
Dynamics of a Stochastic Intraguild Predation Model

Abstract
:1. Introduction
2. Preliminaries
- (i)
- If there exist two positive constants T and such thatfor all where , , are independent standard Brownian motions and , , are constants, then if or if
- (ii)
- If there exist three positive constants T, λ, and such thatfor all where , , are independent standard Browniam motions and , , are constants, then
- (H)
- In the domain U and some neighborhood thereof, the smallest eigenvalue of the diffusion matrix is bounded away from zero;
- (H)
- If , the mean time τ at which a path issuing from x reaches the set U is finite, and for every compact subset .
- (K)
- (K)
- To obtain (H), we only need to prove that there exist a neighborhood U and a nonnegative -function such that for any , (see [39]).
3. Stochastic Persistence and Stochastic Extinction
- (i)
- If , then all the populations are extinction a.s.
- (ii)
- If , then and are extinction a.s. and
- (iii)
- If , and , then is extinction a.s. and
- (iv)
- If , and , then is extinction a.s. and
- (v)
- If , , , then
- Case 1: if , then
- Case 2: if , then by Equation (7), for sufficiently large t, we getIt follows from Lemma 1 and the arbitrariness of ε thatOn the other hand, a direct calculation also shows thatwhere . For sufficiently large t, multiplying both sides of three equalities of Equation (21) by , and , respectively, and then adding these three equalities, we obtainHere, we have In fact, if , then , which implies that and This is a contradiction. By Equation (7), for sufficiently large t, we obtainFrom Lemma 1, we getsince ε is arbitrary. For the third equality of Equation (21) and sufficiently large t, combining Equation (33) with Equation (36) givesThen, if ε is sufficiently small.
4. Stationary Distribution and Ergodicity
5. Conclusions
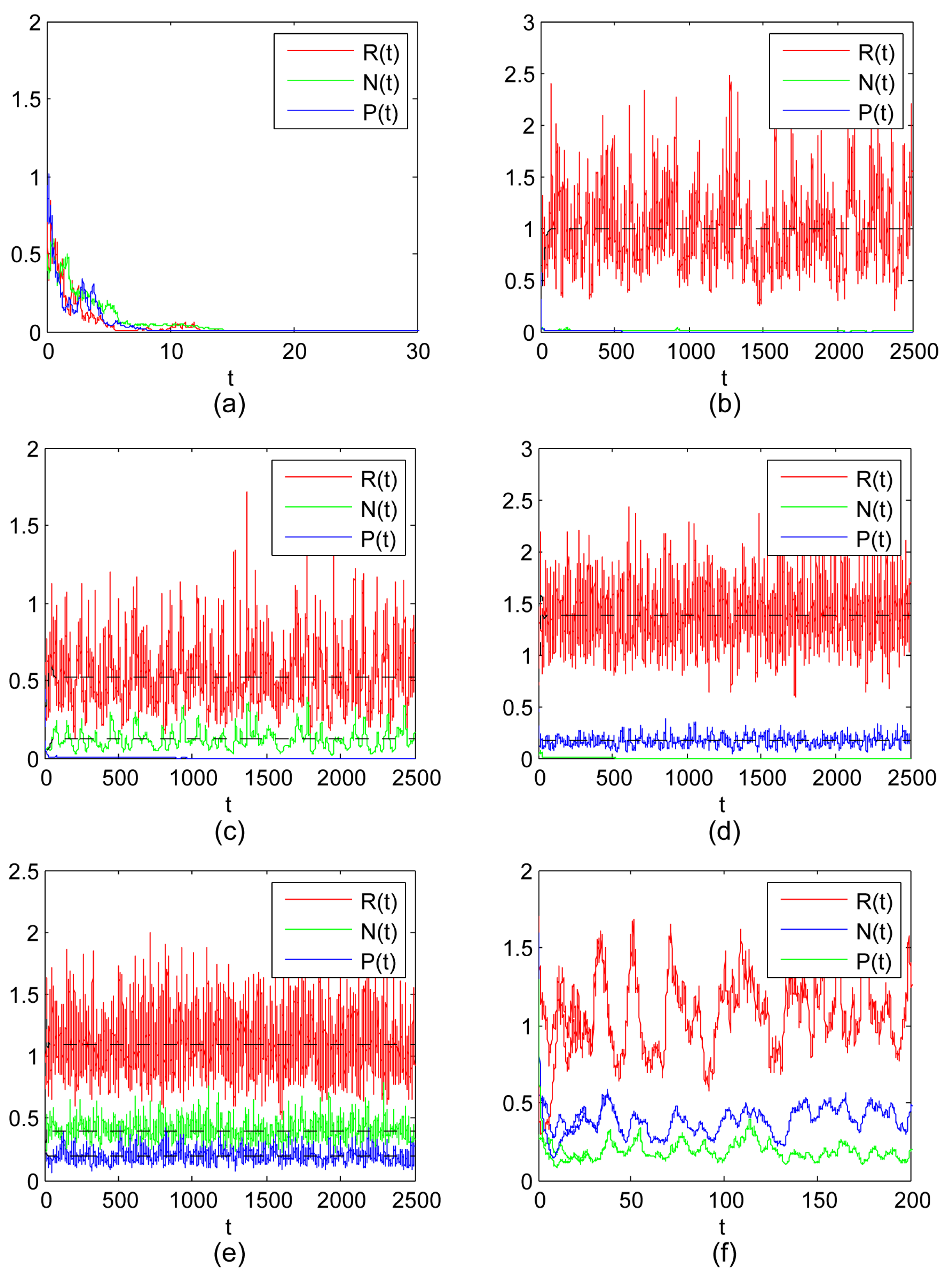
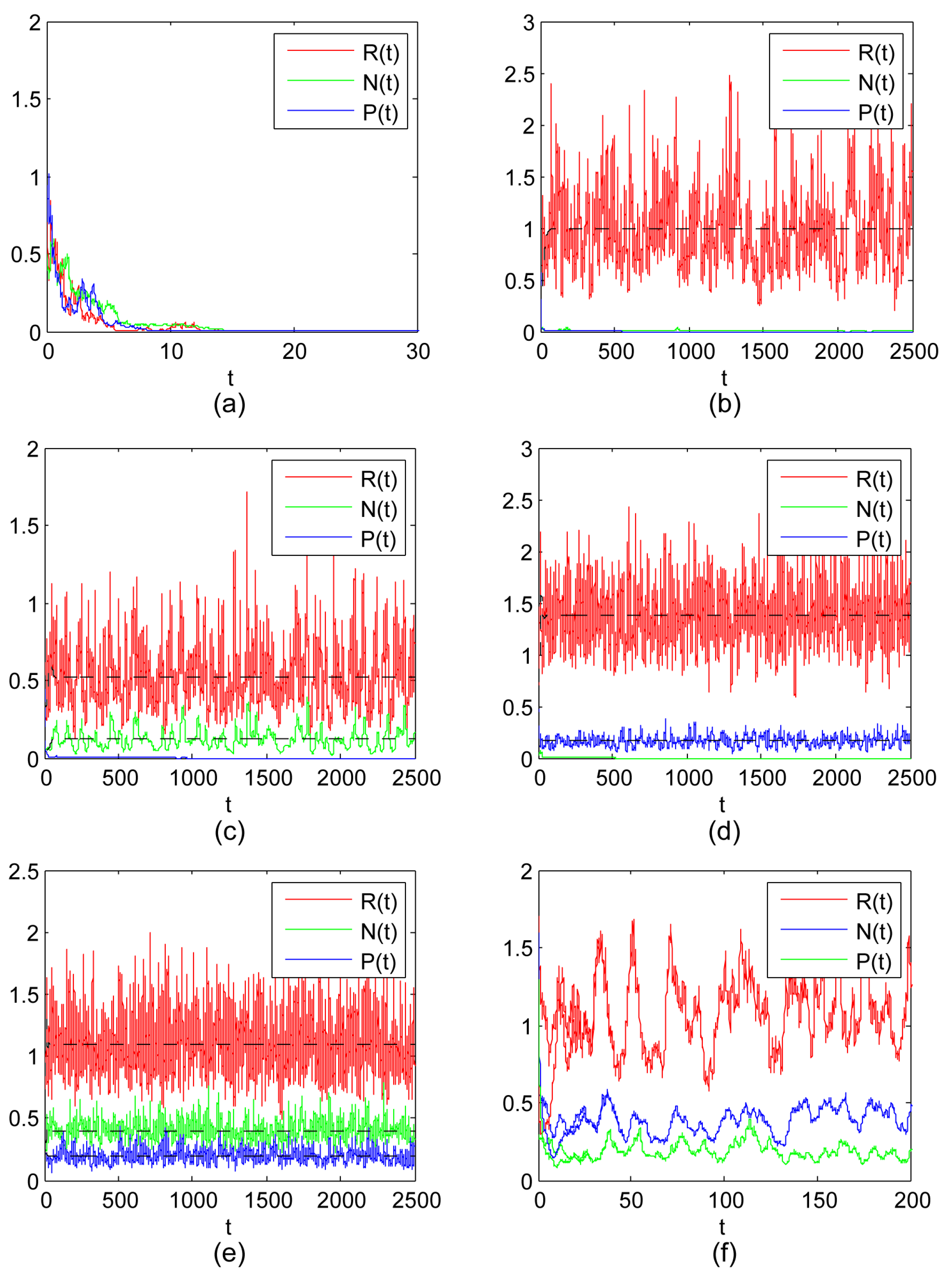
