
Protective Effects of the Ethyl Acetate Fraction of Distylium racemosum Against Metabolic Dysfunction-Associated Steatohepatitis

 , , , , , and
, , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of D. racemosum Fractions
2.2. Cell Viability Assay
2.3. Lipid Accumulation Analysis
2.4. Real-Time Quantitative Polymerase Chain Reaction
2.5. MASH Model Induction via a High-Fat Methionine- and Choline-Deficient (HFMCD) Diet
2.6. Tissue Collection and Histological Analysis
2.7. HPLC Profiling of the Active Components
2.8. Statistical Analyses
3. Results
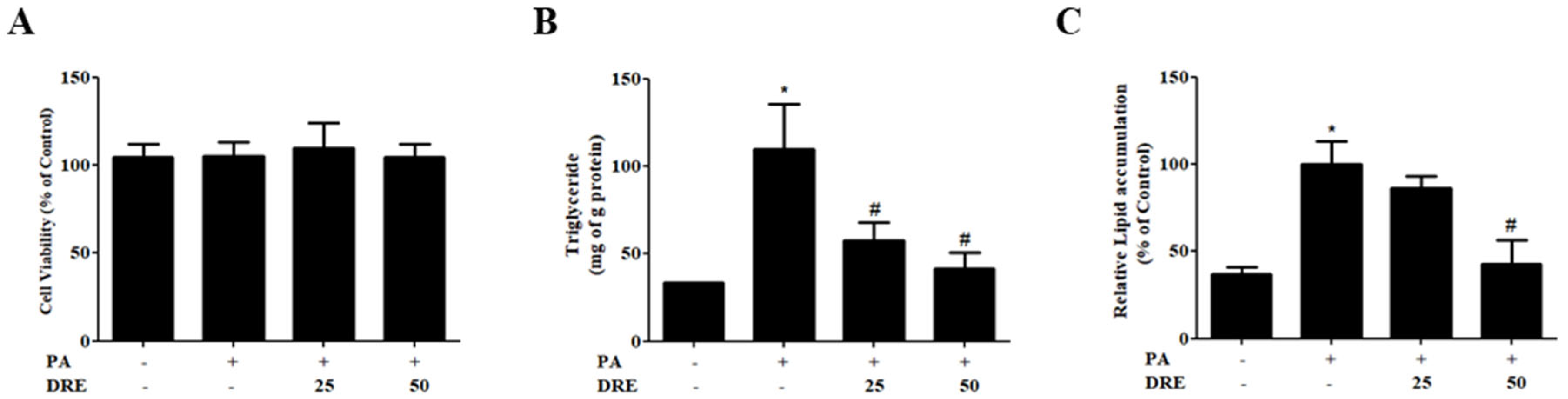
3.1. Effects of DRE on Cell Viability and Lipid Accumulation in PA-Induced AML12 Cells
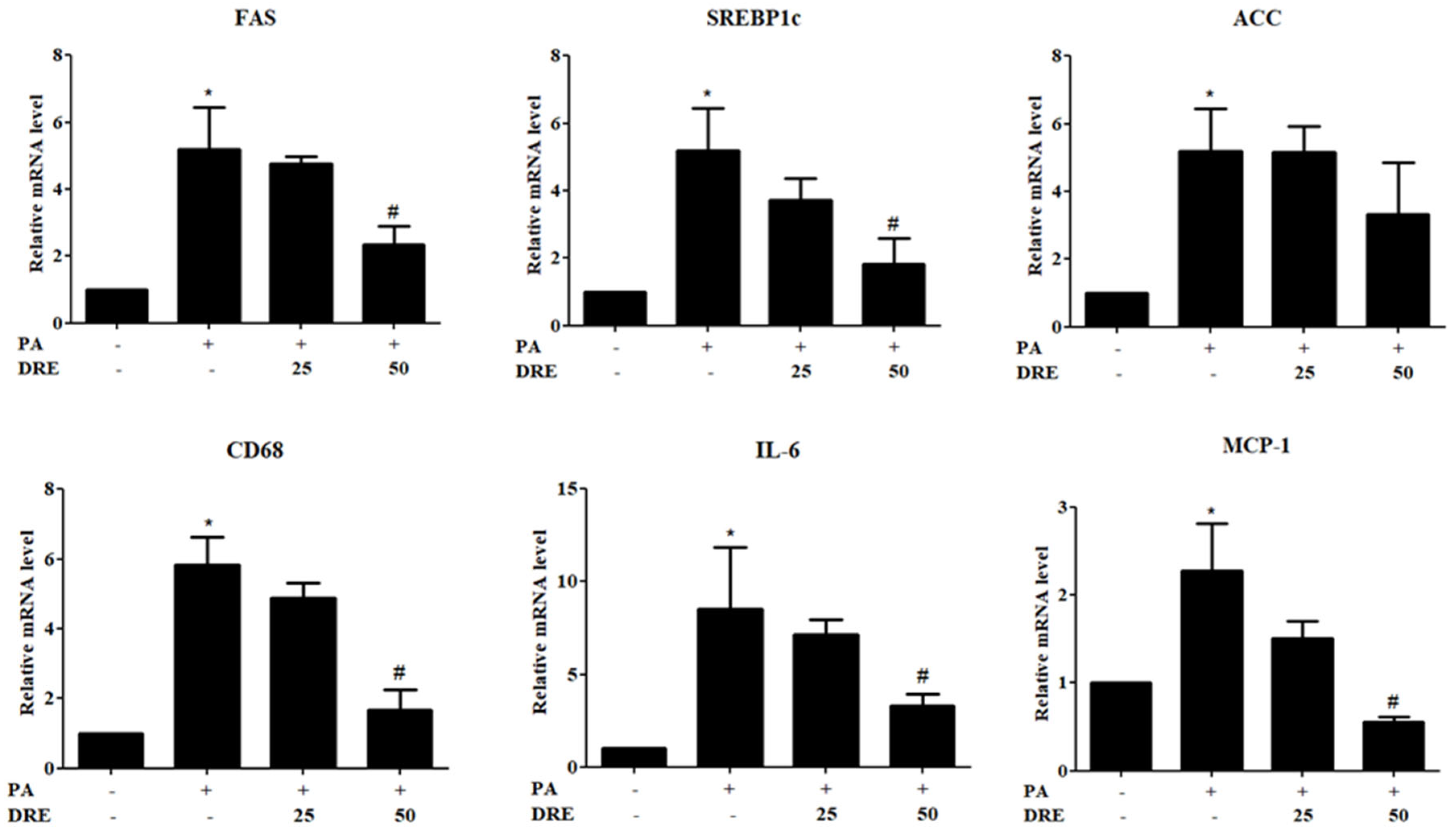
3.2. Effects of DRE on PA-Induced Lipogenesis and Inflammatory Gene Expression Levels in Hepatocytes
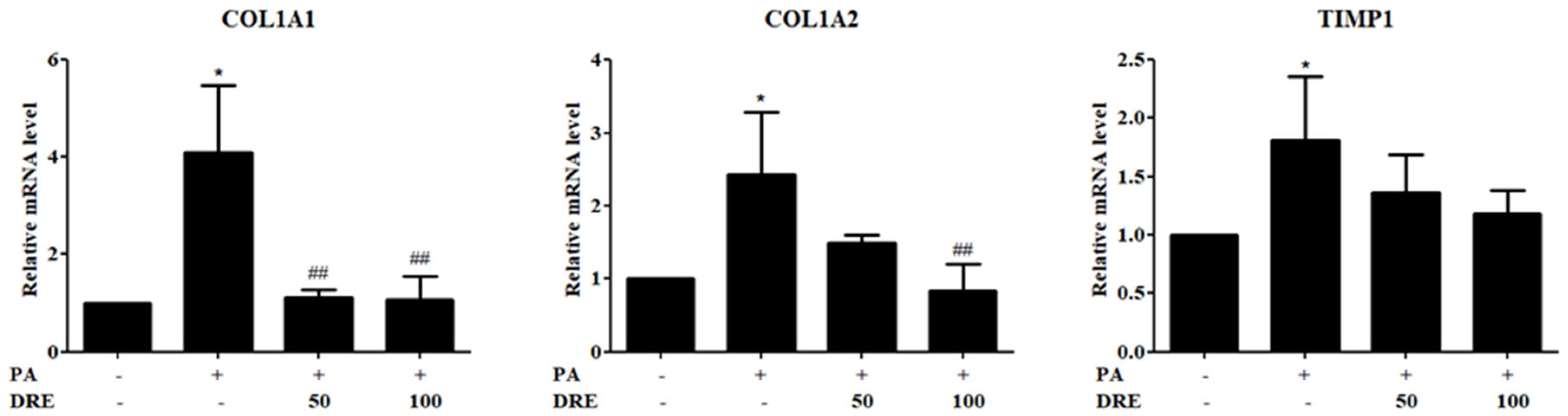
3.3. DRE Attenuates Fibrogenic Gene Expression in PA-Induced AML12 Cells
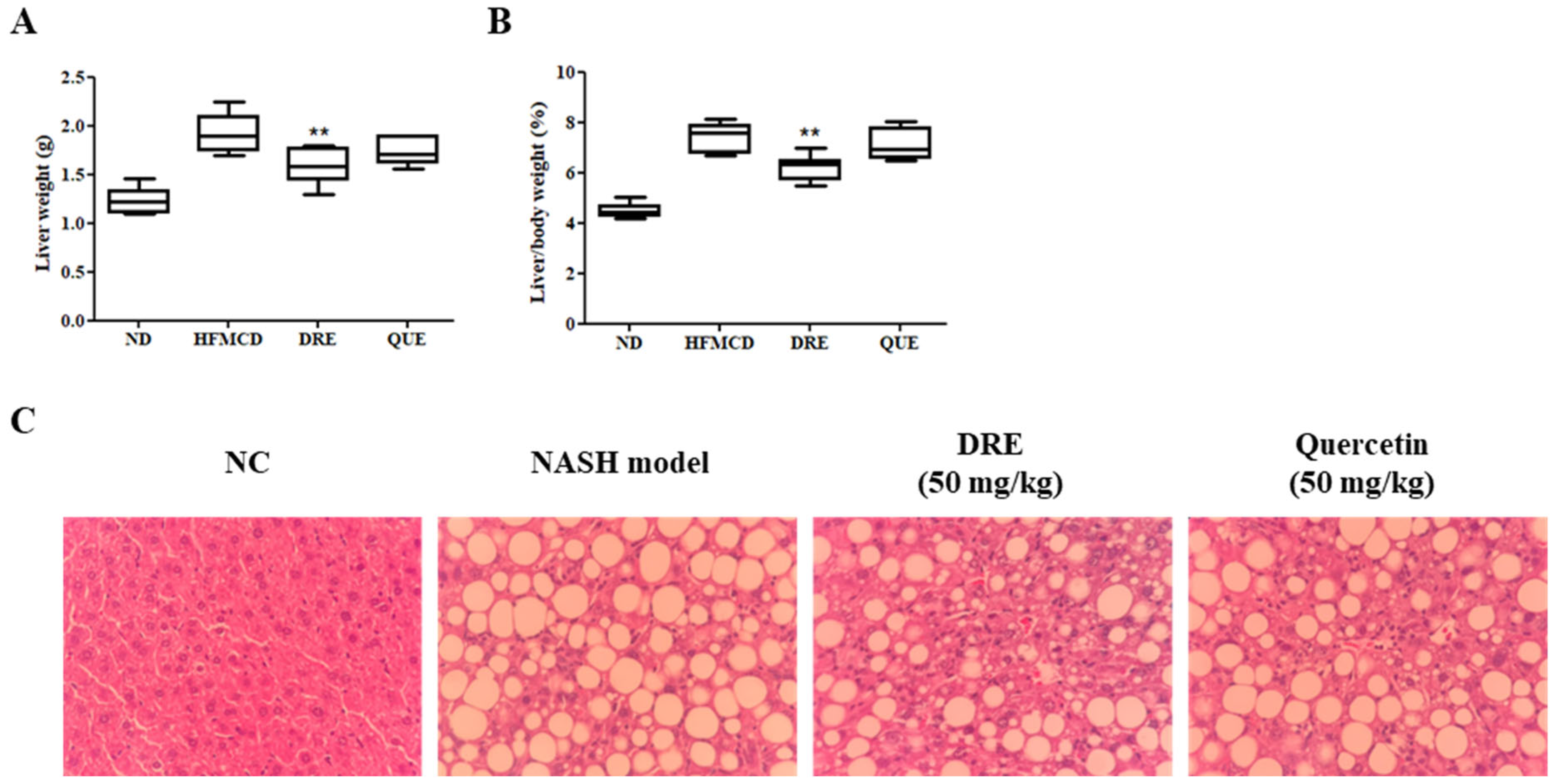
3.4. DRE Reduces the Liver Weight and Improves the Liver Histology in HFMCD Diet-Induced MASH Model Mice
3.5. DRE Modulates the Gene Expression Levels in HFMCD Diet-Induced MASH Model Mice
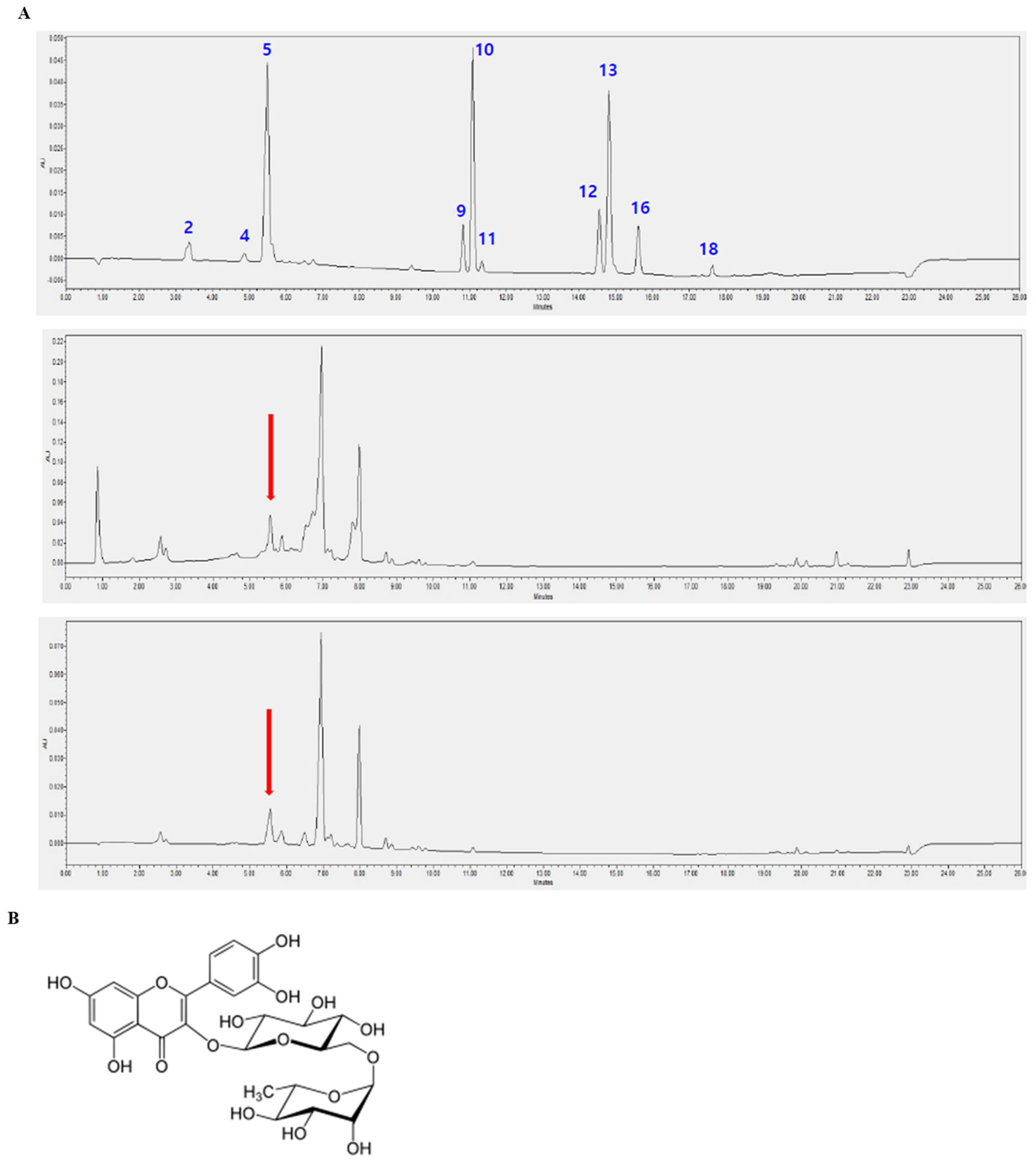
