
Usage of a Sensory-Motor Intervention System for Understanding the Adaptive Behavior of Insects


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
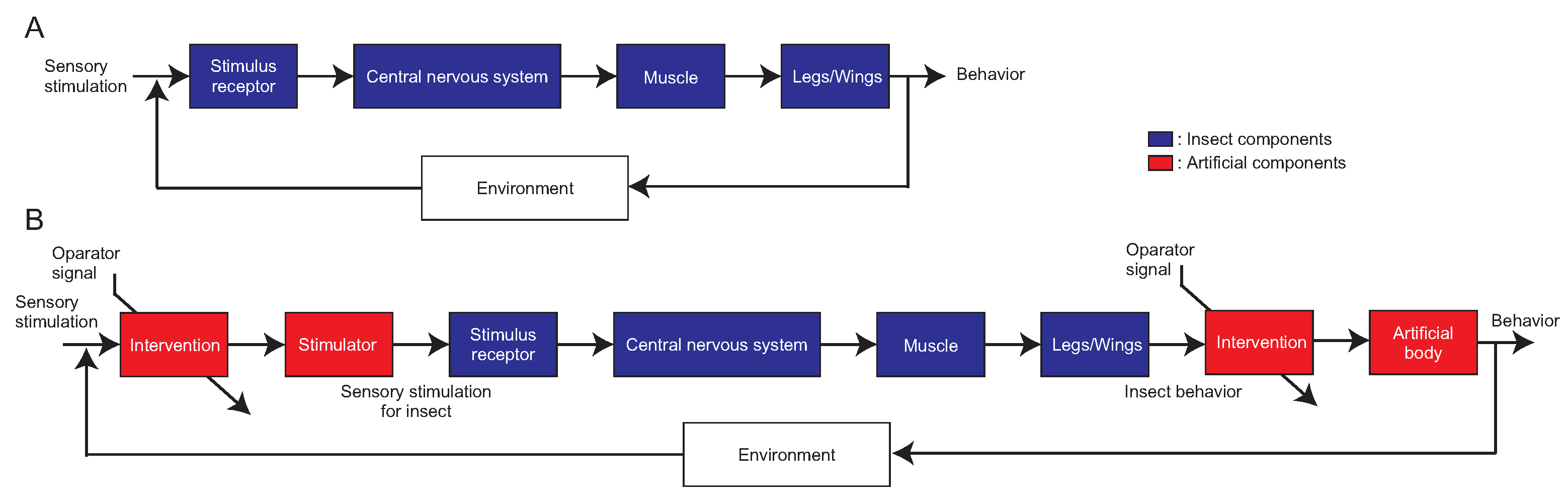
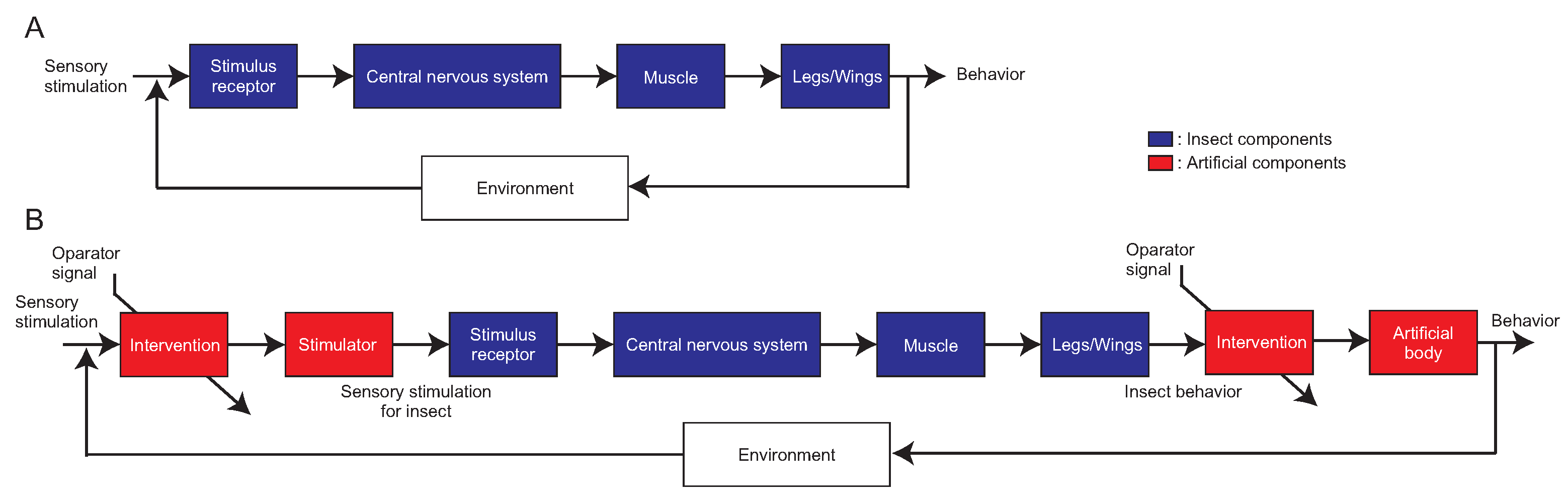
2. Overview of the Biological Experimental Setup Using Engineering Tools
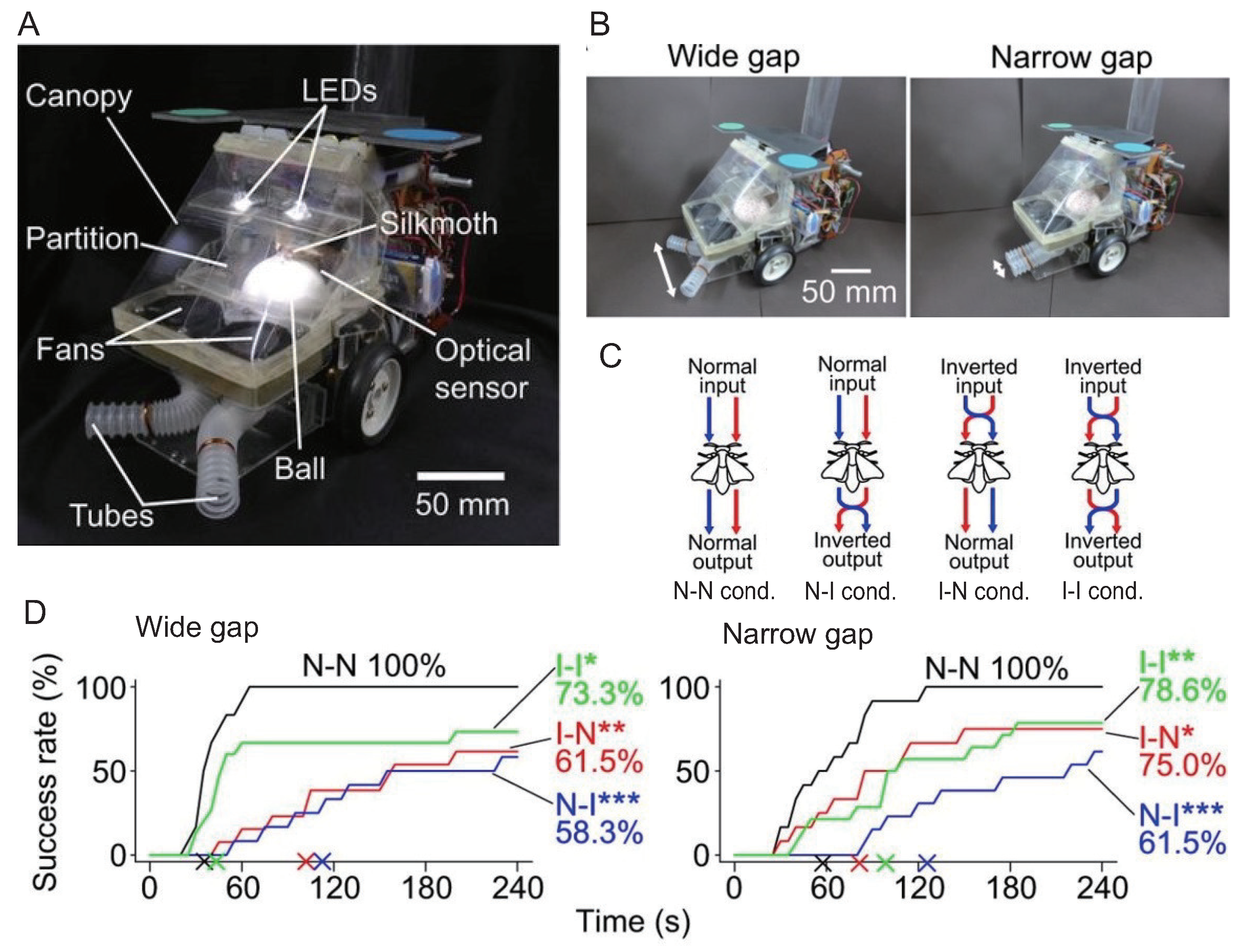