- The existence of both the shared prey and IG prey with the extinction of IG predators, and the existence of both the shared prey and IG predators with the extinction of IG prey are both possible outcomes of the stochastic model (3) with different sets of parameters (see Figure 1c,d). Here, it is worth noting that the noise has a negative effect for IG prey and IG predators, and may also have a positive effect for the shared prey if the values of and grow larger (see (iii) and (iv) of Theorem 8). This also implies that stochastic fluctuation of N or P would help R to grow larger;
- This study suggests that the shared prey, IG prey and IG predators can coexist together for the stochastic model (3), which implies that it is possible for the coexistence of three species under the influence of environmental noise (see Figure 1e). There is recognition that the noise may be favorable to three-species coexistence if , (see (v) of Theorem 8). In addition, we also prove that three-species is stable coexistence for the influence of environmental noise (see Theorem 10 and Figure 1f);
- The study of Theorem 11 suggests that the time average of the population size of model (3) with the development of time is equal to the stationary distribution in space.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Holt, R.D.; Polis, G.A. A theoretical framework for intraguild predation. Am. Nat. 1997, 149, 745–764. [Google Scholar] [CrossRef]
- Polis, G.A.; Holt, R.D. Intraguild predation: The dynamics of complex trophic interactions. Trends Ecol. Evol. 1992, 7, 151–154. [Google Scholar] [CrossRef]
- Hsu, S.B.; Ruan, S.G.; Yang, T.H. Analysis of three species Lotka–Volterra food web models with omnivory. J. Math. Anal. Appl. 2015, 426, 659–687. [Google Scholar] [CrossRef]
- Shchekinova, E.Y.; Löder, M.G.J.; Boersma, M.; Wiltshire, K.H. Facilitation of intraguild prey by its intraguild predator in a three-species Lotka–Volterra model. Theor. Popul. Biol. 2014, 92, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Velazquez, I.; Kaplan, D.; Velasco-Hernandez, J.X.; Navarrete, S.A. Multistability in an open recruitment food web model. Appl. Math. Comput. 2005, 163, 275–294. [Google Scholar] [CrossRef]