3.6. Identification of Rutin in D. racemosum Extracts via HPLC Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACC | Acetyl-CoA carboxylase |
| COL1A1 | Collagen type I alpha 1 |
| CPT-1 | Carnitine palmitoyltransferase-1 |
| DRE | Distylium racemosum ethyl acetate fraction |
| FAS | Fatty acid synthase |
| HFMCD | High-fat methionine- and choline-deficient |
| HPLC | High-performance liquid chromatography |
| NAFLD | Non-alcoholic fatty liver disease |
| MASH | Metabolic dysfunction-associated steatohepatitis |
| PA | Palmitic acid |
| SREBP1c | Sterol regulatory element-binding protein 1c |
| TG | Triglyceride |
| TIMP1 | Tissue inhibitor of metalloproteinases 1 |
References
- Sheka, A.C.; Adeyi, O.; Thompson, J.; Hameed, B.; Crawford, P.A.; Ikramuddin, S. Nonalcoholic steatohepatitis. JAMA 2020, 323, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Abdelmalek, M.F. Nonalcoholic fatty liver disease: Another leap forward. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 85–86. [Google Scholar] [CrossRef]
- Raza, S.; Rajak, S.; Upadhyay, A.; Tewari, A.; Sinha, R. Current treatment paradigms and emerging therapies for NAFLD/NASH. Front. Biosci. 2021, 26, 206–237. [Google Scholar] [CrossRef] [PubMed]