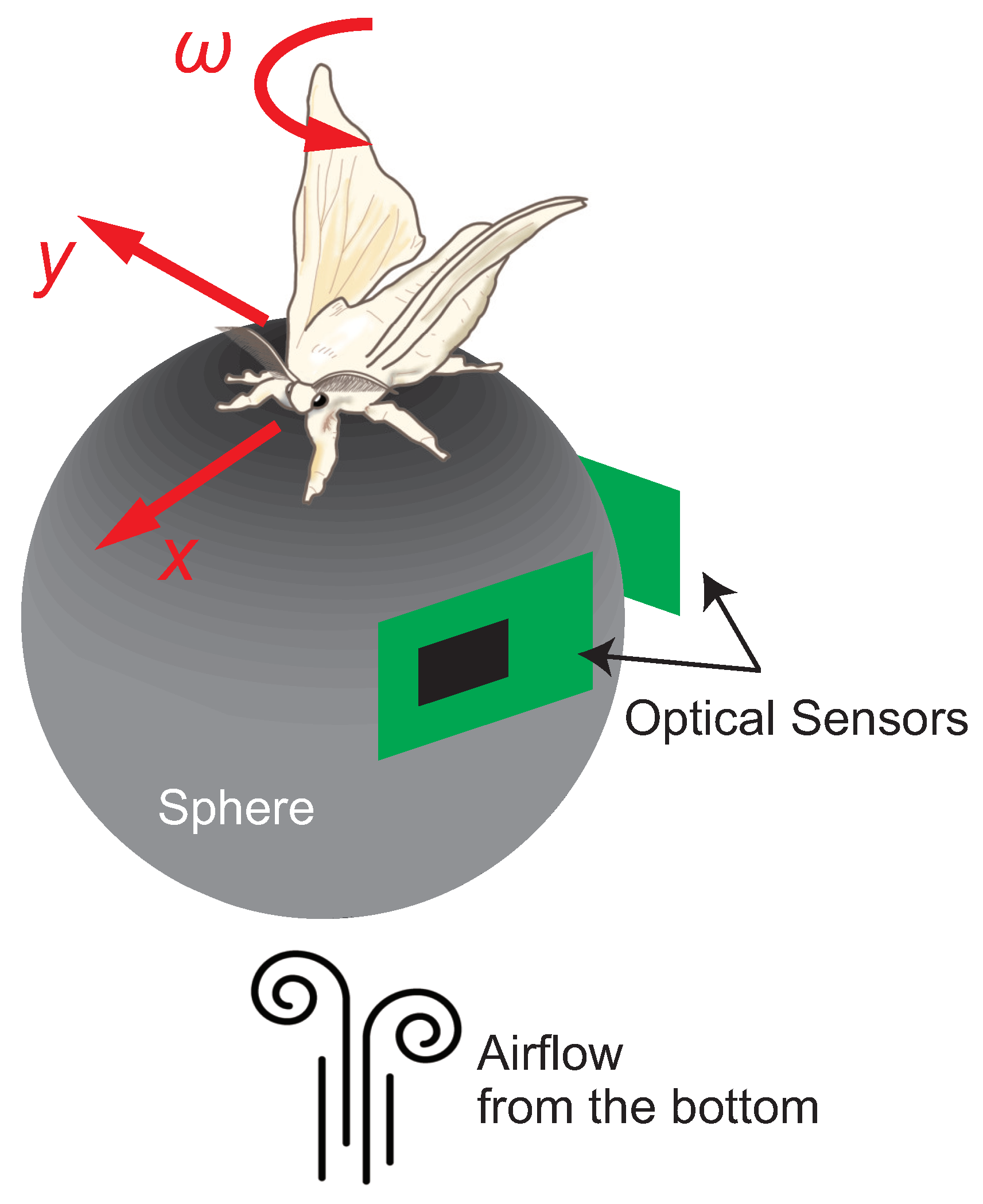
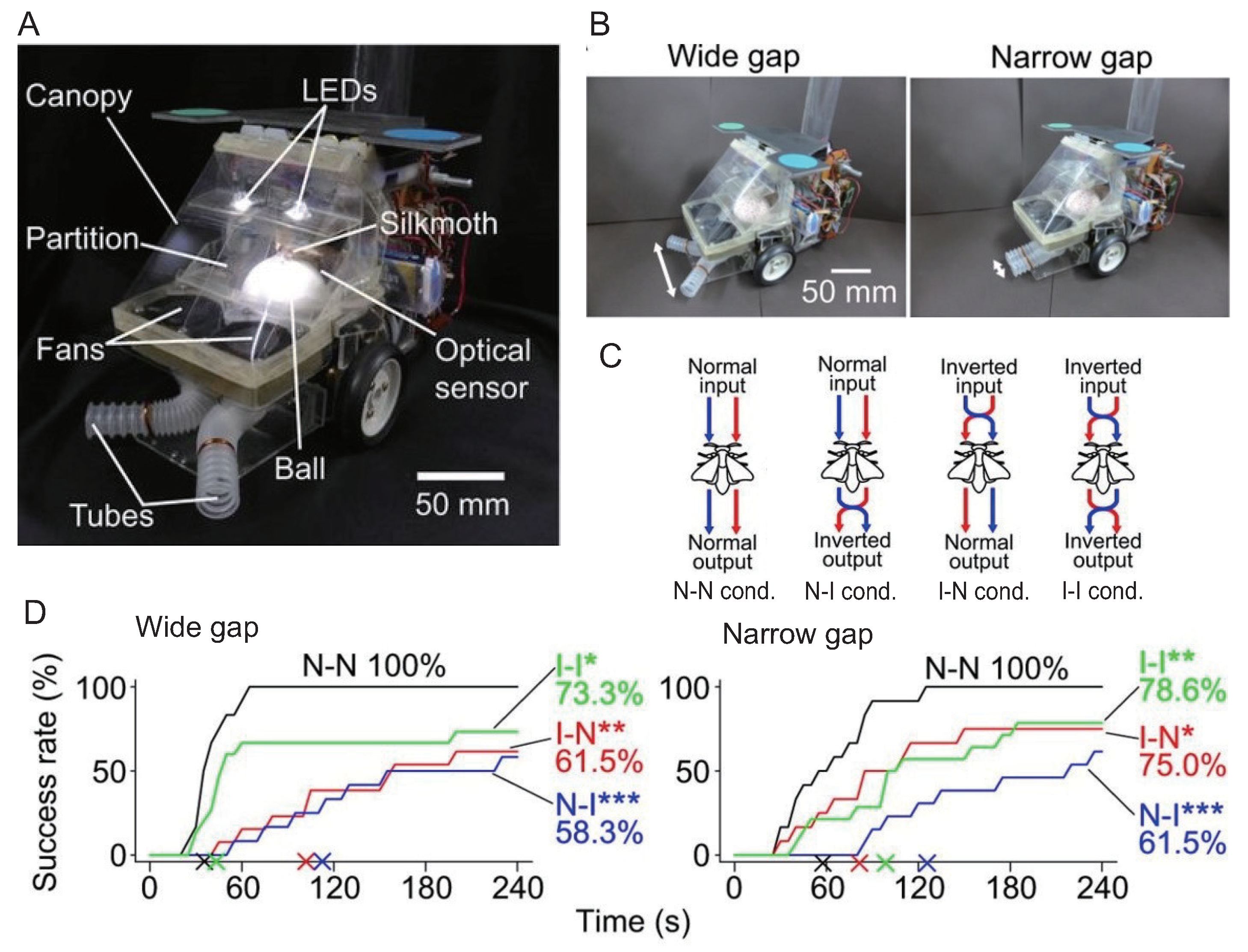
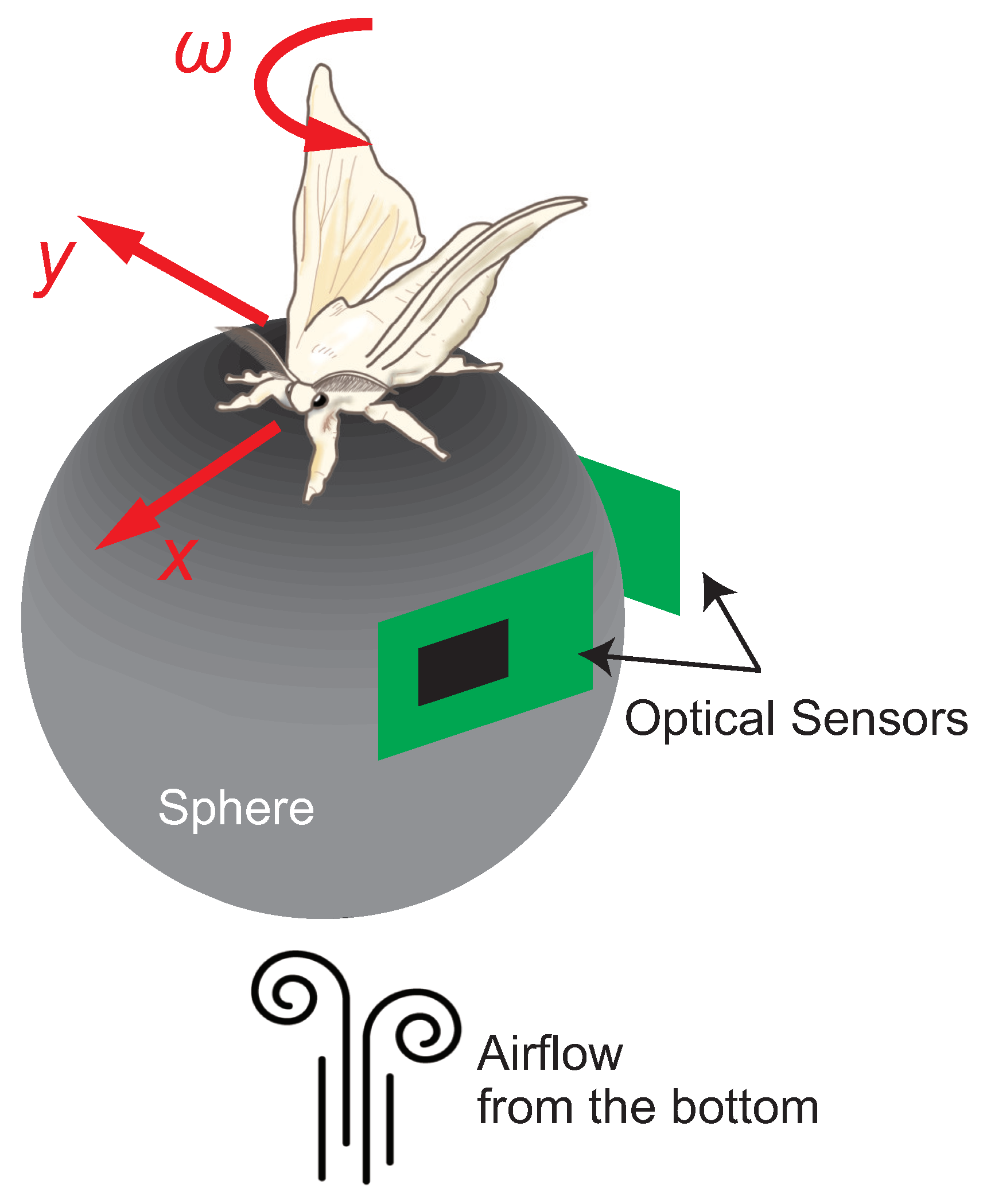
3. Insect On-Board System
