1. Introduction
In Poland, the number of bee colonies has reached 2.35 million [
1], marking a 100% increase over the past two decades. This growth is accompanied by a 100% increase in the average density of colonies per unit area, which currently stands at 7.7 colonies/km
2. In comparison, the average density in the 27 EU countries is 4.8 colonies per square kilometer, which is higher than in China (approximately 1.3 colonies/km
2 if we consider only
Apis mellifera) [
2]. The density calculation for China considered the fact that 52% of China’s area is covered by mountains [
3], where bees are not used. In certain EU countries, the density of colonies per km
2 is higher than in Poland: Hungary (10.6), Greece (16.8), and Malta (19.2). Turkey, the third largest honey producer in the world after China and the EU, also has a high density of 11.1 colonies per km
2. All the above calculations are based on [
4,
5].
It is a commonly held belief that the primary purpose of keeping honeybees is to produce honey. However, it is estimated that the benefits to humanity from bee pollination are significantly greater [
6,
7,
8,
9]. In certain regions of the world, notably those engaging in intensive agricultural practices, bees face progressively challenging living conditions [
10,
11]. In these regions, a decline in honeybee colonies has been observed for approximately two decades, necessitating continuous reproduction to maintain the population levels of the colonies at a sustainable level [
6,
12,
13,
14]. Intensively farmed appear to be increasingly less hostile to honeybees due to large-scale monocultures of crops that do not provide benefit to bees [
14,
15,
16,
17], as well as a significant increase in the amount of insecticides used in agriculture for crop protection [
14]. The occurrence of acute pesticide poisoning clearly demonstrates the negative impact of agriculture on bees. Research has demonstrated that bees exhibit resistance to low to moderate levels of pesticides present in their food [
18,
19]. However, it remains challenging to discern sublethal effects in bees, particularly given that the
Varroa destructor mites are the primary determinant of bee health and longevity [
19].
The high densities of bee colonies in some European countries, as well as the threats from the chemicalization of agriculture, mean that an increasing number of bee colonies are being kept in urban areas, where conditions are conducive to the survival of pollinators [
20,
21]. Urban areas are characterised by a high diversity of nectar- and pollen-bearing plants, which bloom for extended periods, thereby providing food sources for insects. They have a warmer climate zone, which has been demonstrated to be conducive to the production of large quantities of nectar. An example of this phenomenon in Poland is
Robinia pseudoacacia L., whose flowers frequently undergo frost damage or fail to produce nectar. In the villages, aside from the cultivation of nectariferous crops, there is an absence of initiative to expand the forage base. Conversely, there has been a decline in bee forage, particularly those that start later in the season. Old roadside lime trees are being removed, and in forestry settings,
Robinia is being felled. Foresters regard this species as an invasive tree that is not conducive to effective forest management. The population of heather shrubs is declining, and these plants are reaching old age and producing low yields of nectar. This is largely due to restrictions on shrub regeneration management, such as through burning and cutting.
The process of chemisation in agriculture has resulted in the potential risks associated with beekeeping in certain areas. In urban areas, there is either a complete prohibition on the use of pesticides or a significantly reduced scale of their application when compared with rural areas. In addition to their conventional commercial applications, urban apiaries also possess significant educational value. Demonstration apiaries are organised to welcome children and young people to classes and workshops, allowing them to get to know the bees, learn about their lives, and taste the products. In Poland, urban apiaries are subject to the same regulations as other apiaries. These apiaries are typically either private enterprises or funded by local governments or companies as part of a broader marketing strategy.
The issue of whether the development of civilisation, urbanisation, and industrialisation are having a detrimental effect on pollinating insects is a matter of considerable debate. Research to date has focused predominantly on solitary bees. A comparison has been made of bee species richness according to the degree of urbanisation. In urban environments, there is a high abundance of bee species, but social species are more numerous, including polylectic species [
22]. The honeybee is an eminently polylectic species, raising concerns that it may dominate these areas, displacing other insects (
Apoidea) [
23,
24]. The adaptability of social insects has been identified as a key factor in their increasing dominance in urban environments. It is notable that the honeybee is well-suited to urban environments, exhibiting adaptability through its ability to fly over long distances and forage for food with greater efficiency compared to other wild bees [
25]. Several studies have reported that the honeybee poses a threat to wild bees [
26,
27,
28,
29,
30]. Conversely, other studies showed either no impact or inconclusive results [
31].
