
Analysis of Walnut Source–Sink–Flow Anatomical Structure Under Source–Sink Regulation Based on Fruit-Bearing Branch Scale

Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Artificial Regulation of Leaf-to-Fruit Ratio
2.3. Sample Collection
2.4. Preparing and Observing of Paraffin Sections
2.5. Data Analysis
3. Results
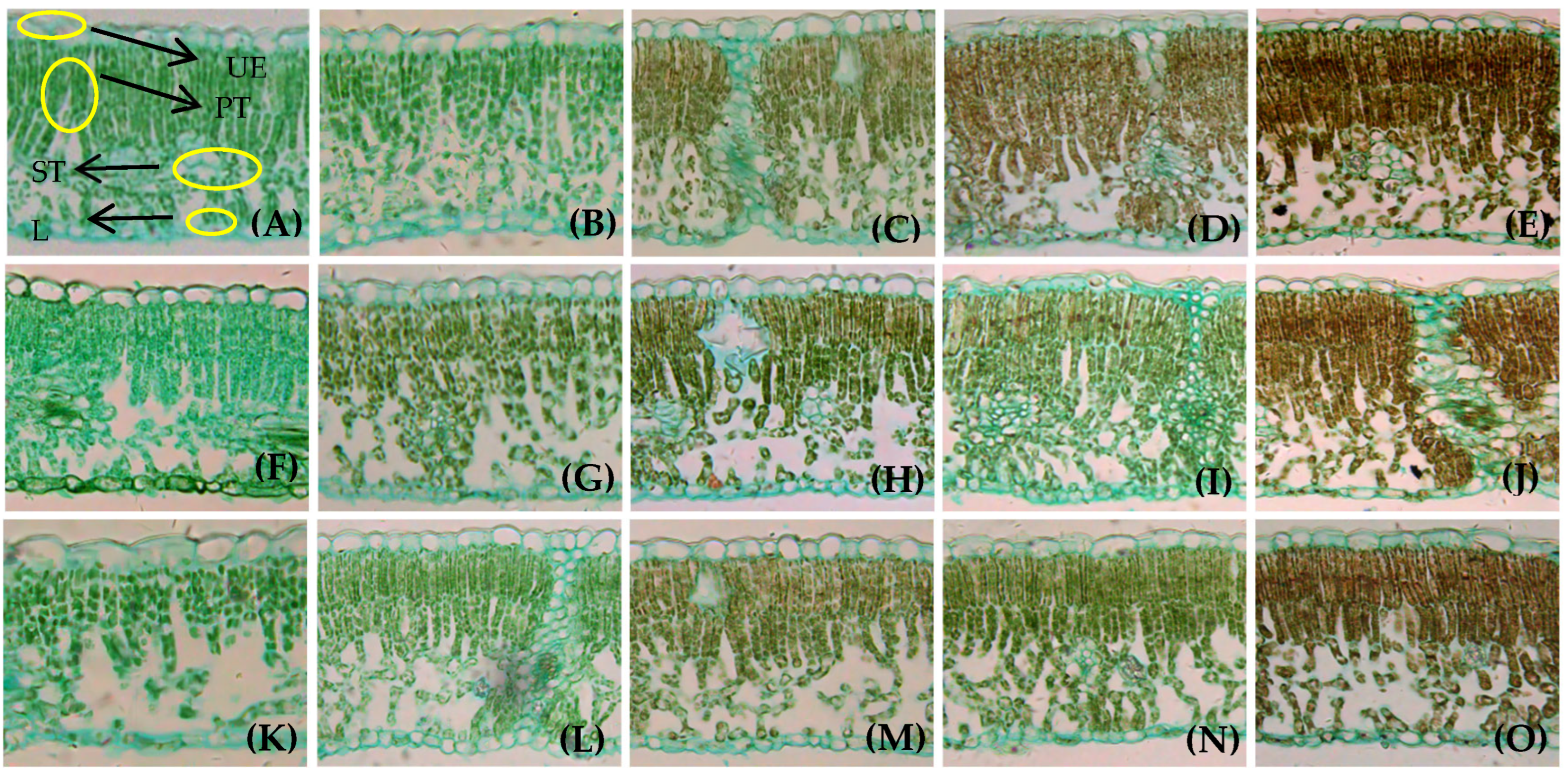
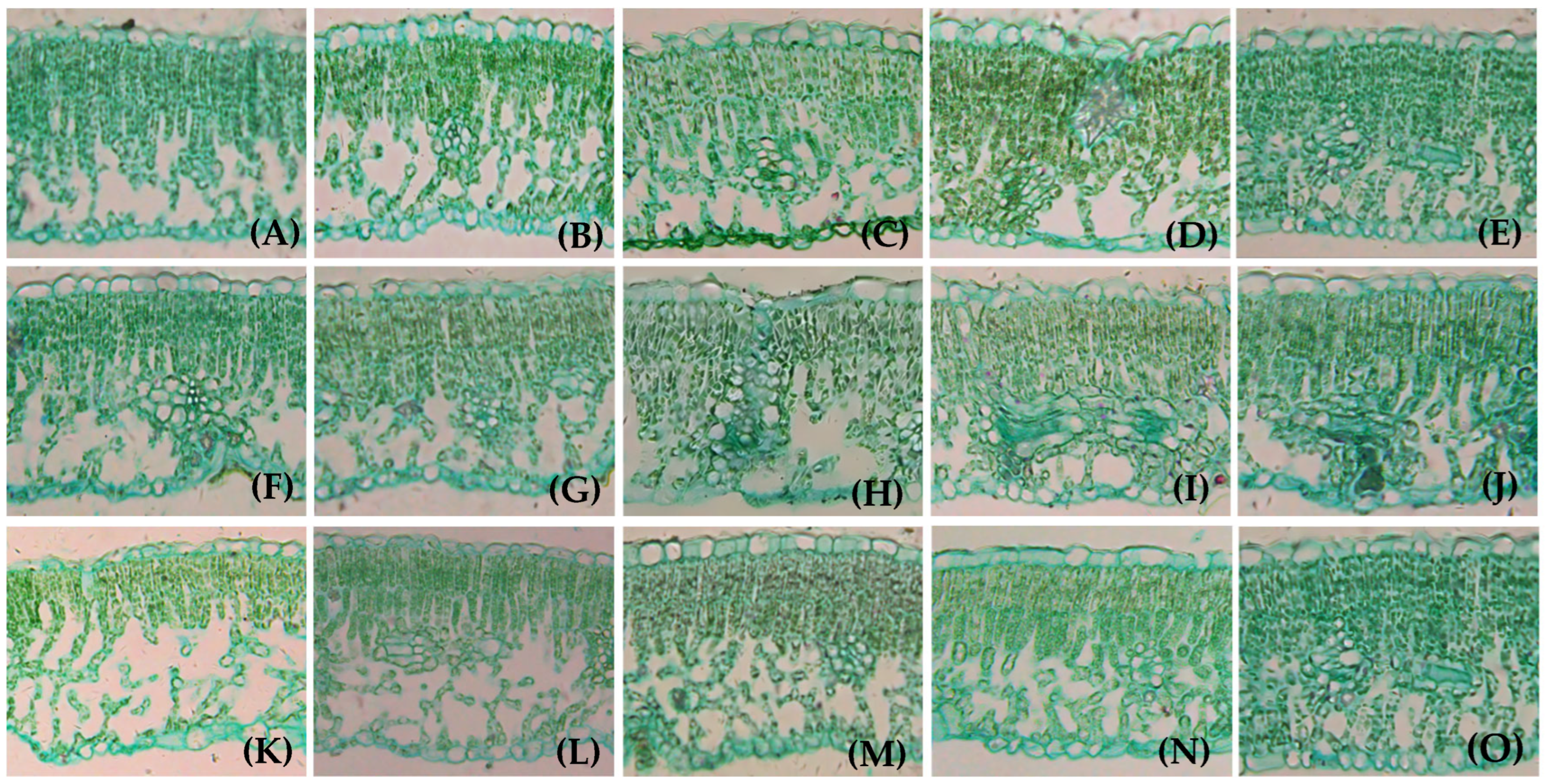
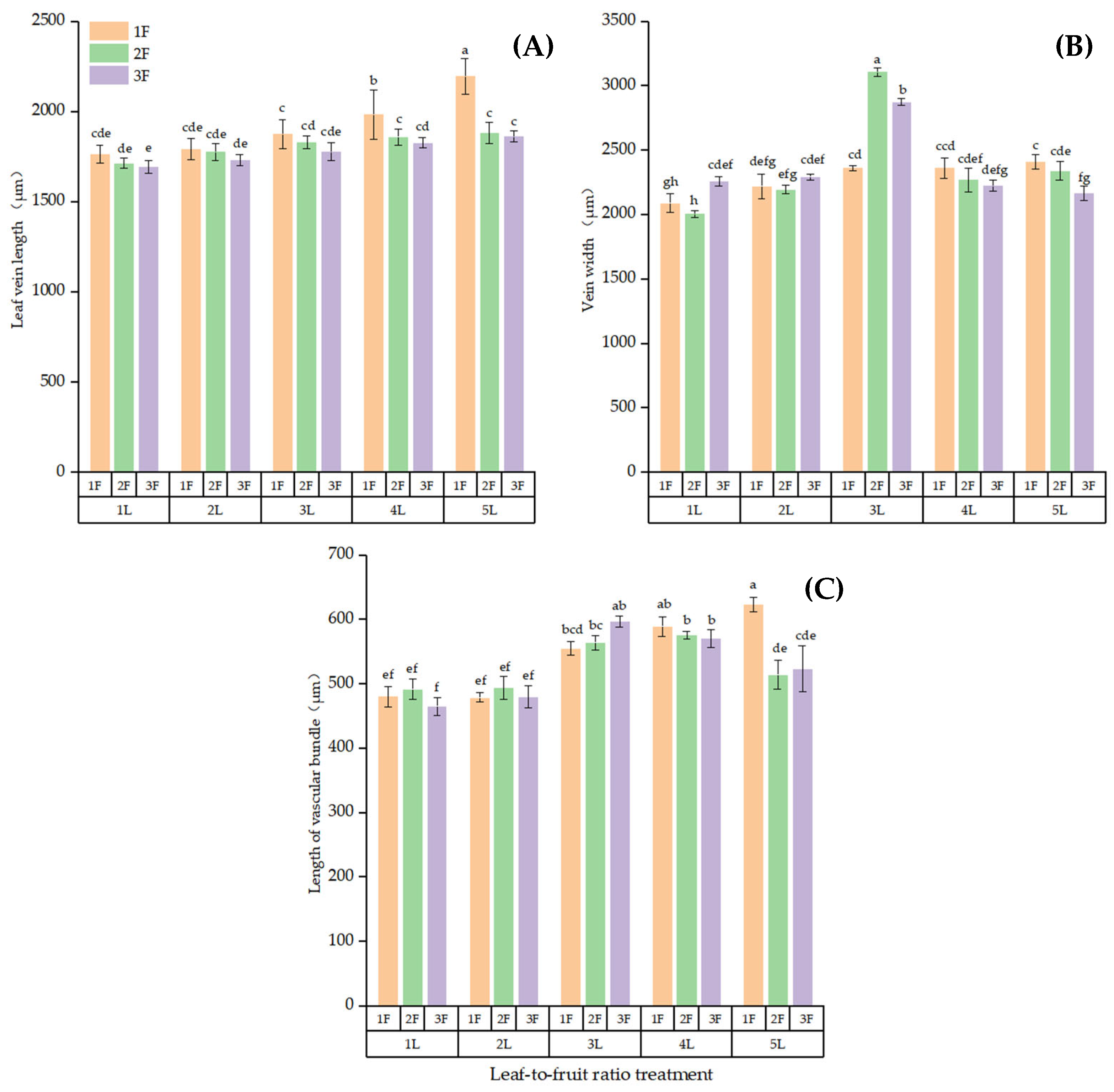
3.1. Effects of Different LFRs on Anatomical Structure of Branch Leaves
3.2. Morphological Structure of Leaf from Girdled Bearing Branches at Different LFRs
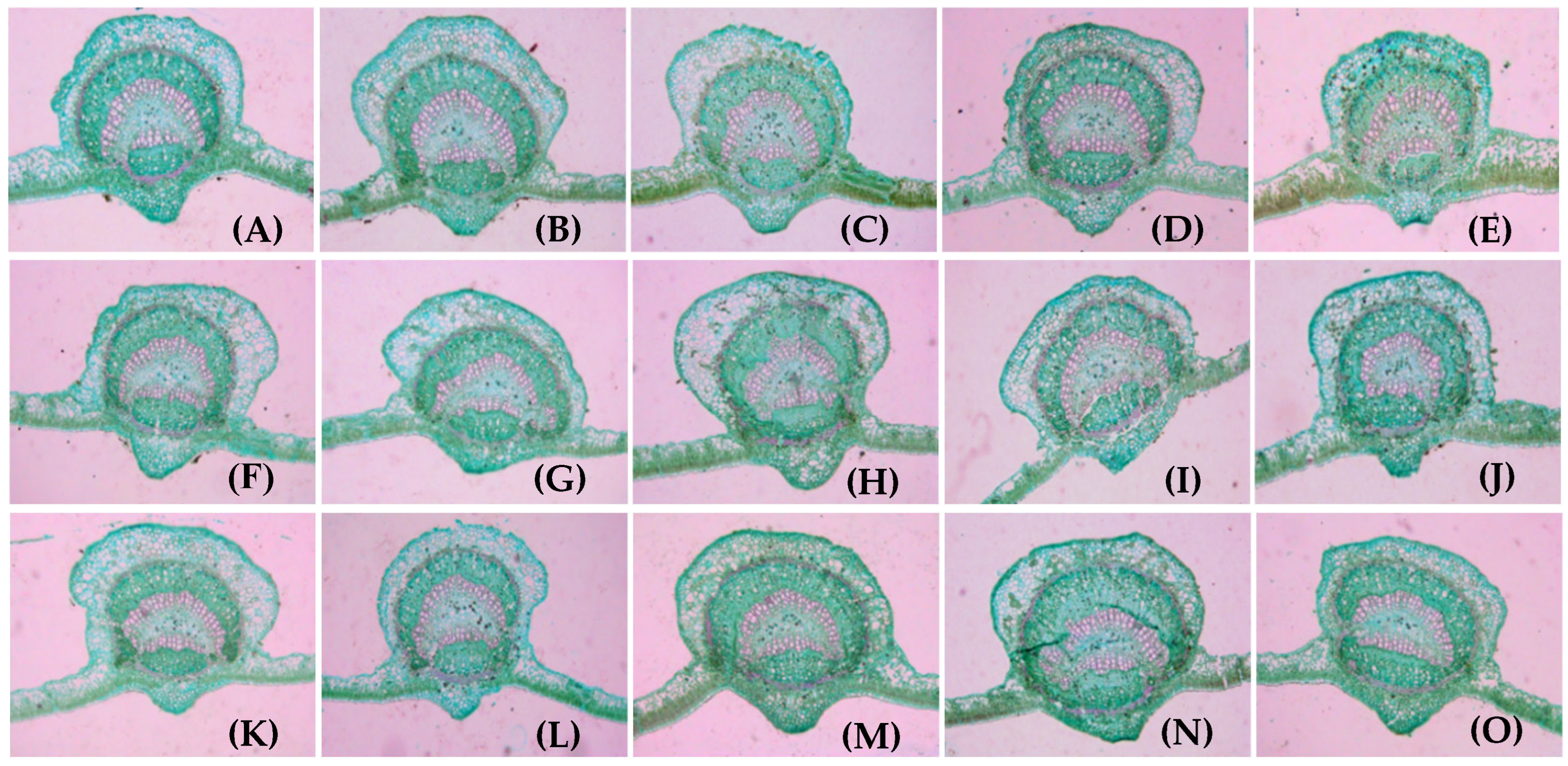
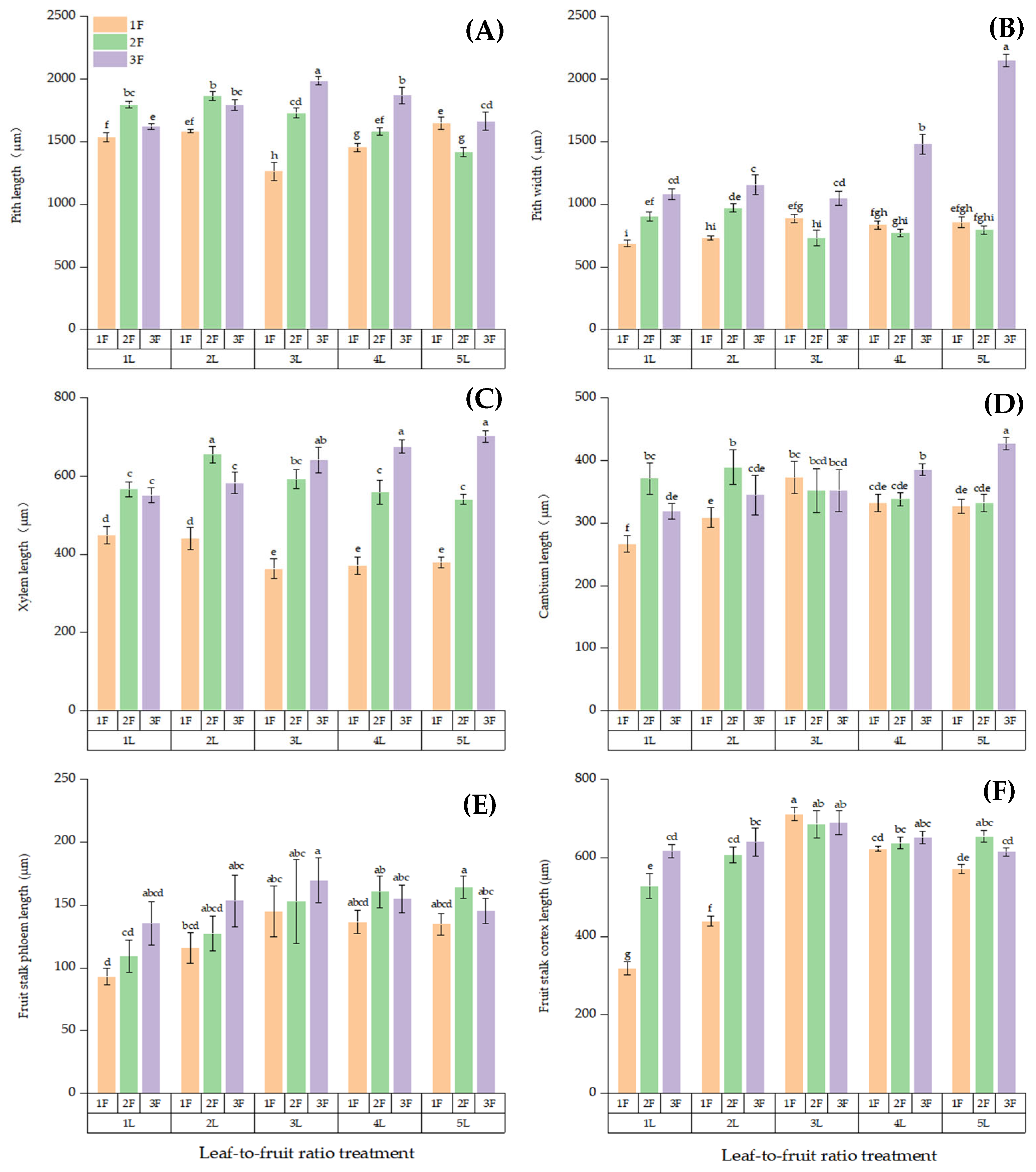
3.3. Anatomical Structure of Fruit Stalk on Girdled Branches at Different LFRs
3.4. Cell Structure of Fruit Stalks on Girdled Bearing Branches at Different LFRs
3.5. Anatomical Structure of Fruits on Branches with Different LFRs After Girdled
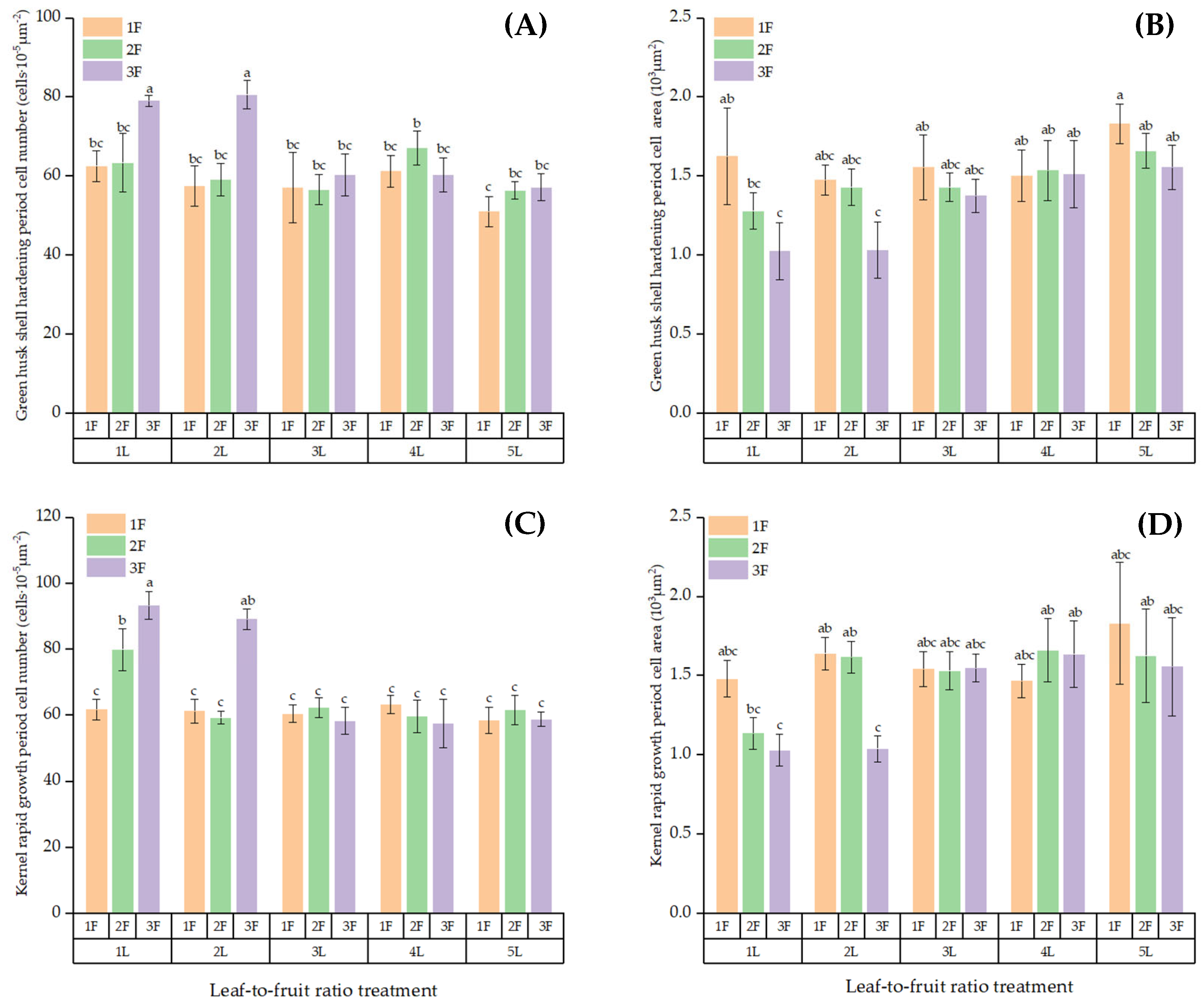
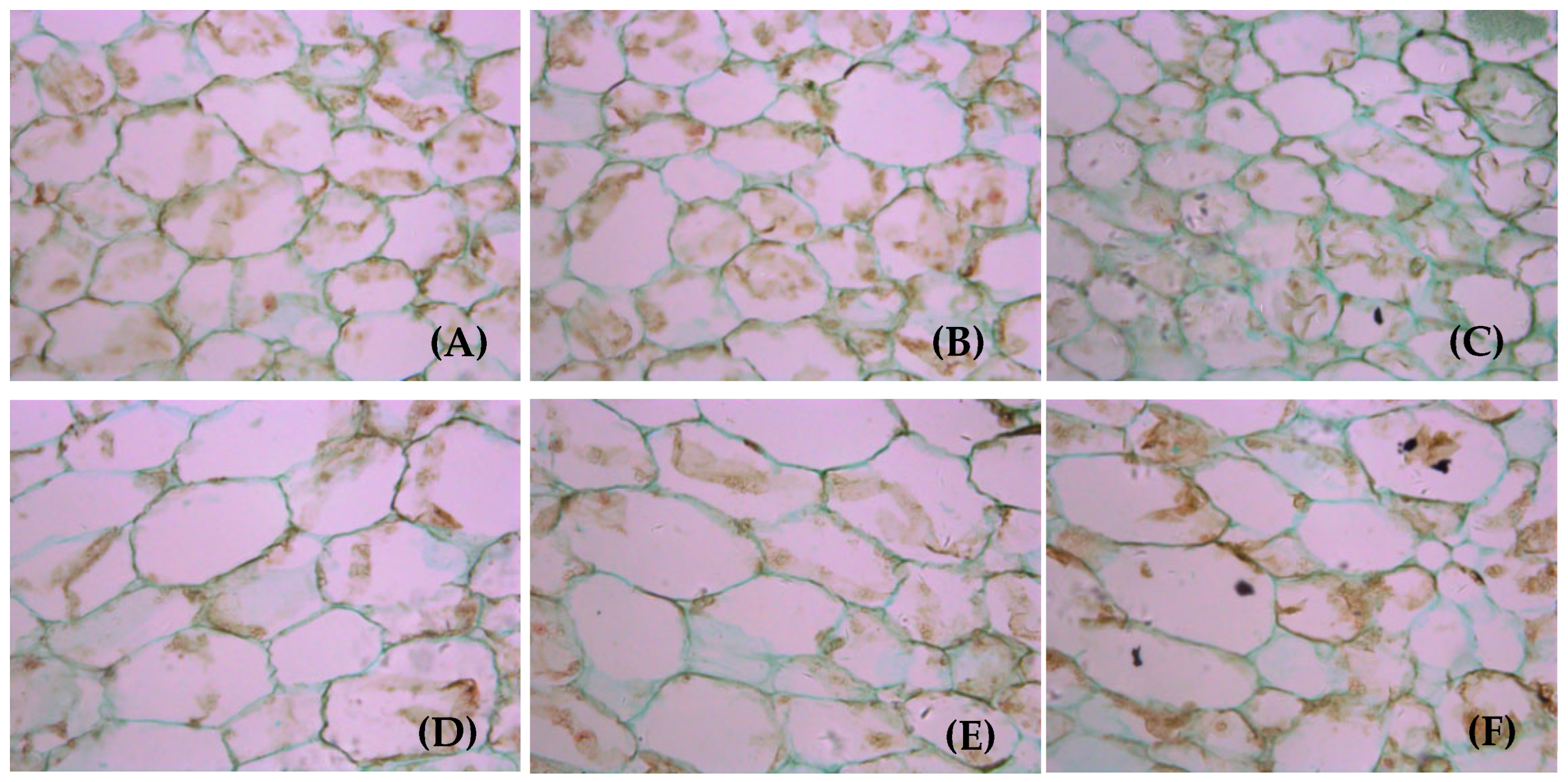
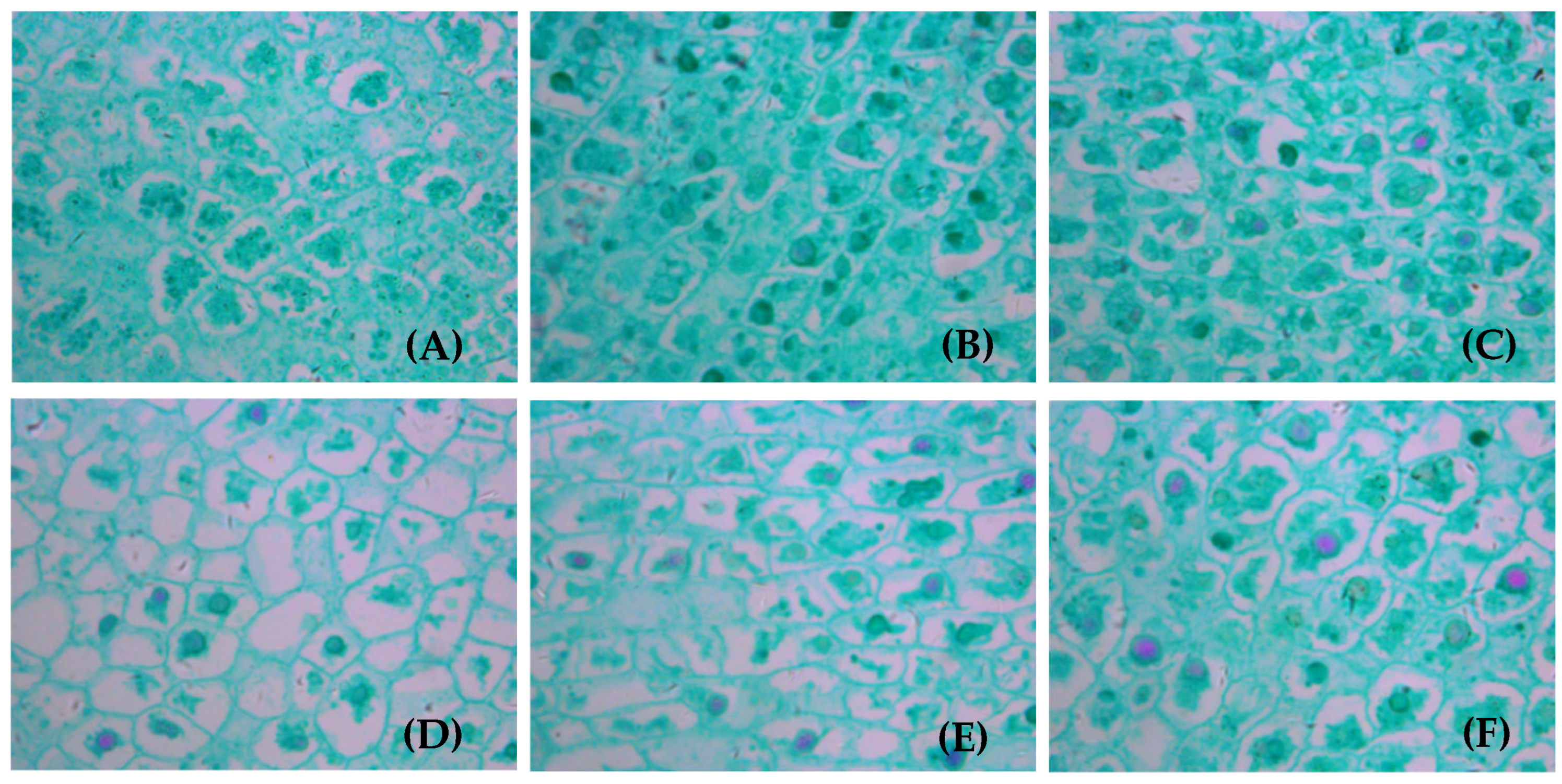
3.6. Cell Structure of Fruits on Girdled Branches with Different LFRs
4. Discussion
4.1. The Relationship Between Walnut Leaf Traits and LFR on the Girdled Bearing Branches
4.2. Relationship Between Fruit Stalk Traits and LFR on Girdled Bearing Branches