- Abrams, P.A.; Fung, S.R. Prey persistence and abundance in systems with intraguild predation and type-2 functional responses. J. Theor. Biol. 2010, 264, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Freeze, M.; Chang, Y.; Feng, W. Analysis of dynamics in a complex food chain with ratio-dependent functional response. J. Appl. Anal. Comput. 2014, 4, 69–87. [Google Scholar]
- Verdy, A.; Amarasekare, P. Alternative stable states in communities with intraguild predation. J. Theor. Biol. 2010, 262, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Urbani, P.; Ramos-Jiliberto, R. Adaptive prey behavior and the dynamics of intraguild predation systems. Ecol. Model. 2010, 221, 2628–2633. [Google Scholar] [CrossRef]
- Zabalo, J. Permanence in an intraguild predation model with prey switching. Bull. Math. Biol. 2012, 74, 1957–1984. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Kuang, Y.; Feng, Z.L. Cats protecting birds revisited. Bull. Math. Biol. 2005, 67, 1081–1106. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Wedekin, L. Dynamics of a intraguild predation model with generalist or specialist predator. J. Math. Biol. 2013, 67, 1227–1259. [Google Scholar] [CrossRef] [PubMed]
- Shu, H.Y.; Hu, X.; Wang, L.; Watmough, J. Delay induced stability switch, multitype bistability and chaos in an intraguild predation model. J. Math. Biol. 2015, 71, 1269–1298. [Google Scholar] [CrossRef] [PubMed]
- Golec, J.; Sathananthan, S. Stability analysis of a stochastic logistic model. Math. Comput. Model. 2003, 38, 585–593. [Google Scholar] [CrossRef]
- Jiang, D.Q.; Shi, N.Z.; Li, X.Y. Global stability and stochastic permanence of a non-autonomous logistic equation with random perturbation. J. Math. Anal. Appl. 2008, 340, 588–597. [Google Scholar] [CrossRef]
- Liu, M.; Wang, K.; Hong, Q. Stability of a stochastic logistic model with distributed delay. Math. Comput. Model. 2013, 57, 1112–1121. [Google Scholar] [CrossRef]
- Aguirre, P.; González-Olivares, E.; Torres, S. Stochastic predator–prey model with Allee effect on prey. Nonlinear Anal. RWA 2013, 14, 768–779. [Google Scholar] [CrossRef]
- Ji, C.Y.; Jiang, D.Q. Dynamics of a stochastic density dependent predator–prey system with Beddington-DeAngelis functional response. J. Math. Anal. Appl. 2011, 381, 441–453. [Google Scholar] [CrossRef]