The most famous urban apiary is the one founded in 1980 in Paris, located on the roof of the Opera Garnier. This apiary gave rise to the urban beekeeping industry, and today, there are more than 3000 bee colonies. Paris bee colonies produce an average of 50 kg of honey per year, with peaks of up to 80 kg in seasons where forage is particularly abundant. In contrast, in the rural environment colonies typically yield 30 kg [
32]. In the United States, urban beekeeping was not permitted until the 1950s, when regulations began to liberalise. Bans on urban beekeeping are usually motivated by concerns for public safety [
33]. A notable increase in the number of urban beekeepers was observed in the US around 2008, likely driven by media attention attracted by reports of ‘colony collapse disorder (CCD)’ that started in 2006 [
34]. As a result, New York City and Los Angeles officially removed restrictions on urban beekeeping in 2010, establishing a significant precedent for cities and municipalities nationwide. This progressive shift was subsequently adopted by Milwaukee, Wisconsin, and Washington, DC, in 2012. The movement toward more supportive regulations continued with Boston in 2013 and Los Angeles again in 2015.
Bee densities in large cities are higher than in agricultural landscapes. For instance, the average colony density in Paris per km
2 is 6.5; colonies that exceed that of the rest of the country are −2.5; in London, it is −10; and in Brussels, it is −15 [
35]. Urban beekeeping has also become an increasingly prevalent practice in other European countries, including Germany and Poland. Most of these colonies are managed, whereas feral colonies are comparatively rare. The average wild colony density in Europe is 0.26/km
2, which is 25 times lower than in Latin America and Africa [
36].
The aim of this study is to compare the rate of development and performance of bees kept in urban and agricultural environments across various regions in Poland.
4. Discussion
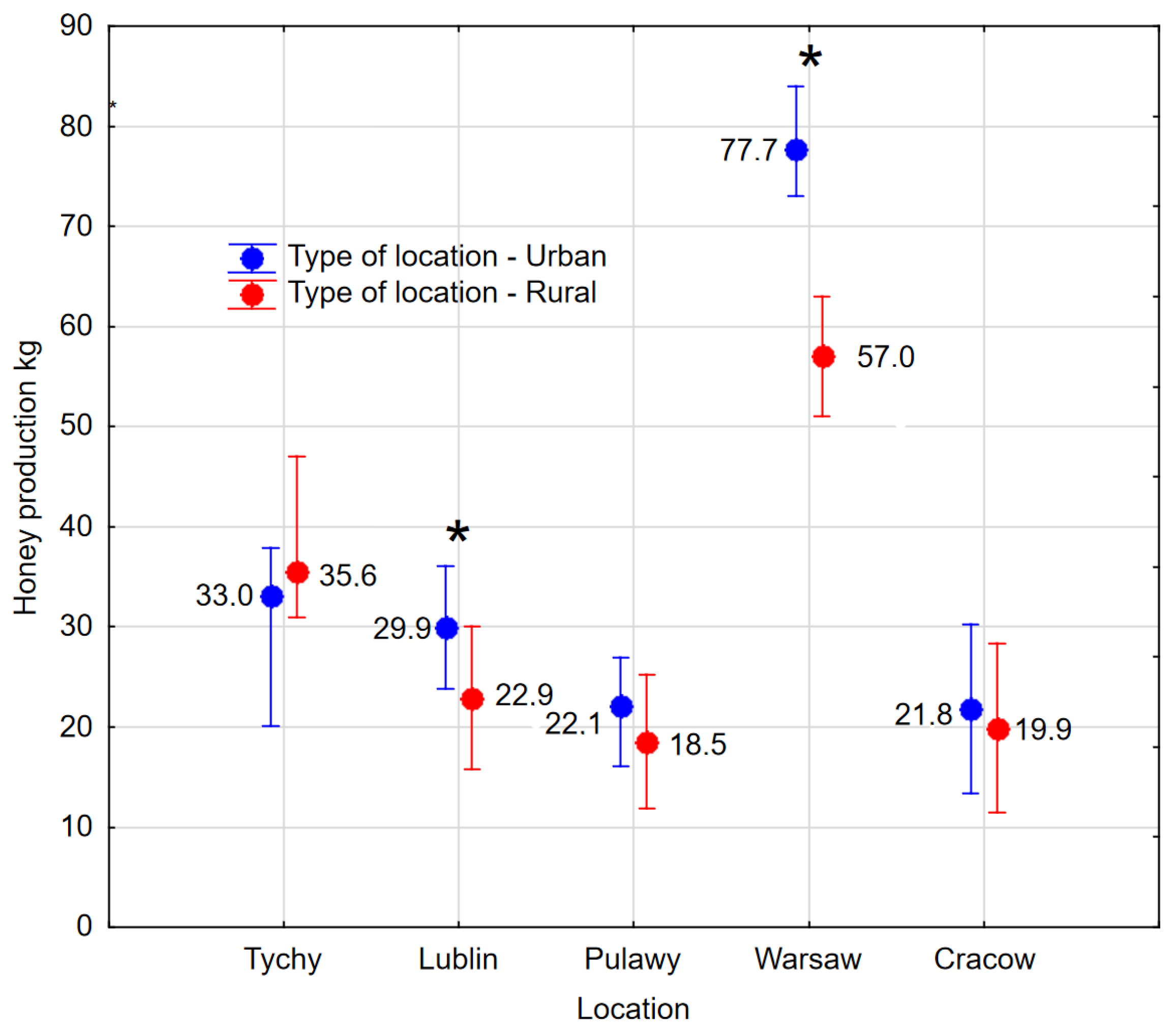
A study was conducted to ascertain the distinguishing characteristics of apiaries located in urban and agricultural areas. It was found that the distinguishing parameter is the number of broods in spring BC1, brood growth rate BGD, and bee infestation with
Varroa mites VInf. The results demonstrated that urban apiaries exhibited higher values for these parameters (
