1. Introduction
Elite youth soccer players must meet substantial physical demands. In a typical game, adolescent players cover ~8–9 km, with roughly 20–22% of that distance at high speed (13–18 km/h) and frequent sprints [
1]. To sustain such intensity, both aerobic and anaerobic energy systems are taxed continuously throughout the match. Consequently, comprehensive conditioning is required to develop endurance, repeated-sprint ability, speed, and power in concert. High-intensity interval training (HIIT) has become a cornerstone of soccer conditioning for this reason. As Buchheit and Laursen (2013) explain, HIIT (or HIT) comprises repeated bouts of high-intensity exercise (from brief all-out sprints of <45 s to longer 2–4 min intervals) interspersed with recovery [
2]. An optimal HIIT session is designed to accumulate several minutes above ~90% of VO
2max (“red zone”) per session, since this high-intensity stimulus elicits both central (cardiac output) and peripheral (muscle oxidative) adaptations [
3].
High-level youth soccer demands the development of a broad range of skills and fitness qualities, often with limited training time and busy schedules. Empirical data support its effectiveness in adolescents: compared to alternative training, HIIT interventions typically produce larger gains in aerobic fitness (e.g., mean VO
2max improvements of ≈7% vs. ≈4%), even though the total training volume may be lower [
4]. Notably, studies on junior players have found that small-sided game drills (sport-specific intervals) and generic running HIIT elicit comparable improvements in aerobic capacity [
5], suggesting that the HIIT principle (high intensity with recovery) can be delivered in multiple forms. Recent trials also report that even short-term, progressively overloaded HIIT programs can enhance the change-of-direction speed, the repeated-sprint ability, and the intermittent shuttle-run capacity of U-15 players [
6].
Despite this progress, most training studies examine outcomes one metric at a time (e.g., VO
2max, sprint time, lactate). In reality, adaptation is integrative: improvements in cardiovascular, metabolic, neuromuscular, and motor domains occur simultaneously and interact. Recent advances in exercise physiology propose that performance should be understood through a systems-level lens, emphasizing coordination across metabolic, cardiovascular, neuromuscular, and cognitive processes. Emerging frameworks like network physiology highlight that physiological states arise from nested interactions across systems [
7,
8]. In line with this, we sought to assess not only univariate improvements but also how key performance variables interact and reorganize under HIIT conditions.
To capture multidimensional adaptation, this study evaluated a broad set of physiological and cognitive parameters previously shown to respond to high-intensity interval training (HIIT). HIIT has been reported to reduce blood lactate accumulation and elevate lactate threshold speed, reflecting improved glycolytic efficiency and buffering capacity [
9,
10]. Related markers such as base excess and pH recovery also improve post-HIIT, indicating enhanced acid–base regulation, though evidence in youth populations remains limited [
3,
11]. Heart rate recovery and kinetics are often positively affected and are linked to improved autonomic control [
12]. While speed reserve and anaerobic work capacity (D′) have not been widely studied in HIIT contexts, they are expected to improve due to enhanced neuromuscular conditioning [
13,
14]. Emerging evidence suggests that HIIT may improve cognitive outcomes, including reduced perceived exertion and enhanced executive functioning. Specifically, it may benefit inhibitory control, cognitive flexibility, working memory, and processing speed—key skills for decision-making and tactical responses in soccer. These effects may be driven by improved neural efficiency and increased neurotrophic signaling, such as BDNF release, supporting both short- and long-term cognitive adaptation [
15,
16]. Notably, HIIT has been associated with increased levels of brain-derived neurotrophic factor (BDNF), a biomarker tied to neuroplasticity and mental resilience, yet its specific role in systemic coordination and fatigue remains unclear. Brain-derived neurotrophic factor (BDNF), a neurotrophin involved in synaptic plasticity, learning, and memory, has also been linked to exercise-induced adaptations in both cognitive performance and metabolic regulation, making it a candidate biomarker for multidomain responsiveness to high-intensity training [
17,
18]. To address these gaps, we adopted an inter-domain correlation structural approach, hypothesizing that a four-week HIIT program will improve not only individual performance metrics but also coherence across physiological systems. Through correlation mapping and cluster analysis, we aimed to identify patterns of integrated adaptation that may serve as sensitive markers of training response, with implications for individualized conditioning and the early detection of maladaptive trends.
In the present study, we applied a novel cross-domain statistical associations framework to elite youth soccer players undergoing HIIT intervention. By examining how the “interaction patterns” among multiple performance and physiological metrics change pre- to post-training, we aimed to identify network signatures of adaptation that go beyond individual test improvements. This multidomain perspective aligns with current calls for integrative exercise physiology and should provide deeper insight into how high-intensity training reshapes the athlete as a whole.