- Liu, M.; Wang, K. Global stability of a nonlinear stochastic predator–prey system with Beddington-DeAngelis functional response. Commun. Nonlinear Sci. 2011, 16, 1114–1121. [Google Scholar] [CrossRef]
- Mandal, P.S.; Banerjee, M. Stochastic persistence and stability analysis of a modified Holling-Tanner model. Math. Method. Appl. Sci. 2013, 36, 1263–1280. [Google Scholar] [CrossRef]
- Saha, T.; Chakrabarti, C. Stochastic analysis of prey-predator model with stage structure for prey. J. Appl. Math. Comput. 2011, 35, 195–209. [Google Scholar] [CrossRef]
- Vasilova, M. Asymptotic behavior of a stochastic Gilpin-Ayala predator–prey system with time-dependent delay. Math. Comput. Model. 2013, 57, 764–781. [Google Scholar] [CrossRef]
- Yagi, A.; Ton, T.V. Dynamic of a stochastic predator–prey population. Appl. Math. Comput. 2011, 218, 3100–3109. [Google Scholar] [CrossRef]
- Jovanović, M.; Vasilova, M. Dynamics of non-autonomous stochastic Gilpin-Ayala competition model with time-varying delays. Appl. Math. Comput. 2013, 219, 6946–6964. [Google Scholar] [CrossRef]
- Li, X.; Mao, X. Population dynamical behavior of non-autonomous Lotka–Volterra competitive system with random perturbation. Discrete Cont. Dyn. Syst. Ser. 2009, 24, 523–593. [Google Scholar]
- Lian, B.S.; Hu, S.G. Asymptotic behaviour of the stochastic Gilpin-Ayala competition models. J. Math. Anal. Appl. 2008, 339, 419–428. [Google Scholar] [CrossRef]
- Zhu, C.; Yin, G. On competitive Lotka–Volterra model in random environments. J. Math. Anal. Appl. 2009, 357, 154–170. [Google Scholar] [CrossRef]
- Ji, C.Y.; Jiang, D.Q.; Liu, H.; Yang, Q.S. Existence, uniqueness and ergodicity of positive solution of mutualism system with stochastic perturbation. Math. Probl. Eng. 2010, 2010. [Google Scholar] [CrossRef]
- Liu, M.; Wang, K. Analysis of a stochastic autonomous mutualism model. J. Math. Anal. Appl. 2013, 402, 392–403. [Google Scholar] [CrossRef]
- Liu, M.; Wang, K. Population dynamical behavior of Lotka–Volterra cooperative systems with random perturbations. Discrete Cont. Dyn. Syst. Ser. 2013, 33, 2495–2522. [Google Scholar] [CrossRef]