4.3. Relationship Between Fruit Traits and LFR on Girdled Bearing Branches
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Özcan, M.M.; Lemiasheuski, V. The Effect of Harvest Times on Mineral Contents of Almond and Walnut Kernels. Erwerbs-Obstbau 2020, 62, 455–458. [Google Scholar] [CrossRef]
- Quan, X.; Guō, Z.; Wang, S.; Li, H.; Gao, Z.; Wang, Y.; Cha, J. Analysis of morphological characteristics and fatty acid composition of walnut fruits from different varieties. Hunan For. Sci. 2020, 47, 7–14. [Google Scholar]
- Zhang, S.B. Association Analysis of the Impact of Leaf-to-Fruit Ratio on Carbohydrate Assimilation, Translocation, and Fruit Growth in Walnuts. Ph.D. Thesis, Xinjiang Agricultural University, Urumqi, China, 2023. [Google Scholar]
- Wang, X.Q.; Huang, W.D. Effects of weak light on the ultrastructure of phloem tissue in leaves of three-year-old oil peach (Prunus persica). Acta Bot. Sin. 2003, 45, 688–697. [Google Scholar]
- Chen, J.W.; Zhang, S.L.; Zhang, L.C.; Zhao, Z.Z.; Xu, J.G. Fruit Photosynthesis and Assimilate Translocation and Partitioning: Their Characteristics and Role in Sugar Accumulation in Developing Citrus unshiu Fruit. J. Integr. Plant Biol. 2002, 44, 158–163. [Google Scholar]