Table 2). This suggests that in urban environments, at the beginning of the season, bee colonies experience accelerated development. This finding has been corroborated in other studies [
18,
20,
21,
40,
41,
42]. In four out of five regions, higher honey yields were reported in urban areas, although this was statistically proven in only two locations. These outcomes have been corroborated by other research studies (see references [
21,
43,
44]. In the Tychy region, honey production was dominated by rural apiaries due to favourable foraging conditions. However, it should be noted that in this study, data were collected over only one season. In addition, oilseed rape was grown in that area. That contributed to a good harvest because the area received the most rainfall, and nectar was abundant. A lack of rainfall is less likely to reduce the nectar production of trees and shrubs because of the root system. This is why the yield is higher in urban areas, where the most important honeys do not come from annual plants.
In Poland, the honey production season is short, namely 3–4 months, and the most important parameter influencing the use of beneficial insects is the intensity of spring development. The number of bees in colonies in summer was slightly lower in rural apiaries.
Intensive colony development in urban apiaries led to higher levels of
Varroa mite infestation during the summer months. The spread of the parasite is favoured by colony development, as has been described in other studies [
45,
46,
47,
48]. However, infestation levels in urban apiaries have not exceeded colony-threatening values of 3/10 g bees [
49,
50], although some studies already consider 1/10 g bees as a critical level [
51,
52]. In urban areas, infestation levels were found to be higher, though not to a sufficient extent to necessitate a modification in the control strategy. However, under certain conditions, the critical level of infestation may be exceeded, particularly when the onset of spring occurs earlier than is customary. In such cases, it is recommended that bee infestation be monitored and that summer treatment be applied. Given the temporal proximity to harvest, biotechnical methods are deemed most suitable or in combination with treatments that have a short withdrawal period, such as organic acids.