2. Materials and Methods
Sampling Protocol and Schedule: This randomized controlled trial was conducted across five distinct phases from February to June 2024, including baseline testing, a four-week intervention, post-intervention evaluation, standard training continuation, and final assessments. Ethical approval for the study was granted by the Commission for Research Ethics at the University of Physical Education in Wrocław. Written informed consent was obtained from all participants and their legal guardians prior to enrollment, in accordance with the Declaration of Helsinki and institutional guidelines. This study focused on multidomain adaptations in elite youth soccer players and employed a comprehensive array of physiological, cognitive, and biochemical assessments. Participants were randomly assigned to either a HIIT intervention group (n = 12) or a control group (n = 11). A total of 23 elite male soccer players (HIIT: n = 12; control: n = 11), aged 17 to 20 years (mean ± SD: 19.3 ± 0.7 years), completed physiological and cognitive testing across three phases (R1, R2, R3). A G*Power 3.1 analysis was conducted to estimate the required sample size for detecting a medium group × time interaction effect (f = 0.25) in a 2 × 3 mixed-design ANOVA, assuming α = 0.05, power = 0.80, correlation among repeated measures = 0.5, and ε = 0.75. This yielded a required total sample of 28–36 participants. Our sample (n = 23) was slightly below this threshold but remained adequately powered to detect large effects, such as those observed in the Anaerobic Fatigue Index (η2 = 0.21, p = 0.035). Goalkeepers were excluded from the analysis due to the distinct physiological and tactical demands of their position; thus, only outfield players were included. The semi-professional participants followed a structured microcycle consisting of five on-field training sessions, two gym-based strength sessions (conducted on Tuesdays and Fridays), and one competitive match held on Saturdays.
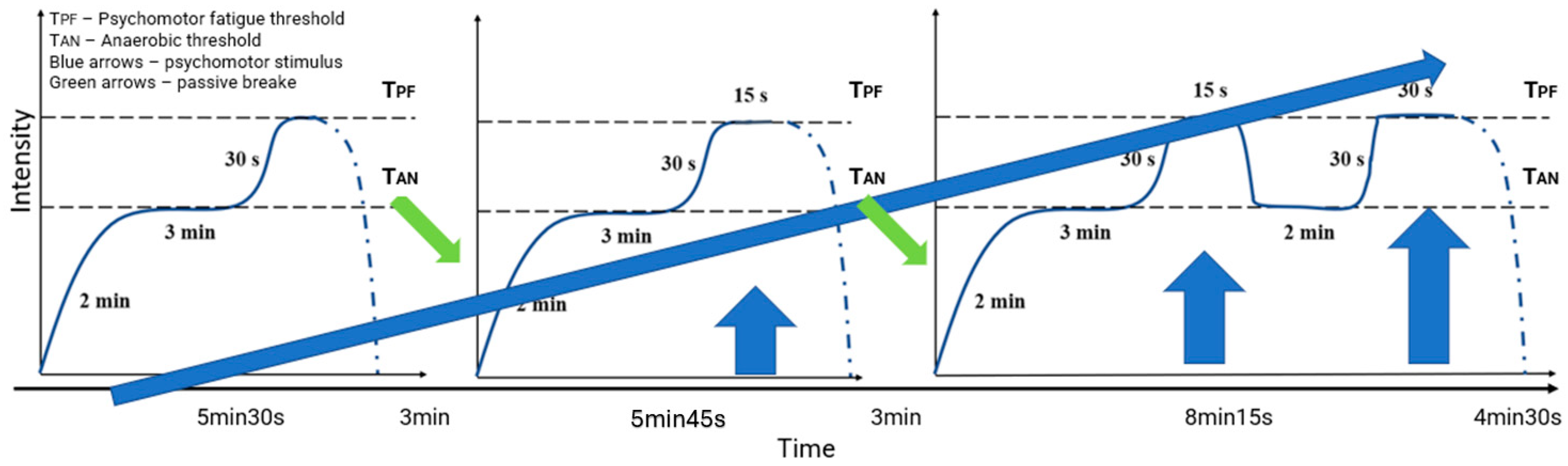
This study comprised three laboratory testing sessions (pre-, post-, and final), along with ongoing field-based and wellness monitoring. Daily subjective wellness questionnaires tracked sleep quality, fatigue, muscle soreness, mood, and perceived exertion. Athletes were instructed to maintain regular hydration habits, and all laboratory testing sessions were conducted in a euhydrated state,. Recovery was standardized by requiring 48 h without intense physical activity before each testing session. During the four-week intervention, the HIIT group performed twice weekly 30 min motor intermissions consisting of three specific long intervals during which the athletes ran at an intensity between their anaerobic threshold and the psychomotor fatigue threshold determined in the first laboratory session (
Figure 1). The anaerobic threshold (ANT) was identified using fingertip blood lactate sampling and is defined as the exercise intensity corresponding to the individual anaerobic threshold, determined using lactate curve analysis following the protocol by Konefał et al. (2022) [
9]. The psychomotor fatigue threshold (PFT) was determined using choice reaction time (CRT) assessments performed in the last minute of each 3 min treadmill stage. CRT deterioration, as described by Chmura et al. (2010) [
15], was operationally defined as the first intensity at which a statistically significant increase in response latency or the error rate was observed compared to previous stages. The control group followed standard preseason training with a similar session frequency but without high-intensity interval exposure.
Training Load Monitoring: Training loads were continuously monitored using Catapult Vector (Melberne, Australia) GPS systems and heart rate telemetry, capturing internal and external workload indices such as total distance, heart rate exertion, and player load. Each player’s session data were phase-coded (R1–R3), enabling within- and between-group comparisons [
2,
3].
Physiological and Cardiometabolic Measurements: Oxygen uptake (VO
2) was continuously monitored during laboratory-based incremental exercise testing using the Cortex MetaMax 3B (Leipzig, Germany) mobile ergospirometer, a reliable device for breath-by-breath gas exchange analysis. Heart rate (HR) was recorded with the Polar RS400 (Kempele, Finland) heart rate monitor, which provided beat-by-beat data throughout all laboratory and field tests. The anaerobic threshold (ANT) was determined through fingertip blood lactate analysis after each exercise load, with lactate samples collected using standard capillary procedures and interpreted using the method described by Konefał et al. (2022) [
9]. The progressive treadmill protocol began at 8 km/h and increased by 2 km/h every 3 min until volitional exhaustion, following a fixed incremental load design [
11].