- Shen, C.H.; Chen, R.F.; Liu, Z.; Xi, R. Characteristics of Photosynthetic Product Transportation and Allocation During Fruit Expansion Period of Gaozhou Camellia oleifera. J. Cent. South Univ. For. Technol. 2024, 44, 58–66. [Google Scholar]
- Ma, Y.L.; Guo, S.J.; Liao, Y.N.; Wang, F. Effects of Different Canopy Layer Burr Loads on Photosynthetic Characteristics and Fruit Quality in Chestnut. Sci. Silvae Sin. 2022, 58, 90–97. [Google Scholar]
- Wang, S.; Pan, C.; Zhang, C.; Zhao, S. Key enzymes of sucrose metabolism play an important role in source-sink regulation of walnut fruit growth and development. N. Z. J. Crop Hortic. Sci. 2023, 51, 400–419. [Google Scholar] [CrossRef]
- Zhang, S.B.; Chen, H.; Pan, C.D. Effects of the Source-Sink Relationship on Walnut Nut Quality at the Scale of the Fruit-Bearing Branchs. Forest 2022, 13, 1034. [Google Scholar] [CrossRef]
- He, C.C.; Zhare, M.; Abudureyimu, A.; Yili, K. Effects of Different Load Levels on Photosynthetic Efficiency and Chloroplast Ultrastructure of Korla Fragrant Pear. J. Xinjiang Agric. Univ. 2012, 35, 368–372. [Google Scholar]
- Wu, T.; Tao, S.T.; Zhang, H.P.; Song, Y.; Yao, G.; Zhang, S. Effects of fruit thinning on sugar accumulastion in pear fruits and photosynthetic characteristics of leaves. Acta Hortic. Sin. 2011, 38, 2041–2048. [Google Scholar]