- Powell, E.E.; Wong, V.W.; Rinella, M. Non-alcoholic fatty liver disease. Lancet 2021, 397, 2212–2224. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Wang, L.; Evans, P.C.; Xu, S. NAFLD and NASH: Etiology, targets and emerging therapies. Drug Discov. Today 2024, 29, 103910. [Google Scholar] [CrossRef]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver (EASL); European Association for the Study of Diabetes (EASD); European Association for the Study of Obesity (EASO). EASL-EASD-EASO Clinical Practice Guidelines on the management of metabolic dysfunction-associated steatotic liver disease (MASLD). J. Hepatol. 2024, 81, 492–542. [Google Scholar] [CrossRef]
- Huby, T.; Gautier, E.L. Immune cell-mediated features of non-alcoholic steatohepatitis. Nat. Rev. Immunol. 2022, 22, 429–443. [Google Scholar] [CrossRef]
- Huang, Q.; An, Z.; Xin, X.; Gou, X.; Tian, X.; Hu, Y.; Mei, Z.; Feng, Q. The Effectiveness of Curcumin, Resveratrol, and Silymarin on MASLD: A Systematic Review and Meta-Analysis. Food Sci. Nutr. 2024, 12, 10010–10029. [Google Scholar] [CrossRef]
- Kim, H.R.; Jung, B.K.; Yeo, M.H.; Yoon, W.J.; Chang, K.S. Inhibition of lipid accumulation by the ethyl acetate fraction of Distylium racemosum in vitro and in vivo. Toxicol. Rep. 2019, 23, 215–221. [Google Scholar] [CrossRef]
- Larsson, S.; Wolk, A. Overweight, obesity and risk of liver cancer: A meta-analysis of cohort studies. Br. J. Cancer 2007, 97, 1005–1008. [Google Scholar] [CrossRef] [PubMed]
- Nagappan, A.; Jung, D.Y.; Kim, J.H.; Lee, H.; Jung, M.H. Gomisin N alleviates ethanol-induced liver injury through ameliorating lipid metabolism and oxidative stress. Int. J. Mol. Sci. 2018, 19, 2601. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Jung, D.Y.; Kim, S.; Jung, M.H. Preventive effect of poricoic acid against nonalcoholic steatohepatitis. J. Life Sci. 2022, 32, 962–970. [Google Scholar]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease—Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Tiniakos, D.G.; Vos, M.B.; Brunt, E.M. Nonalcoholic fatty liver disease: Pathology and pathogenesis. Annu. Rev. Pathol. 2010, 5, 145–171. [Google Scholar] [CrossRef]
- Jadeja, R.; Devkar, R.V.; Nammi, S. Herbal Medicines for the Treatment of Nonalcoholic Steatohepatitis: Current Scenario and Future Prospects. Evid. Based Complement. Alternat. Med. 2014, 2014, 648308. [Google Scholar] [CrossRef]