BDNF and Blood Biochemical Analysis: Venous blood samples were collected to assess brain-derived neurotrophic factor (BDNF) concentrations, both before and after the treadmill tests, and additionally after the 14 km/h load. BDNF analysis was performed using standard ELISA protocols. Additional venous blood draws captured the complete blood count and platelet levels, while fingertip samples tracked dynamic lactate fluctuations across each stage. Post-test blood sampling occurred 10 min after the final load to assess recovery-phase BDNF response [
18,
19].
Cognitive and Psychomotor Testing: Cognitive and neuromotor fatigue were quantified through choice reaction time (CRT) testing using the APR Reaction Meter (UNI–PAR, Poland). CRT tasks were administered during the final minute of each 3 min treadmill stage and involved responding to 25 audiovisual stimuli, thereby allowing precise identification of the psychomotor fatigue threshold (PFT) [
14,
15]. A familiarization phase ensured procedural consistency and minimized learning effects.
Field-Based Endurance Testing: The on-field performance capacity was assessed via a Critical Power Test (CPT) conducted on a 200 m oval track. Athletes completed two-time trials (2600 m and 1000 m), performed in sequence on the same day with 30 min of passive recovery in between. The tests were preceded by a standardized 20 min warm-up. Running times were measured using electronic timing, and the critical speed (CS) and D′ were calculated using the linear distance–time model described by Galbraith et al. (2011) [
20].
2.1. Derived Physiological and Performance Metrics
Several derived variables were computed to capture multidomain physiological and cognitive performance adaptations. For cardiorespiratory performance, we calculated VO2/kg (VO2max normalized to body mass) and O2 Pulse (oxygen consumption per heartbeat), which serve as markers of aerobic capacity and stroke volume, respectively. Heart rate dynamics were characterized using HR recovery, HR reserve, HR threshold speed, and HR kinetics slope. Ventilatory efficiency was assessed via VE/VO2 and VE/VCO2 ratios.
Blood-based metrics included lactate peak, lactate threshold speed, lactate rise slope, and the lactate accumulation index (quantifying lactate sensitivity to speed), alongside acid–base markers such as ΔHCO3std and ΔBE. Hematological and immune response variables included ΔHGB, ΔWBC, PLRrest, and PLRafter.
Subjective exertion and fatigue were quantified via the RPE–Speed Ratio, the perceived exertion slope, and the Anaerobic Fatigue Index. Cognitive and neuroplasticity-related metrics included ΔBDNF, BDNF–Cognition, the cognitive load index, cognitive load accumulation, and cognitive–physical divergence. Neuromuscular integrity under fatigue was captured using ΔCRR and the Psychomotor Score at the Final Stage.
All formulas, computational definitions, and domain classifications for these derived metrics are detailed in
Supplementary Table S1.
2.2. Data Analysis—Variable Classification
To quantify multidomain physiological and cognitive adaptation to high-intensity interval training (HIIT), a series of advanced metrics were computed from raw performance and laboratory data using a standardized Python (v3.10) pipeline. These derived variables spanned four domains: metabolic, neuromuscular, physical, and cognitive (refer to
Supplementary Table S1). Unless otherwise stated, measurements were recorded across incremental treadmill running stages (8–20 km/h), and delta values represent post-intervention minus baseline differences. Slope metrics were derived using first-order polynomial fitting.
To facilitate multidomain analysis, all outcome measures were categorized into four a priori classifications based on physiological relevance: metabolic, physical, neuromuscular, and cognitive. The classification scheme is detailed.
2.3. Statistical Analysis
A 2 × 3 mixed factorial design was employed, incorporating one between-subjects factor—group (high-intensity interval training [HIIT] vs. control)—and one within-subjects factor—the test phase (R1–R3). For each physiological and cognitive metric, a mixed-design ANOVA was conducted to examine the main effects of the group and the phase, as well as their interaction. To investigate temporal dynamics within each group independently, a repeated-measures ANOVA was performed separately for the HIIT and control groups. Independent samples t-tests were additionally applied at each phase (R1–R3) to compare group-level differences.
To evaluate test–retest reliability, the derived metrics were analyzed using Pearson’s correlation-based intraclass correlation coefficients (ICCs), reflecting the stability of each metric over time. Concurrently, measurement variability was quantified via the coefficient of variation (CV%), and the metrics were subsequently classified according to standard interpretive thresholds: the ICC was labeled as excellent (≥0.90), good (0.75–0.89), moderate (0.50–0.74), or poor (<0.50); the CV was categorized as very low (<5%), low (5–9.9%), moderate (10–19.9%), or high (≥20%). A combined reliability tier designation (high, acceptable, borderline, unreliable, or mixed/needs review) was applied to synthesize the ICC and CV outcomes. Greenhouse–Geisser corrections were implemented where the assumption of sphericity was violated. Effect sizes were reported using partial eta squared (η2), and the statistical significance was set at p < 0.05.
To examine cross-domain associations, Pearson correlation coefficients were computed between all metric pairs within each group (HIIT, control) and test phase (R1–R3). Comparisons were performed both across time (R1 vs. R3 within group) and between groups at each phase (HIIT vs. control at R3). Fisher’s r-to-z transformation was applied to determine the statistical significance of correlation changes across conditions. For each pairwise comparison, the z-statistic was computed, and the corresponding
p-value was derived based on the standard normal distribution [
7,
8,
21].