- Wu, H.B. Photosynthetic Physiological Characteristics and Photosynthate Allocation during the Development of Korean Pine Cones. Ph.D. Thesis, Northeast Forestry University, Harbin, China, 2022. [Google Scholar]
- Jackson, D.; Coombe, B. The growth of apricot fruit. I. Morphological changes during development and the effects of various tree factors. Aust. J. Agric. Res. 1966, 17, 465. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Peng, Y.B.; Pelleschi-Travier, S.; Fan, Y.; Lu, Y.F.; Lu, Y.M.; Gao, X.P.; Shen, Y.Y.; Delrot, S.; Zhang, D.P. Evidence for Apoplasmic Phloem Unloading in Developing Apple Fruit. Plant Physiol. 2004, 135, 574–586. [Google Scholar] [CrossRef]
- Wang, J.; Chen, W.; Peng, J.; Liu, Y.; Liang, K.; Li, C.; Mao, X.; Pan, J. Study on the “Source-Sink-Flow” Characteristics and Super High-Yield Potential of the Extra-Large Panicle Rice Germplasm DS23 in South China. Guangdong Agric. Sci. 2023, 50, 150–159. [Google Scholar]
- Zhang, Y.; Ren, F.X.; Jing, J.J.; Du, H.; Sun, H.C.; Liu, L.T.; Bai, Z.Y.; Li, C.D. Effects of leaf and branch removal on the anatomical structure of cotton leaves, petioles, boll stalks, and yield. Cotton Sci. 2016, 28, 268–275. [Google Scholar]
- Wang, Q.J.Y.; Li, J.; Li, T.; Xing, Y.; Qing, L.; Fang, K. Anatomical Structure and Cell Wall Components Changes in Chestnut ‘Yanshan Hongli’ Female Flowers at Developmental Stages. Bull. Bot. Res. 2024, 44, 670–680. [Google Scholar]