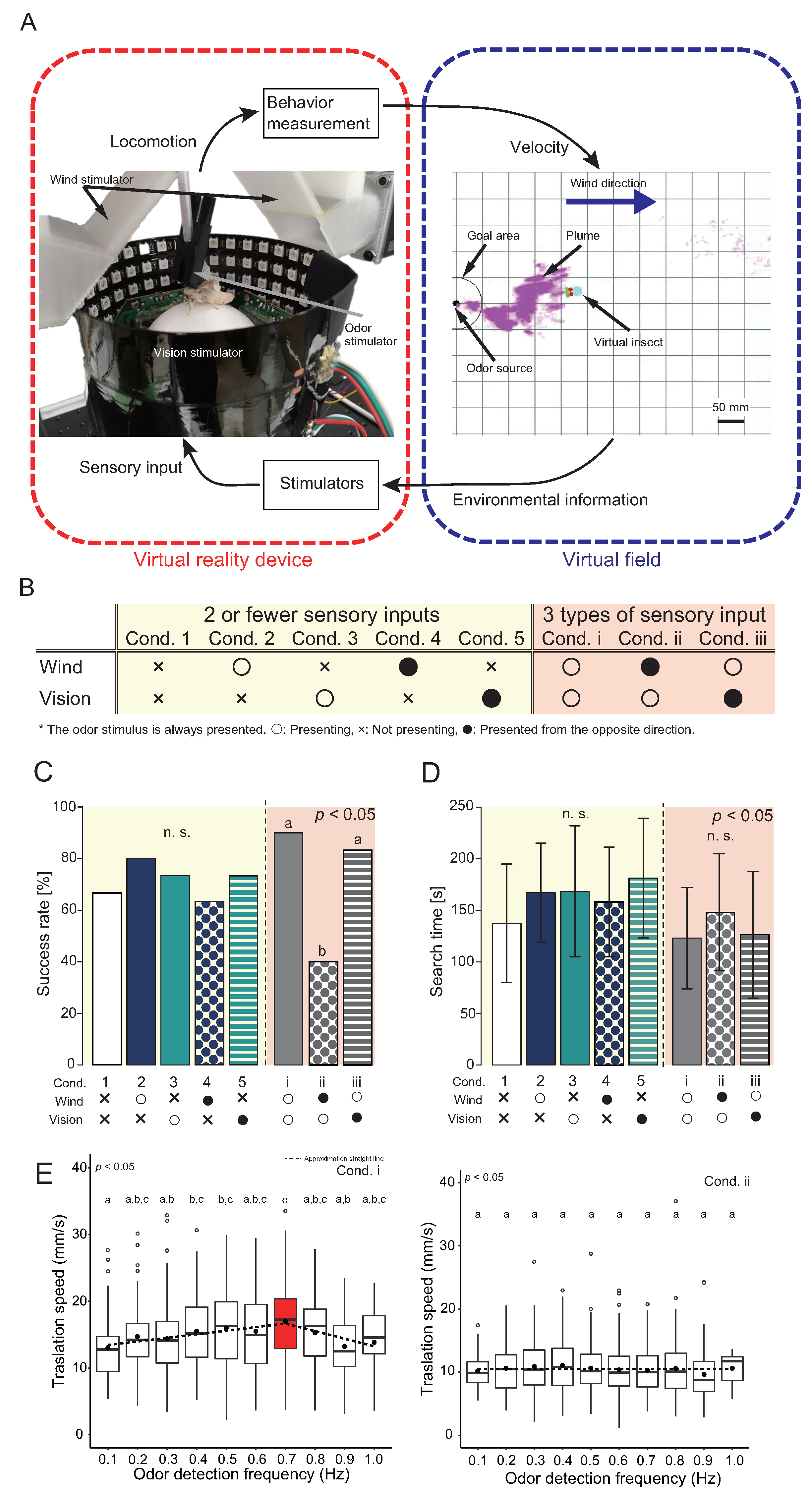
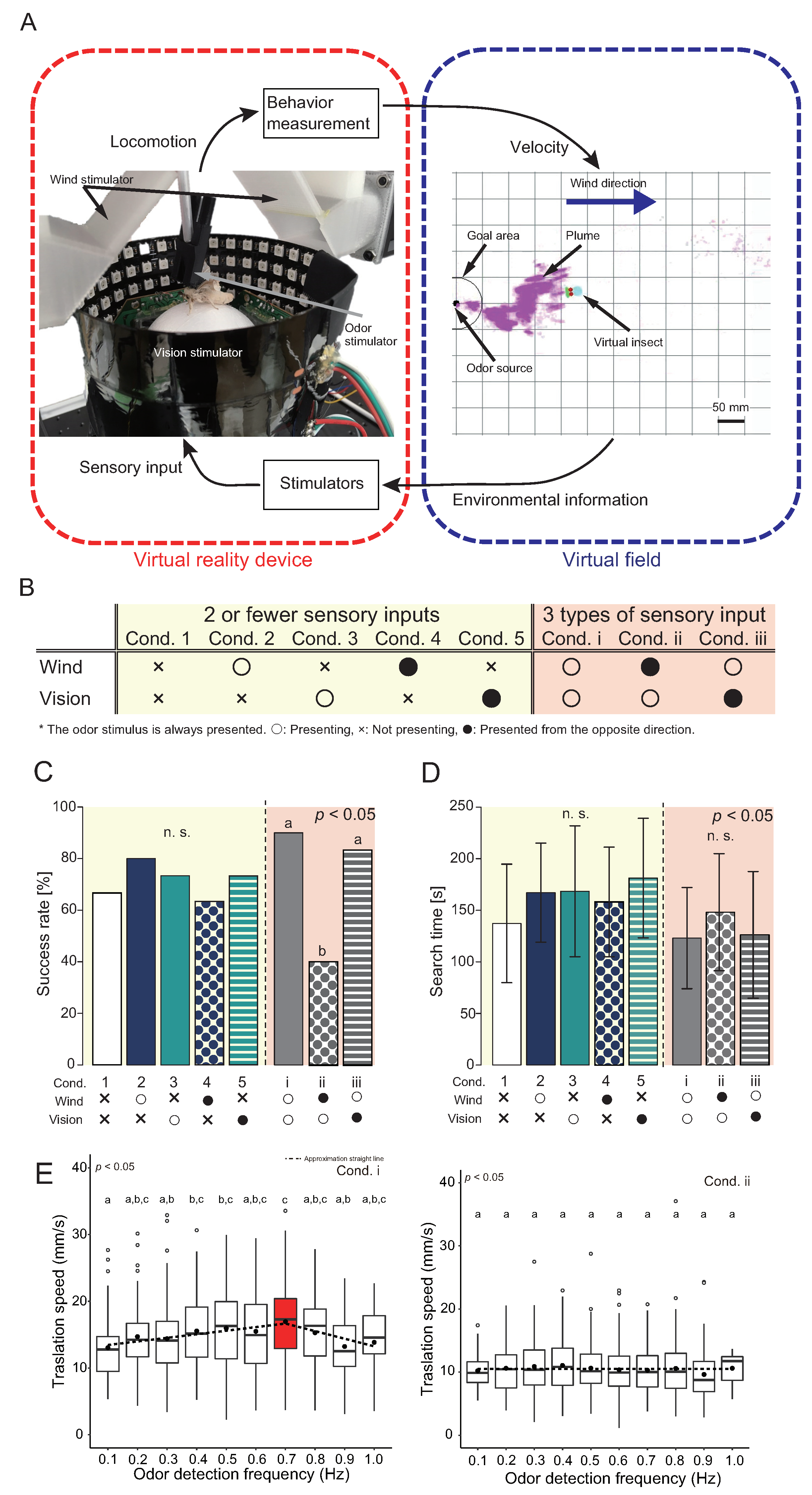
4. Virtual Reality System for an Insect
5. Conclusions and Future Direction
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IFR Presents World Robotics 2021 Reports. Available online: https://ifr.org/ifr-press-releases/news/robot-sales-rise-again (accessed on 16 December 2023).