Statistically significant shifts in correlation (p < 0.05) and absolute delta values (|Δr| > 0.5) were retained for further visualization. To focus on cross-domain dynamics, only interactions between variables of different classifications (e.g., cognitive↔metabolic) were considered for heatmap generation.
Heatmaps were created using Seaborn (v0.12) in Python (v3.10), with the color intensity representing Δr values and tick labels color-coded by variable classification. Legends indicating type–pair interactions were constructed to guide visual interpretation. All results, including z-statistics, correlation deltas, and significance levels, were exported to Excel (XLSX) files for reproducibility. Analysis code and output visualizations are available upon request.
3. Results
3.1. Validation of Training Load
A repeated-measures ANOVA was performed to evaluate load metrics across time (R1 to R3) and between groups (HIIT vs. control). No significant group × time interactions were observed for the total weekly training load, cumulative distance, or sRPE (all p > 0.1), indicating comparable overall workload volumes across groups. However, PlayerLoad per minute—an intensity-sensitive variable—was significantly higher in the HIIT group (p = 0.02), with a moderate-to-large effect size (Cohen’s d = 0.77).
At baseline (R1), no significant differences were observed between the HIIT and control groups in any measured variable, confirming that the random allocation effectively created two statistically comparable cohorts. Metrics such as VO2max, HR recovery, BDNF levels, the blood lactate concentration, cognitive performance, and perceived exertion did not differ (p > 0.05), establishing a consistent starting point for evaluating training effects.
3.2. Reliability of Derived Metrics
The reliability analysis of the derived physiological and neurocognitive metrics revealed a wide spectrum of measurement consistency, as assessed by test–retest intraclass correlation coefficients (ICCs, Pearson’s r) and the coefficient of variation (CV%). Among the 36 metrics evaluated, three demonstrated excellent reliability, including HR
last (ICC = 0.915, CV = 2.00%) and Cognitive–Physical Divergence (ICC = 0.915, CV = −2.00%), both of which were classified as high reliability due to very low variability and strong reproducibility (refer to
Supplementary Table S2). The Anaerobic Fatigue Index also showed a good ICC (0.786) and a very low CV (2.8%), qualifying as acceptable. A moderate number of metrics, such as O
2 Pulse, the lactate threshold speed, VO
2/kg, and the VE/VCO
2 Ratio, fell into the borderline or mixed/needs review categories due to moderate ICCs and CVs, indicating fair but inconsistent reproducibility. In contrast, several inflammatory (e.g., PLR
rest, ΔWBC), acid–base (e.g., ΔBE, ΔHCO
3std), and hematological markers (e.g., ΔHGB), alongside certain metabolic indices (e.g., lactate peak, the lactate accumulation index), demonstrated poor ICCs combined with high CVs, and were thus classified as Unreliable. These findings suggest that while some derived variables offer robust reproducibility for use in longitudinal or interventional studies, others should be interpreted with caution or further refined to improve their measurement stability.
3.3. Training-Induced Changes over Time
In the control group (standard training), several fitness parameters declined significantly across the 4-week period, consistent with a detraining or insufficient stimulus effect (notable given the elite status of players). Most concerning, the control group’s relative VO2max dropped by ~11% from baseline (from ~56.9 to 50.6 mL·kg−1·min−1), a significant decline (F(2.16) = 5.47, p = 0.0176, η2 ≈ 0.27). The absolute VO2max also showed a downward trend (−6%, p = 0.064). Markers of anaerobic endurance deteriorated: the control group exhibited a significant increase in lactate accumulation (i.e., worse lactate handling), with the lactate accumulation index rising (time effect F ≈ 15.3, p = 0.0003, η2 ≈ 0.45) and the lactate rise slope steepening (F ≈ 8.96, p = 0.0031), indicating that at a given work rate they accumulated lactate faster by the end of the study. Consistently, the control’s peak blood [La] tended to increase (mean +0.8 mmol/L, p > 0.2) and the velocity at the OBLA (onset of blood lactate ~4 mmol) improved only modestly (p = 0.074, n.s.), despite normal soccer training. Maximal ventilation in the control group dropped significantly (F(2.18) = 3.89, p = 0.045), as did VCO2max (−14% by Week 4), suggesting a reduction in peak exercise capacity. Unlike the HIIT group, the control showed a blunted BDNF response to exercise over time: the BDNF measured during submaximal exertion (e.g., running at 14 km·h−1) fell markedly in the control group (F(2.22) = 13.33, p = 0.0009), whereas it was maintained in HIIT. There were also slight impairments in heart rate recovery in the control group: the post-exercise HR drop (1 min recovery) became smaller (from ~27 to 25 bpm, F(2.18) = 3.47, p = 0.0508), although the resting HR and HRreserve remained unchanged. On the positive side, the control group did exhibit a small improvement in lactate threshold speed (+~1.2 km·h−1 on average) and maintained critical speed gains similar to HIIT (CS p = 0.020). However, these improvements did not reach statistical significance in the control group (e.g., lactate threshold p = 0.074), and were accompanied by declines in other domains. No significant within-group changes were seen in the control’s cognitive performance scores or cognitive load metrics, though there was a non-significant trend of worsening cognitive-load tolerance mid-study.