- Xu, J.W.; Tang, J.M.; Jiang, H.L.; Zou, R.; Wei, X. A comparative study on leaf anatomy and photosynthetic characteristics of different growth stages of Horsfieldia hainanensis. PeerJ 2024, 12, e18640. [Google Scholar] [CrossRef]
- Li, K.T.; Syvertsen, J.P. Morphological Responses Induced by Walnut Leaf Blight Disease. Nonwood For. Res. 2024, 42, 188–196. [Google Scholar]
- Zhang, S.B.; Zhang, R.; Xu, C.Z.; Guo, L.; Gao, S.; Guo, L.; Zhang, J. Effects of shading on physiological characteristics of Juglans regia ‘Wen 185’ leaves. North. Hortic. 2017, 18, 28–34. [Google Scholar]
- Zhang, S.; Chongzhi, X.U.; Zhang, R.; Zhou, W.; Gao, S.; Guo, L. Effects of shading on photosynthetic characteristics of Juglans regia ‘Wen 185’. China Fruits 2017, 3, 22–27. [Google Scholar]
- Fang, J.B.; Zheng, F.; Chen, J.Y.; Tian, L.; Zhang, W. The Effect of Source-Sink Relationship Changes on Photosynthetic Intensity in Kiwifruit Leaves. In Proceedings of the 9th Annual Academic Conference of the Chinese Society for Horticultural Science, Chongqing, China, 1 October 2001. [Google Scholar]
- Sun, H.Q.; Wang, N.N.; Liu, W.J.; Ma, C.H. Analysis of pear fruit stalk characteristics and their correlation with main economic traits of fruits. Acta Bot. Boreali-Occident. Sin. 2019, 39, 1416–1424. [Google Scholar]
