrossRef]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Liu, S.; Cui, J.; Li, J. A Simple Predator-Prey Population Model with Rich Dynamics. Appl. Sci. 2016, 6, 151. https://doi.org/10.3390/app6050151
Li B, Liu S, Cui J, Li J. A Simple Predator-Prey Population Model with Rich Dynamics. Applied Sciences. 2016; 6(5):151. https://doi.org/10.3390/app6050151
Chicago/Turabian StyleLi, Bing, Shengqiang Liu, Jing’an Cui, and Jia Li. 2016. "A Simple Predator-Prey Population Model with Rich Dynamics" Applied Sciences 6, no. 5: 151. https://doi.org/10.3390/app6050151
APA StyleLi, B., Liu, S., Cui, J., & Li, J. (2016). A Simple Predator-Prey Population Model with Rich Dynamics. Applied Sciences, 6(5), 151. https://doi.org/10.3390/app6050151