The HIIT group demonstrated significant improvements in several performance metrics over time. Notably, the critical running speed (CS) increased significantly (e.g., CS in m·s−1: F(2.14) = 6.58, p = 0.0096, η2 ≈ 0.41; CS in km·h−1: F(2.14) = 6.60, p = 0.0096) in the HIIT group, indicating enhanced high-intensity running capacity. By contrast, the associated anaerobic capacity d′ (running distance above critical speed) showed no significant change (p > 0.5). Aerobic metabolic performance was largely maintained in the HIIT group: neither the absolute VO2max (F(2.18) = 1.23, p = 0.320) nor the relative VO2max (mL·kg−1·min−1) changed significantly over the 4 weeks (p = 0.999)—reflecting that the intervention group sustained their aerobic fitness level while the control declined (see below). The maximal VE (ventilation) and VCO2 showed no significant time effect in HIIT (p > 0.9), and HRmax remained unchanged as expected (no training effect on maximal heart rate). In terms of lactate dynamics, the HIIT group did not show significant changes in lactate threshold speed or peak blood lactate (both p > 0.2). Interestingly, resting and exercise BDNF levels in the HIIT group were generally stable: resting BDNF showed no clear increase (one BDNF measure trended lower, p = 0.12, while another context showed a time effect, p = 0.007, albeit reflecting a slight decline in concentration). Cognitive and psychomotor performance metrics within the HIIT group did not change significantly over time. For example, the reaction/processing speed at rest and under fatigue (e.g., CRT/psychomotor tests) remained statistically unchanged (p > 0.1). The cognitive load index (quantifying cognitive performance under physical load) showed no significant within-group change in HIIT (p = 0.188), nor did the Cognitive–Physical Divergence score (p = 0.919), suggesting these novel cognitive metrics were stable over the short intervention.
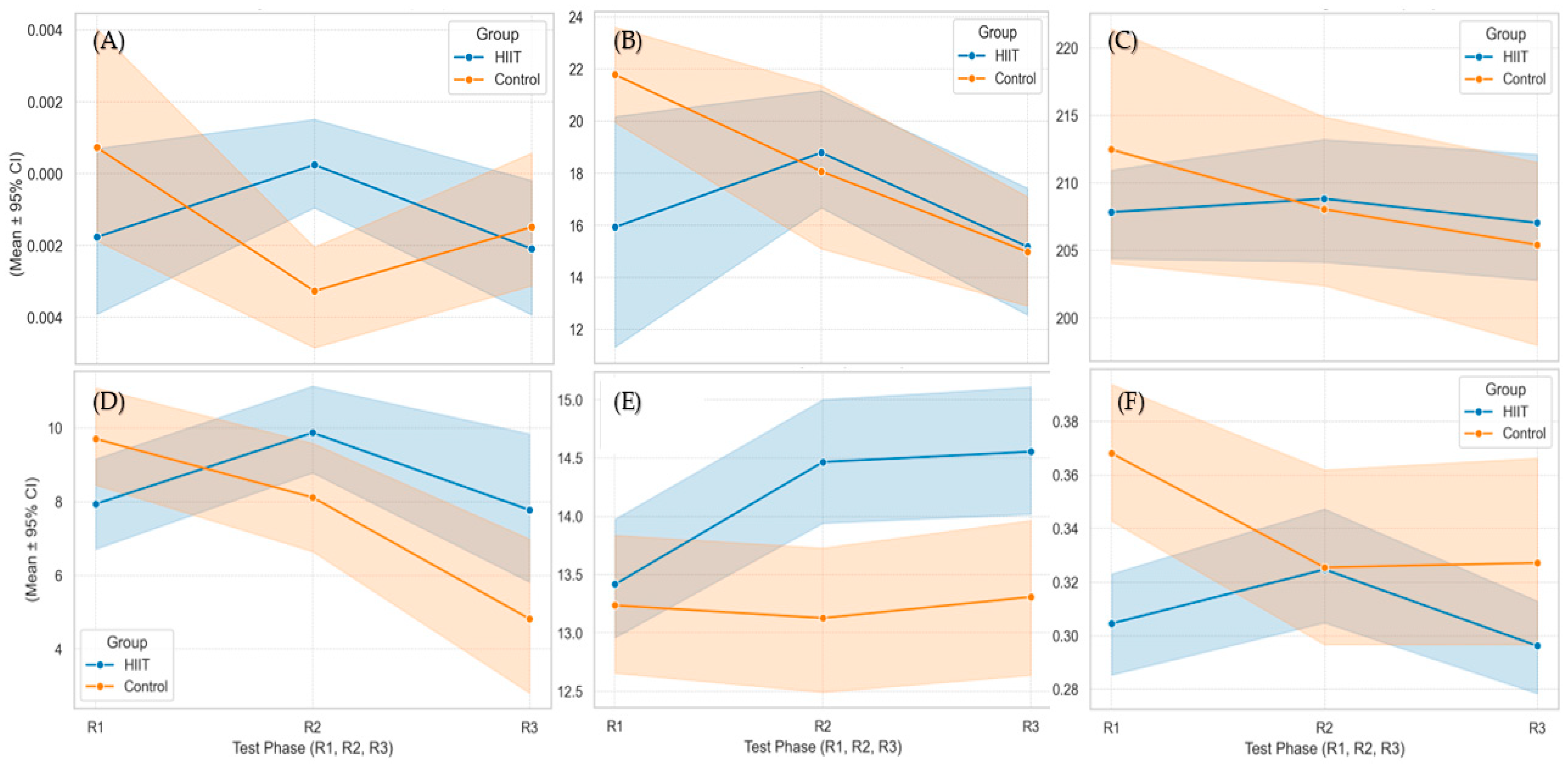
3.4. Group × Time Interaction Effects
A two-way mixed ANOVA (group × time) was conducted to examine whether the trajectory of physiological and cognitive outcomes differed between the HIIT and control groups across testing phases. Significant interaction effects were observed for several metrics, indicating differential adaptations over time between groups (
Figure 2) (
Table 1).