- Ermolov, I. Industrial Robotics Review. In Robotics: Industry 4.0 Issues & New Intelligent Control Paradigms. Studies in Systems, Decision and Control; Kravets, A., Ed.; Springer Nature: Cham, Switzerland, 2020; Volume 272. [Google Scholar]
- Liu, Z.; Quan, L.; Wenjun, X.; Lihui, W.; Zude, Z. Robot learning towards smart robotic manufacturing: A review. Robot. Comput.-Integr. Manuf. 2022, 77, 102360. [Google Scholar] [CrossRef]
- Robla-Gómez, S.; Becerra, V.M.; Llata, J.R.; Gonzalez-Sarabia, E.; Torre-Ferrero, C.; Perez-Oria, J. Working together: A review on safe human–robot collaboration in industrial environments. IEEE Access 2017, 5, 26754–26773. [Google Scholar] [CrossRef]
- Boada, J.P.; Maestre, B.R.; Genís, C.T. The ethical issues of social assistive robotics: A critical literature review. Technol. Soc. 2021, 67, 101726. [Google Scholar] [CrossRef]
- Calo, M.R.; Froomkin, M.; Kerr, I.R. Robot Law; Edward Elgar Publishing: Northampton, MA, USA, 2016. [Google Scholar]
- Rajan, K.; Saffiotti, A. Towards a science of integrated AI and Robotics. Artif. Intell. 2017, 247, 1–9. [Google Scholar] [CrossRef]
- Kitano, H.; Tambe, M.; Stone, P.; Veloso, M.; Coradeschi, S.; Osawa, E.; Hitoshi, M.; Itsuki, N.; Asada, M. The RoboCup synthetic agent challenge 97. In RoboCup-97: Robot Soccer World Cup I; Springer: Berlin/Heidelberg, Germany, 1998; Volume 1, pp. 62–73. [Google Scholar]
- Buehler, M.; Iagnemma, K.; Singh, S. The DARPA Urban Challenge: Autonomous Vehicles in City Traffic; Springer: Berlin/Heidelberg, Germany, 2009; Volume 56. [Google Scholar]
- Krotkov, E.; Hackett, D.; Jackel, L.; Perschbacher, M.; Pippine, J.; Strauss, J.; Pratt, G.; Orlowski, C. The DARPA robotics challenge finals: Results and perspectives. In The DARPA Robotics Challenge Finals: Humanoid Robots to the Rescue; Springer Nature: Cham, Switzerland, 2018; pp. 1–26. [Google Scholar]
- Rubio, F.; Valero, F.; Llopis-Albert, C. A review of mobile robots: Concepts, methods, theoretical framework, and applications. Int. J. Adv. Robot. Syst. 2019, 16, 1729881419839596. [Google Scholar] [CrossRef]
- Theis, T.N.; Wong, H.S.P. The end of moore’s law: A new beginning for information technology. Comput. Sci. Eng. 2017, 19, 41–50. [Google Scholar] [CrossRef]
- Ahmed, F.; Waqas, M.; Jawed, B.; Soomro, A.M.; Kumar, S.; Hina, A.; Umair, K.; Kim, K.H.; Choi, K.H. Decade of bio-inspired soft robots: A review. Smart Mater. Struct. 2022, 31, 073002. [Google Scholar] [CrossRef]
- Webster-Wood, V.A.; Akkus, O.; Gurkan, U.A.; Chiel, H.J.; Quinn, R.D. Organismal engineering: Toward a robotic taxonomic key for devices using organic materials. Sci. Robot. 2017, 2, eaap9281. [Google Scholar] [CrossRef]
- Prescott, T.J.; Wilson, S.P. Understanding brain functional architecture through robotics. Sci. Robot. 2023, 8, eadg6014. [Google Scholar] [CrossRef]
- Owaki, D.; Ishiguro, A. A quadruped robot exhibiting spontaneous gait transitions from walking to trotting to galloping. Sci. Rep. 2017, 7, 277. [Google Scholar] [CrossRef]
- Shigaki, S.; Yamada, M.; Kurabayashi, D.; Hosoda, K. Robust moth-inspired algorithm for odor source lo-calization using multimodal information. Sensors 2023, 23, 1475. [Google Scholar] [CrossRef]
- Siljak, H.; Nardelli, P.H.; Moioli, R.C. Cyborg Insects: Bug or a Feature? IEEE Access 2022, 10, 49398–49411. [Google Scholar] [CrossRef]
- Alivisatos, A.P.; Chun, M.; Church, G.M.; Greenspan, R.J.; Roukes, M.L.; Yuste, R. The brain activity map project and the challenge of functional connectomics. Neuron 2012, 74, 970–974. [Google Scholar] [CrossRef]
- Stork, N.E. How many species of insects and other terrestrial arthropods are there on Earth? Annu. Rev. Entomol. 2018, 63, 31–45. [Google Scholar] [CrossRef]
- Pfeifer, R.; Scheier, C. Understanding Intelligence; MIT Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Wystrach, A. Movements, embodiment and the emergence of decisions. Insights from insect navigation. Biochem. Biophys. Res. Commun. 2021, 564, 70–77. [Google Scholar] [CrossRef]
- David, I.; Ayali, A. From Motor-Output to Connectivity: An In-Depth Study of in vitro Rhythmic Patterns in the Cockroach Periplaneta americana. Front. Insect Sci. 2021, 1, 655933. [Google Scholar] [CrossRef]
- Roth, E.; Hall, R.W.; Daniel, T.L.; Sponberg, S. Integration of parallel mechanosensory and visual pathways resolved through sensory conflict. Proc. Natl. Acad. Sci. USA 2016, 113, 12832–12837. [Google Scholar] [CrossRef] [PubMed]
- Hughes, G.M. The co-ordination of insect movements: III. Swimming in Dytiscus, Hydrophilus and a dragonfly nymph. J. Exp. Biol. 1958, 35, 567–583. [Google Scholar] [CrossRef]
- Heinze, S. Unraveling the neural basis of insect navigation. Curr. Opin. Insect Sci. 2017, 24, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Weitekamp, C.A.; Libbrecht, R.; Keller, L. Genetics and evolution of social behavior in insects. Annu. Rev. Genet. 2017, 51, 219–239. [Google Scholar] [CrossRef]