- Cao, Y.J.; Maimaiti, A.; Tayifu, X.; Shi, Z.; Abulitipu, Y. Comparative Analysis of Different Types of Fruit Stalks in Korla Fragrant Pear. Xinjiang Agric. Sci. 2023, 60, 1442–1450. [Google Scholar]
- Lalonde, S.; Tegeder, M.; Throne-Holst, M.; Frommer, W.B.; Patrick, J.W. Phloem loading and unloading of sugars and amino acids. Plant Cell Environ. 2010, 26, 37–56. [Google Scholar] [CrossRef]
- Cai, X.L.; Pan, J.C.; Zhou, Y.M.; Liu, H.H.; Huang, G.X. Research on the Mathematical Model of Fruit Growth and Development of Taiwan Pineapple Sugar Apple. J. Anhui Agric. Sci. 2016, 44, 44–45. [Google Scholar]
- Garcia-Luis, A.; Oliveira, M.E.M.; Bordón, Y.; Siqueira, D.L.; Tominaga, S.; Guardiola, J.L. Dry Matter Accumulation in Citrus Fruit is Not Limited by Transport Capacity of the Pedicel. Ann. Bot. 2002, 90, 755–764. [Google Scholar] [CrossRef]
- Zhao, P.; Xia, Y.; He, J.Y.; Wu, J.; Ma, J.Q.; Huang, C.S. Effects of leaf-fruit ratio on 13C assimilate partitioning and fruit quality in mulberry during veraison. Acta Sericologica Sin. 2023, 49, 1–7. [Google Scholar]
- Cannell, M.G.R.; Dewar, R.C. Carbon Allocation in Trees: A Review of Concepts for Modelling. Trees 1994, 25, 59–104. [Google Scholar]