A statistically significant interaction was found for OBLA (onset of blood lactate accumulation) velocity (F(2.22) = 6.35, p = 0.0040, η2 = 0.24). The HIIT group showed a greater increase in OBLA speed (from ~14.2 to 14.5 km·h−1) compared to the control group (from ~12.8 to 13.3 km·h−1). Similarly, both the lactate accumulation index (F(2.22) = 5.58, p = 0.0084, η2 ≈ 0.26) and the lactate rise slope (F(2.22) = 6.71, p = 0.0037, η2 ≈ 0.30) demonstrated significant interactions.
A large interaction effect was also found for exercise-induced BDNF levels (F(2.30) = 6.28, p = 0.0053, η2 ≈ 0.30), with divergent trajectories between groups. The cognitive load index (F(2.34) = 5.67, p = 0.0075, η2 = 0.25) and cumulative cognitive load index (F(2.34) = 4.90, p = 0.0135, η2 = 0.22) also showed significant group × time effects. The cerebrovascular index CRR at PFT (F(2.40) = 4.86, p = 0.0129, η2 = 0.20), the final-stage psychomotor reaction time (PSYlast; F(2.34) = 3.62, p = 0.037, η2 = 0.18), post-exercise hemoglobin (F(2.32) = 5.70, p = 0.0076), the resting blood pH (F(2.32) = 4.83, p = 0.0147), standard bicarbonate change (F(2.32) = 4.04, p = 0.027), and the Anaerobic Fatigue Index (F(2.28) = 3.78, p = 0.035) all demonstrated statistically significant interactions.
No significant interactions were observed for VO2max, heart rate metrics, or baseline anthropometric measures.
3.5. Cross-Domain Relationship Shifts
In addition to univariate improvements, significant shifts in inter-variable correlations were observed in the HIIT group, suggesting enhanced physiological integration. Notably, the correlation between the lactate rise slope and the perceived exertion slope increased (∆r = +0.58,
p < 0.01), as did the association between HR recovery and ∆Base Excess (∆r = +0.54,
p < 0.01). Strong statistical associations emerged between the cognitive load index and both the speed reserve (∆r = +0.61,
p = 0.003) and the lactate threshold speed (∆r = +0.92,
p = 0.014). These changes highlight emergent coordination between metabolic, cardiovascular, and cognitive domains in the HIIT group. By contrast, the control group did not show substantial shifts in inter-variable correlations, reinforcing the idea that multidomain integration was specific to the high-intensity intervention (
Figure 3).
In the clustering analysis, we examined how physiological and cognitive variables clustered across test phases (R1–R3) in both the HIIT and control groups (
Figure 4). Statistical tests of domain distributions within clusters using the Chi-square test revealed no significant deviation from randomness in either group or phase (all
p > 0.05). However, the control group in phase R1 showed a trend toward significance (χ
2 = 17.28,
p = 0.068, df = 10), suggesting early, non-random clustering of domains under passive conditions. In contrast, the HIIT group remained more uniformly distributed across all phases (R1: χ
2 = 11.61,
p = 0.312; R3: χ
2 = 11.51,
p = 0.174).
Cluster stability analysis, quantified as the number of transitions a variable made between clusters (range: 0 to 2), showed that variables from the cognitive domain in the control group remained entirely stable (mean = 0.00, SD = 0.00, n = 11), indicating persistent clustering patterns. Similarly, physical (mean = 0.00, n = 3) and metabolic (mean = 0.00, n = 1) variables in the HIIT group exhibited complete cluster stability. On the other hand, cardiovascular variables in the control group showed maximum instability (mean = 2.00, SD = 0.00, n = 4), indicating that without training intervention, these variables shifted clusters across all phases.
Transition Matrix Analysis further emphasized these trends. In the HIIT group, anthropometric and cardiovascular domains demonstrated broader transitions between clusters (e.g., two transitions in 1→2 and 2→3 for the anthropometric domain; one transition each for 2→1, 3→1, and 3→2 in the cardiovascular domain). Conversely, the control group displayed more conservative transitions in cognitive–physical variables, including stable patterns like 1→1 and 2→2, suggesting inertia in hybrid domain clustering when no training adaptation occurs.
4. Discussion
The integration of multidomain correlation mapping and hierarchical clustering analyses revealed that HIIT fosters tighter coupling among metabolic, cognitive, and neuromuscular functions, reflecting a more unified systemic adaptation. These findings support the emerging paradigm in exercise physiology that emphasizes coordination among systems, rather than isolated improvements, as a marker of functional adaptation [
7,
8,
21]. Given the number of variables analyzed across physiological, cognitive, and perceptual domains, we acknowledge the increased risk of Type I errors associated with multiple comparisons—particularly in the context of a short, four-week intervention. While our use of dual-threshold filtering (
p < 0.05 and |Δr| > 0.5) helped minimize the likelihood of spurious correlations, these findings should be interpreted as exploratory and hypothesis-generating. Future studies with larger samples and longer follow-up durations are warranted to confirm the observed inter-domain patterns and account for potential cumulative testing effects.
4.1. HIIT-Induced Improvements in Derived Physiological and Cognitive Indices
The reductions observed in several physiological and cognitive metrics among control group participants over the 4-week period are noteworthy, particularly given their elite training background. The most plausible explanation is that while these athletes participated in routine soccer training, the sessions may not have provided a sufficient overload stimulus to maintain high-level adaptations. This aligns with previous findings that technical–tactical sessions, if not supplemented with targeted conditioning, may not elicit the neurometabolic demand required to preserve multidomain performance metrics. The decline likely reflects a combination of training specificity and relative detraining, rather than random measurement errors.