- Dibal, N.I.; Garba, S.H.; Jacks, T.W. Onion Peel Quercetin Attenuates Ethanol-Induced Liver Injury in Mice by Preventing Oxidative Stress and Steatosis. Biomed. Res. Ther. 2022, 9, 5102–5112. [Google Scholar] [CrossRef]
- Ann, J.Y.; Eo, H.; Lim, Y. Mulberry Leaves (Morus alba L.) Ameliorate Obesity-Induced Hepatic Lipogenesis, Fibrosis, and Oxidative Stress in High-Fat Diet-Fed Mice. Genes Nutr. 2015, 10, 46. [Google Scholar] [CrossRef]
- Li, W.; Lu, Y. Hepatoprotective Effects of Sophoricoside against Fructose-Induced Liver Injury via Regulating Lipid Metabolism, Oxidation, and Inflammation in Mice. J. Food Sci. 2018, 83, 552–558. [Google Scholar] [CrossRef]
- Ganeshpurkar, A.; Saluja, A.K. The pharmacological potential of rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef]
- Loguercio, C.; Festi, D. Silybin and the liver: From basic research to clinical practice. World J. Gastroenterol. 2011, 17, 2288–2301. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Moustaid-Moussa, N.; Chen, L.; Mo, H.; Shastri, A.; Su, R.; Bapat, P.; Kwun, I.; Shen, C.L. Novel insights of dietary polyphenols and obesity. J. Nutr. Biochem. 2014, 25, 1–18. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-H.; Yeo, M.-H.; Chang, K.-S.; Yoon, W.-J.; Kim, H.-S.; Kim, J.; Kim, H.-R. Protective Effects of the Ethyl Acetate Fraction of Distylium racemosum Against Metabolic Dysfunction-Associated Steatohepatitis. Appl. Sci. 2025, 15, 7238. https://doi.org/10.3390/app15137238
Lee Y-H, Yeo M-H, Chang K-S, Yoon W-J, Kim H-S, Kim J, Kim H-R. Protective Effects of the Ethyl Acetate Fraction of Distylium racemosum Against Metabolic Dysfunction-Associated Steatohepatitis. Applied Sciences. 2025; 15(13):7238. https://doi.org/10.3390/app15137238
Chicago/Turabian StyleLee, Young-Hyeon, Min-Ho Yeo, Kyung-Soo Chang, Weon-Jong Yoon, Hye-Sook Kim, Jongwan Kim, and Hye-Ran Kim. 2025. "Protective Effects of the Ethyl Acetate Fraction of Distylium racemosum Against Metabolic Dysfunction-Associated Steatohepatitis" Applied Sciences 15, no. 13: 7238. https://doi.org/10.3390/app15137238
APA StyleLee, Y.-H., Yeo, M.-H., Chang, K.-S., Yoon, W.-J., Kim, H.-S., Kim, J., & Kim, H.-R. (2025). Protective Effects of the Ethyl Acetate Fraction of Distylium racemosum Against Metabolic Dysfunction-Associated Steatohepatitis. Applied Sciences, 15(13), 7238. https://doi.org/10.3390/app15137238