- Smith, A.M.; Stitt, M. Coordination of carbon supply and plant growth. Plant Cell Environ. 2007, 30, 1126–1149. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response Variable | df | SS | MS | F Value | p Value |
|---|---|---|---|---|---|
| Leaf thickness (LT) | 14 | 19,840.28 | 1417.16 | 52.41 | 0.000 |
| Thickness of palisade tissue (PTT) | 14 | 4472.30 | 319.45 | 25.21 | 0.000 |
| Thickness of spongy tissue (STT) | 14 | 3159.05 | 225.65 | 16.40 | 0.000 |
| Thickness of upper epidermis (UET) | 14 | 115.51 | 8.25 | 3.98 | 0.001 |
| Thickness of lower epidermis (LET) | 14 | 13.15 | 0.94 | 1.11 | 0.162 |
| Area of main vein phloem (MVPA) | 14 | 4258.29 | 304.16 | 36.61 | 0.000 |
| Response Variable | df | SS | MS | F Value | p Value |
|---|---|---|---|---|---|
| Area of fruit stalk Phloem (FSPA) | 14 | 0.382 | 0.027 | 5.799 | 0.000 |
| LFR | 1L:1F | 2L:1F | 3L:1F | 4L:1F | 5L:1F |
| FSPA (105 μm2) | 1.05 ± 0.04 c | 1.04 ± 0.03 c | 1.06 ± 0.10 c | 1.08 ± 0.07 c | 1.05 ± 0.04 c |
| LFR | 1L:2F | 2L:2F | 3L:2F | 4L:2F | 5L:2F |
| FSPA (105 μm2) | 1.15 ± 0.04 bc | 1.25 ± 0.09 ab | 1.17 ± 0.10 bc | 1.26 ± 0.10 ab | 1.23 ± 0.04 ab |
| LFR | 1L:3F | 2L:3F | 3L:3F | 4L:3F | 5L:3F |
| FSPA (105 μm2) | 1.23 ± 0.05 ab | 1.25 ± 0.11 ab | 1.26 ± 0.01 ab | 1.23 ± 0.05 ab | 1.31 ± 0.02 a |
| Response Variable | df | SS | MS | F Value | p Value |
|---|---|---|---|---|---|