- Sakura, M.; Okada, R.; Aonuma, H. Evidence for instantaneous e-vector detection in the honeybee using an associative learning paradigms. Proc. R. Soc. B 2012, 279, 535–542. [Google Scholar] [CrossRef]
- Römer, H. Directional hearing in insects: Biophysical, physiological and ecological challenges. J. Exp. Biol. 2020, 223, jeb203224. [Google Scholar] [CrossRef] [PubMed]
- Merriaux, P.; Dupuis, Y.; Boutteau, R.; Vasseur, P.; Savatier, X. Robust robot localization in a complex oil and gas industrial environment. J. Field Robot. 2018, 35, 213–230. [Google Scholar] [CrossRef]
- Li, W.; Farrell, J.A.; Pang, S.; Arrieta, R.M. Moth-inspired chemical plume tracing on an autonomous underwater vehicle. IEEE Trans. Robot. 2006, 22, 292–307. [Google Scholar] [CrossRef]
- Russell, R.A.; Bab-Hadiashar, A.; Shepherd, R.L.; Wallace, G.G. A comparison of reactive robot chemotaxis algorithms. Robot. Auton. Syst. 2003, 45, 83–97. [Google Scholar] [CrossRef]
- Renou, M. Pheromones and general odor perception in insects. Neurobiol. Chem. Commun. 2014, 1, 23–56. [Google Scholar]
- Kaissling, K.E. Pheromone-controlled anemotaxis in moths. In Orientation and Communication in Arthropods; Birkhauser: Basel, Switzerland, 1997; pp. 343–374. [Google Scholar]
- Willis, M.A.; Avondet, J.L. Odor-modulated orientation in walking male cockroaches Periplaneta americana, and the effects of odor plumes of different structure. J. Exp. Biol. 2005, 208, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, T.D. Pheromones and Animal Behavior: Chemical Signals and Signatures; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Shiota, Y.; Sakurai, T.; Ando, N.; Haupt, S.S.; Mitsuno, H.; Daimon, T.; Kanzaki, R. Pheromone binding protein is involved in temporal olfactory resolution in the silkmoth. Iscience 2021, 24, 103334. [Google Scholar] [CrossRef]
- Mizunami, M.; Yokohari, F.; Takahata, M. Exploration into the adaptive design of the arthropod “microbrain”. Zool. Sci. 1999, 16, 703–709. [Google Scholar] [CrossRef]
- Asama, H. Mobiligence: Emergence of adaptive motor function through interaction among the body, brain and environment. Environment 2007, 1, 3. [Google Scholar]
- Ando, N.; Kanzaki, R. Insect-machine hybrid robot. Curr. Opin. Insect Sci. 2020, 42, 61–69. [Google Scholar] [CrossRef]
- Naik, H.; Bastien, R.; Navab, N.; Couzin, I.D. Animals in Virtual Environments. IEEE Trans. Vis. Comput. Graph. 2020, 26, 2073–2083. [Google Scholar] [CrossRef] [PubMed]
- Kuwana, Y.; Nagasawa, S.; Shimoyama, I.; Kanzaki, R. Synthesis of the pheromone-oriented behaviour of silkworm moths by a mobile robot with moth antennae as pheromone sensors. Biosens. Bioelectron. 1999, 14, 195–202. [Google Scholar] [CrossRef]
- Yamada, N.; Ohashi, H.; Umedachi, T.; Shimizu, M.; Hosoda, K.; Shigaki, S. Dynamic Model Identification for Insect Electroantennogram with Printed Electrode. Sens. Mater. 2021, 33, 4173–4184. [Google Scholar] [CrossRef]
- Martinez, D.; Chaffiol, A.; Voges, N.; Gu, Y.; Anton, S.; Rospars, J.P.; Lucas, P. Multiphasic on/off pheromone signalling in moths as neural correlates of a search strategy. PLoS ONE 2013, 8, e61220. [Google Scholar] [CrossRef] [PubMed]
- Borst, A. Fly visual course control: Behaviour, algorithms and circuits. Nat. Rev. Neurosci. 2014, 15, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Egelhaaf, M.; Kern, R.; Lindemann, J.P. Motion as a source of environmental information: A fresh view on biological motion computation by insect brains. Front. Neural Circuits 2014, 8, 127. [Google Scholar] [CrossRef] [PubMed]
- Ejaz, N.; Krapp, H.G.; Tanaka, R.J. Closed-loop response properties of a visual interneuron involved in fly optomotor control. Front. Neural Circuits 2013, 7, 50. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.V.; Wei, Y.; Krapp, H.G. A biohybrid fly-robot interface system that performs active collision avoidance. Bioinspiration Biomim. 2019, 14, 065001. [Google Scholar] [CrossRef]
- Wittlinger, M.; Wehner, R.; Wolf, H. The desert ant odometer: A stride integrator that accounts for stride length and walking speed. J. Exp. Biol. 2007, 210, 198–207. [Google Scholar] [CrossRef]
- Hertz, G. Cockroach Controlled Robot, Version 1. New York Times, 7 June 2005. [Google Scholar]
- Wessnitzer, J.; Asthenidis, A.; Petrou, G.; Webb, B.A. cricket controlled robot orienting towards a sound source. In Proceedings of the Conference towards Autonomous Robotic Systems, Sheffield, UK, 31 August–2 September 2011; pp. 1–12. [Google Scholar]
- Emoto, S.; Ando, N.; Takahashi, H.; Kanzaki, R. Insect-controlled robot–evaluation of adaptation ability–. J. Robot. Mechatron. 2007, 19, 436–443. [Google Scholar] [CrossRef]
- Ando, N.; Emoto, S.; Kanzaki, R. Odour-tracking capability of a silkmoth driving a mobile robot with turning bias and time delay. Bioinspir. Biomim. 2013, 8, 016008. [Google Scholar] [CrossRef]