Several factors have been identified as potentially contributing to low honey yields, with adverse effects on the condition of bees. In rural areas, where agricultural intensification can lead to a reduction in biodiversity, bees often face a lack of sufficient food resources, especially during periods of drought. In addition, the presence of pesticides, herbicides, and other chemicals used in agriculture can significantly reduce the ability of bees to collect nectar and produce honey. Consequently, in such instances, the quality of the soil and the diversity of the vegetation can be constrained, resulting in diminished floral availability and reduced yields. The contrasting factors observed in both urban and rural areas underscore the notion that honey yields are influenced not only by the environmental type but also by the specific characteristics of the immediate environment.
However, the 14 developmental, biological, and productivity parameters of the bee colonies were found to be most dependent on the different geographical parameters. The existence of geographical variation in living conditions for bees is not a novel finding, as evidenced by other publications [
53,
54,
55,
56]. In most cases, 12 parameters showed significant variation between geographical locations (
Table 3). The apiary in Warsaw demonstrated the highest honey yield; this was a noteworthy achievement that outperformed both other regions and the rural control apiary. The average yield in Warsaw was 20.7 kg higher than in the rural apiary, a discrepancy that can be explained by the urban heat island effect, which is a term used to describe the phenomenon by which urban areas are found to be warmer than the surrounding countryside. It has been demonstrated that species which are more demanding in relation to temperature produce more nectar than those which inhabit areas outside the city, where other weather parameters are similar, e.g., insolation or amount of rainfall. The apiary in Warsaw exhibited the lowest overwintering strength while simultaneously achieving the highest honey yield among all the locations studied. This phenomenon may be attributed to the excessive workload placed on the bees. The practical advice to be derived from this experience is that, in areas where temperatures are higher and bees develop more rapidly during the spring months, it is possible to overwinter them in a state of reduced strength and yet still expect them to demonstrate high levels of productivity.
The urban apiary in Lublin demonstrated exceptional performance in the majority of the 14 parameters under consideration. It was characterised by notably dynamic colony development during the spring period and the presence of stronger colonies throughout the season. This phenomenon may be attributed to the specific location of the apiary, which was the sole facility situated on the roof of a five-storey building. This particular location provides effective protection from wind while also ensuring optimal sun exposure. The distinctiveness of this region is reflected in
Figure S5.
The climatic conditions at the macro- and micro-scale and forage conditions in which the experimental colonies were tested had a very significant effect on the parameters of the bee colonies. The existence of a factorial interaction was confirmed by means of a principal component analysis (PCA).
Figure 2 and
Figures S1–S3 demonstrate additionally that an interaction of factors of location and year of testing occurs.
In the temperate climate zones, seasonal variation in bee traits plays a crucial role. This assertion is corroborated by studies [
25,
57,
58]. This finding is consistent with the results of the present study, which demonstrated that climatic conditions in each year significantly influenced bee overwintering B1, OIndex, spring colony development BC0, BC1, and honey yield. In 2022, the values of the key traits of colony development were higher than in 2021. These fluctuations can be attributed to various bee responses to weather conditions, such as the number of flightless days when bees are unable to forage, as well as the presence of forage plants that do not produce nectar during cold weather, such as
R.
pseudoacacia, which flowers briefly [
57].
The environmental impact on bees was so significant in the experiment that the correlation between brood and bee numbers and honey production was low at 0.2 and 0.26, respectively. Much higher correlation coefficients were obtained by other researchers, but they kept their apiaries in one place. The correlation coefficients for brood and honey production were 0.73 [
59], 0.55–0.62 [
60], and 0.96 [
61]. However, the correlation coefficients between the number of bees and honey production were 0.71 [
59], 0.54–0.64 [
60], and 0.96 [
61]. In the context of urban apiculture, the conservation of wild pollinating insects should be a paramount consideration. While the majority of studies do not corroborate the competitive impact of honey bees on other
Apoidea, green area planners must be cognizant of the fact that certain plant species are less attractive to honey bees but are favoured by certain bee species.






{kind=link}
{kind=link}
{kind=link}
{kind=link}