| Cell number in green husk | 14 | 2790.55 | 199.33 | 8.93 | 0.000 |
| Cell area in green husk | 14 | 1.94 | 0.14 | 5.09 | 0.000 |
| Cell number in kernel | 14 | 5717.47 | 408.39 | 24.16 | 0.000 |
| Cell area in kernel | 14 | 2.33 | 0.17 | 4.78 | 0.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, L.; Zhang, C.; Yang, Z.; Zhao, Z.; Wang, S. Analysis of Walnut Source–Sink–Flow Anatomical Structure Under Source–Sink Regulation Based on Fruit-Bearing Branch Scale. Appl. Sci. 2025, 15, 7523. https://doi.org/10.3390/app15137523
Lv L, Zhang C, Yang Z, Zhao Z, Wang S. Analysis of Walnut Source–Sink–Flow Anatomical Structure Under Source–Sink Regulation Based on Fruit-Bearing Branch Scale. Applied Sciences. 2025; 15(13):7523. https://doi.org/10.3390/app15137523
Chicago/Turabian StyleLv, Luyu, Cuifang Zhang, Zhilong Yang, Zeyun Zhao, and Shiwei Wang. 2025. "Analysis of Walnut Source–Sink–Flow Anatomical Structure Under Source–Sink Regulation Based on Fruit-Bearing Branch Scale" Applied Sciences 15, no. 13: 7523. https://doi.org/10.3390/app15137523
APA StyleLv, L., Zhang, C., Yang, Z., Zhao, Z., & Wang, S. (2025). Analysis of Walnut Source–Sink–Flow Anatomical Structure Under Source–Sink Regulation Based on Fruit-Bearing Branch Scale. Applied Sciences, 15(13), 7523. https://doi.org/10.3390/app15137523