These findings indicate that the HIIT intervention elicited specific adaptations not observed in the control group. The improvements in OBLA speed, lactate kinetics, and buffering capacity suggest enhanced metabolic efficiency in the HIIT group, consistent with the prior literature on lactate clearance and monocarboxylate transporter adaptation following high-intensity training. The observed cognitive-related interactions, particularly in indices of cognitive load and BDNF response, suggest that HIIT may confer protection against cognitive decline under fatigue—an emerging area of interest in neuroexercise research.
The preservation of exercise-induced BDNF levels in the HIIT group, in contrast to the decline observed in the controls, supports the hypothesis that HIIT may maintain neurotrophic signaling under repeated physiological stress. Additionally, the stabilization of cognitive load metrics in the HIIT group, relative to mid-program deterioration in the control group, may reflect better neuromotor resilience and recovery capacity. The CRR and reaction time interactions further point to improved cerebrovascular and psychomotor stability in the HIIT cohort.
Notably, the lack of interaction effects for VO2max and heart rate metrics suggests similar trends in cardiorespiratory adaptations between groups, differing primarily in magnitude rather than direction. Overall, the moderate-to-large effect sizes (η2 ≈ 0.20–0.30) across several domains underscore the specificity and breadth of HIIT-induced adaptations, particularly in metabolic and cognitive domains.
The intervention produced marked improvements in key performance metrics, including VO
2max, lactate threshold speed, and critical speed, confirming prior evidence of HIIT’s efficacy in improving both aerobic and anaerobic performance [
2,
3,
10]. Improvements in buffering capacity, as evidenced by elevated base excess (ΔBE) and bicarbonate recovery, suggest enhanced metabolic resilience during high-intensity efforts. These results align with previous work demonstrating that repeated supramaximal exertion upregulates lactate clearance and acid–base homeostasis [
11,
13].
Cognitive indices such as the cognitive load accumulation index and the perceived exertion slope decreased significantly post-HIIT, implying improved psychophysiological efficiency under stress. This is consistent with studies indicating that regular high-intensity training modulates central fatigue responses and may improve executive functioning in athletes [
16,
17]. In this context, the observed increase in BDNF responses post-training may signal a neurotrophic mechanism supporting this cognitive resilience. The elevation in BDNF observed in the HIIT group may be explained by increased cerebral metabolic demand and neuromuscular activation during high-intensity efforts, which stimulate cortical and hippocampal activity—regions known to upregulate BDNF expression. This process is thought to be mediated via increased lactate turnover, catecholamine release, and neurotrophic signaling cascades involving PGC-1α and FNDC5/irisin pathways. These mechanisms align with previous studies showing acute and chronic BDNF elevations following intense aerobic and interval-based training, particularly in youth and adult populations under high cognitive–motor demand [
18,
22,
23].
4.2. Emergence of Systemic Coordination via Correlational Shifts
Beyond changes in individual metrics, HIIT significantly altered the correlational architecture among physiological domains. Notably, several cross-domain correlations strengthened exclusively in the HIIT group from R1 to R3. The emergence of a strong association between the lactate rise slope and the exertion slope (Δr = +0.58, p < 0.01) and between ΔBE and HR recovery (Δr = +0.54, p < 0.01) suggests enhanced coherence between metabolic buffering and autonomic recovery. Similarly, the strengthening of the relationship between speed reserve and the cognitive load index (Δr = +0.61, p = 0.003) illustrates a newfound coupling between physical and cognitive resilience.
These inter-domain relationships were absent or divergent in the control group, where changes appeared more fragmented or even reversed in direction. This divergence suggests that systemic integration is a specific adaptation to the training load, not merely a consequence of repeated testing. The increased alignment among variables in the HIIT group supports the view that exercise promotes emergent network-level adaptation [
19].
An important cross-domain marker in this study was the psychomotor fatigue threshold (PFT), which reflects the velocity at which cognitive performance begins to deteriorate under increasing physical load [
14,
15,
24]. As a hybrid indicator of both neuromuscular strain and cognitive resilience, the PFT plays a critical role in understanding how fatigue impacts decision-making and motor control in sport-specific conditions. In our analysis, the PFT was indirectly integrated through the derived speed reserve metric (PFT velocity minus critical speed), which emerged as significantly correlated with the cognitive load index in the HIIT group. This finding suggests that high-intensity training enhances the alignment between an athlete’s physical endurance and their capacity to maintain cognitive function under stress. Such coupling did not appear in the control group, reinforcing the hypothesis that HIIT fosters more efficient coordination between central and peripheral fatigue mechanisms.
4.3. Functional Interpretation of Hierarchical Clustering
Our exploratory hierarchical clustering across three time points (R1: baseline, R2: mid-training, R3: post-training) provides novel insight into how high-intensity interval training (HIIT) drives a systemic reorganization of physiological relationships. In the HIIT group, variables did not remain in their initial clusters; instead, they regrouped into new functional clusters over time, as visualized by Sankey diagram flows of variable membership. Many variables that clustered together at R1 had diverged by R3, joining different clusters alongside variables from other domains. By contrast, the control group exhibited a far more stable clustering structure, with most variables staying in the same clusters from R1 to R3 and only minimal re-shuffling. This stark difference indicates that the HIIT intervention—but not standard training –perturbed the underlying network of physiological metrics, prompting it to “rewire” into a new configuration [
11,
20]. While the observed shifts in cross-domain clustering were more pronounced in the HIIT group, we recognize that some pre-existing connectivity may be attributable to individual or maturational characteristics, and not exclusively to the intervention. Therefore, the interpretation of these shifts as HIIT-driven should be made cautiously, acknowledging that baseline coordination among systems could also play a role.