- Liu, Q. Analysis of a stochastic non-autonomous food-limited Lotka–Volterra cooperative model. Appl. Math. Comput. 2015, 254, 1–8. [Google Scholar] [CrossRef]
- Liu, M.; Wang, K. Dynamics of a two-prey one-predator system in random environments. J. Nonlinear Sci. 2013, 23, 751–775. [Google Scholar] [CrossRef]
- May, R.M. Stability and Complexity in Model Ecosystems; Princeton University Press: Princeton, NJ, USA, 1973. [Google Scholar]
- Has’minskii, R.Z. Stochastic Stability of Differential Equations; Sijthoff & Noordhoff: Alphen aan den Rijn, the Netherlands, 1980. [Google Scholar]
- Karatzas, I.; Shreve, S. Brownian Motion and Stochastic Calculus; Springer: Berlin, Germany, 1991. [Google Scholar]
- Barbalat, I. Systems d’equations differentielles d’oscillations nonlineaires. Rev. Roum. Math. Pures Appl. 1959, 4, 267–270. [Google Scholar]
- Gard, T.C. Introduction to Stochastic Differential Equations; Marcel Dekker, Inc.: New York, NY, USA, 1988. [Google Scholar]
- Strang, G. Linear Algebra and Its Applications; Wellesley-Cambridge Press: London, UK, 1988. [Google Scholar]
- Zhu, C.; Yin, G. Asymptotic properties of hybrid diffusion systems. SIAM J. Control Optim. 2007, 46, 1155–1179. [Google Scholar] [CrossRef]

{kind=link}
| Equilibria | Existence | Local Stability |
|---|---|---|
| Always | Never | |
| Always | ||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, Z.; Cui, H.; Zhang, J. Dynamics of a Stochastic Intraguild Predation Model. Appl. Sci. 2016, 6, 118. https://doi.org/10.3390/app6040118
Xing Z, Cui H, Zhang J. Dynamics of a Stochastic Intraguild Predation Model. Applied Sciences. 2016; 6(4):118. https://doi.org/10.3390/app6040118
Chicago/Turabian StyleXing, Zejing, Hongtao Cui, and Jimin Zhang. 2016. "Dynamics of a Stochastic Intraguild Predation Model" Applied Sciences 6, no. 4: 118. https://doi.org/10.3390/app6040118
APA StyleXing, Z., Cui, H., & Zhang, J. (2016). Dynamics of a Stochastic Intraguild Predation Model. Applied Sciences, 6(4), 118. https://doi.org/10.3390/app6040118