These changes in cluster composition provide a functional portrait of training responsiveness. Classical training outcomes in the HIIT group were evident—for instance, we observed significant improvements in aerobic capacity (as reflected in VO
2max) and high-intensity performance, consistent with previous studies. Participants also reported lower perceived exertion for equivalent efforts post-HIIT (indicating improved exercise tolerance), and neuromuscular tests showed gains in power. However, beyond these expected domain-specific improvements, the clustering analysis demonstrates that HIIT altered how these improvements relate to each other across domains. The emergence of new cross-domain clusters at R3 suggests that the physiological systems of the HIIT-trained players began to coordinate in new ways [
7,
8].
Essentially, HIIT not only improved individual capacities but also restructured the network of interactions among those capacities. This finding resonates with the concept of “network-level” adaptation: the training effect is seen not just in isolated variables, but in the architecture of their inter-relations. An increase in VO2max (the cardiovascular domain) might reduce excessive acute lactate buildup (the metabolic domain) and moderate perceived fatigue (the cognitive/psychological domain) during intense exercise, thereby tying these domains closer together than before. Indeed, our HIIT group’s final clusters often contained a mix of cardiovascular, metabolic, and perceptual variables, reflecting such integrated adaptation. No such reorganization was found in the controls; their physiological network appeared unchanged and compartmentalized, which aligns with the expectation that standard training of an equal volume but lower intensity mainly maintains the status quo. The control group’s lack of cluster changes underscores that the network plasticity observed in the HIIT group was a targeted effect of the high-intensity stimulus rather than a generic time effect.
Limitations and Future research: This investigation was conducted on a narrowly defined cohort of elite male adolescent soccer players, which imposes inherent limitations on the generalizability of the findings to broader populations, including female athletes, older individuals, or participants from non-elite or non-sporting backgrounds. While the four-week HIIT intervention elicited measurable physiological and cognitive adaptations, the relatively short duration may have constrained the full expression or long-term consolidation of network-level reorganization. Although control group athletes followed their regular training schedules, the lack of structured high-intensity sessions likely contributed to the insufficient stimulus required to sustain performance. This discrepancy in training quality—confirmed by both statistical and effect size comparisons—should be acknowledged as a limitation in isolating the intervention effect. Although recovery and hydration were monitored via daily wellness questionnaires and pre-test protocols, individual dietary intake and sleep behaviors were not directly controlled, representing potential confounding factors that may have influenced the magnitude or variability of physiological adaptations observed. Given the exploratory, systems-level intent of the correlation mapping, a conservative dual-threshold approach (p < 0.05 and |Δr| > 0.5) was preferred over strict Bonferroni correction, which may be overly stringent and reduce power in small samples. However, the risk of false positives remains and is acknowledged as a limitation of the present design.
Future research should aim to replicate these findings across more diverse athletic and clinical populations to evaluate the external validity of training-induced interaction patterns. Longitudinal studies extending across competitive cycles would be particularly valuable for determining the temporal stability of these physiological correlation patterns and their predictive utility for performance maintenance, injury prevention, or maladaptation. Advanced computational approaches, including dynamic Bayesian modeling or causality-inferred network analyses, may further elucidate the directionality and latency of systemic interactions under variable load conditions. Collectively, this study provides compelling evidence that HIIT induces both univariate performance gains and a qualitative reorganization of systemic interdependencies across physiological, cognitive, and neuromuscular domains. The emergence of robust cross-domain correlations and functionally coherent clusters in the HIIT group—but not in the control group—suggests that effective training adaptation is characterized not solely by improvements within isolated subsystems but by enhanced inter-system coordination. These findings substantiate a systems-level framework of performance, wherein multidomain integration serves as a core feature of functional adaptability.
5. Conclusions
This study provides novel evidence that high-intensity interval training (HIIT) induces not only improvements in isolated performance metrics—such as VO2max, lactate threshold speed, and BDNF—but also a systemic reorganization of physiological function, as reflected in emergent cross-domain correlations and hierarchical clustering patterns. In the HIIT group, previously independent variables spanning metabolic, cognitive, and neuromuscular domains formed tightly integrated clusters, suggesting that training enhances the dynamic coupling of these systems. In contrast, the control group showed domain-specific clustering without functional convergence, underscoring the specificity of the adaptive response to structured high-intensity training. While the emergence of multidomain integration following HIIT represents a robust group-level trend, it is important to acknowledge that such adaptations are not universal. Inter-individual variability in responsiveness—due to genetic, maturational, recovery-related, and psychophysiological factors—may influence the magnitude and stability of these systemic changes. Future research should aim to delineate such individual response profiles and their underlying determinants.
Practical Application: From an applied perspective, this underscores the importance of multidimensional monitoring strategies in elite sport. Practitioners should incorporate network-informed metrics—such as cognitive load indices, acid–base buffering markers (e.g., ΔBE), and inter-domain correlation profiles (lactate kinetics vs. exertion perception)—to evaluate systemic coherence alongside traditional outcome measures. Such an approach may facilitate the optimization of individualized training interventions that promote resilient, high-functioning athlete profiles.



 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}