- Shirai, K.; Shimamura, K.; Koubara, A.; Shigaki, S.; Fujisawa, R. Development of a behavioral trajectory measurement system (Bucket-ANTAM) for or-ganisms moving in a two-dimensional plane. Artif. Life Robot. 2022, 27, 698–705. [Google Scholar] [CrossRef]
- Kaushik, P.K.; Renz, M.; Olsson, S.B. Characterizing long-range search behavior in Diptera using complex 3D virtual environments. Proc. Natl. Acad. Sci. USA 2020, 117, 12201–12207. [Google Scholar] [CrossRef]
- Ando, N.; Shidara, H.; Hommaru, N.; Ogawa, H. Auditory Virtual Reality for Insect Phonotaxis. J. Robot. Mechatron. 2021, 33, 494–504. [Google Scholar] [CrossRef]
- Radvansky, B.A.; Dombeck, D.A. An olfactory virtual reality system for mice. Nat. Commun. 2018, 9, 839. [Google Scholar] [CrossRef]
- Dahmen, H.J. A simple apparatus to investigate the orientation of walking insects. Experientia 1980, 36, 685–687. [Google Scholar] [CrossRef]
- Obara, Y. Bombyx mori Mating dance: An essential in locating the female. Appl. Entomol. Zool. 1979, 14, 130–132. [Google Scholar] [CrossRef]
- Takasaki, T.; Namiki, S.; Kanzaki, R. Use of bilateral information to determine the walking direction during orientation to a pheromone source in the silkmoth Bombyx mori. J. Comp. Physiol. A 2012, 198, 295–307. [Google Scholar] [CrossRef]
- Ando, N.; Kanzaki, R. A simple behaviour provides accuracy and flexibility in odour plume tracking–the robotic control of sensory-motor coupling in silkmoths. J. Exp. Biol. 2015, 218, 3845–3854. [Google Scholar] [CrossRef]
- Shigaki, S.; Ando, N.; Sakurai, T.; Kurabayashi, D. Analysis of Odor Tracking Performance of Silk moth using a Sensory-Motor Intervention System. Integr. Comp. Biol. 2023, 63, icad055. [Google Scholar] [CrossRef]
- Crapse, T.B.; Sommer, M.A. Corollary discharge across the animal kingdom. Nat. Rev. Neurosci. 2008, 9, 587–600. [Google Scholar] [CrossRef]
- Poulet, J.F.; Hedwig, B. New insights into corollary discharges mediated by identified neural pathways. Trends Neurosci. 2007, 30, 14–21. [Google Scholar] [CrossRef]
- Gatellier, L.; Nagao, T.; Kanzaki, R. Serotonin modifies the sensitivity of the male silkmoth to pheromone. J. Exp. Biol. 2004, 207, 2487–2496. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, T.; Kazawa, T.; Sakurai, T.; Fukushima, R.; Uchino, K.; Yamagata, T.; Namiki, S.; Haupt, S.S.; Kanzaki, R. Odorant concentration differentiator for intermittent olfactory signals. J. Neurosci. 2014, 34, 16581–16593. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Ohashi, H.; Hosoda, K.; Kurabayashi, D.; Shigaki, S. Multisensory-motor integration in olfactory navigation of silkmoth, Bombyx mori, using virtual reality system. eLife 2021, 10, e72001. [Google Scholar] [CrossRef]
- Shigaki, S.; Shiota, Y.; Kurabayashi, D.; Kanzaki, R. Modeling of the adaptive chemical plume tracing algorithm of an insect using fuzzy inference. IEEE Trans. Fuzzy Syst. 2019, 28, 72–84. [Google Scholar] [CrossRef]
- Pansopha, P.; Ando, N.; Kanzaki, R. Dynamic use of optic flow during pheromone tracking by the male silkmoth, Bombyx mori. J. Exp. Biol. 2014, 217, 1811–1820. [Google Scholar] [CrossRef]
- Demir, M.; Kadakia, N.; Anderson, H.D.; Clark, D.A.; Emonet, T. Walking Drosophila navigate complex plumes using stochastic decisions biased by the timing of odor encounters. eLife 2020, 9, e57524. [Google Scholar] [CrossRef]
- Murlis, J.; Elkinton, J.S.; Carde, R.T. Odor plumes and how insects use them. Annu. Rev. Entomol. 1992, 37, 505–532. [Google Scholar] [CrossRef]
- Vickers, N.J.; Baker, T.C. Reiterative responses to single strands of odor promote sustained upwind flight and odor source location by moths. Proc. Natl. Acad. Sci. USA 1994, 91, 5756–5760. [Google Scholar] [CrossRef]
- Rutkowski, A.J.; Quinn, R.D.; Willis, M.A. Three-dimensional characterization of the wind-borne pheromone tracking behavior of male hawkmoths, Manduca sexta. J. Comp. Physiol. A 2009, 195, 39–54. [Google Scholar] [CrossRef] [PubMed]
- Lafon, G.; Howard, S.R.; Paffhausen, B.H.; Avarguès-Weber, A.; Giurfa, M. Motion cues from the background influence associative color learning of honey bees in a virtual-reality scenario. Sci. Rep. 2021, 11, 21127. [Google Scholar] [CrossRef]
- Geng, H.; Lafon, G.; Avarguès-Weber, A.; Buatois, A.; Massou, I.; Giurfa, M. Visual learning in a virtual reality environment upregulates immediate early gene ex-pression in the mushroom bodies of honey bees. Commun. Biol. 2022, 5, 130. [Google Scholar] [CrossRef]
- Shigaki, S.; Fikri, M.R.; Hernandez Reyes, C.; Sakurai, T.; Ando, N.; Kurabayashi, D.; Kanzaki, R.; Sezutsu, H. Animal-in-the-loop system to investigate adaptive behavior. Adv. Robot. 2018, 32, 945–953. [Google Scholar] [CrossRef]
- Shigaki, S.; Minakawa, N.; Yamada, M.; Ohashi, H.; Kurabayashi, D.; Hosoda, K. Animal-in-the-loop System with Multimodal Virtual Reality to Elicit Natural Olfactory Localization Behavior. Sens. Mater. 2021, 33, 4211–4228. [Google Scholar] [CrossRef]
- Minegishi, R.; Takashima, A.; Kurabayashi, D.; Kanzaki, R. Construction of a brain–machine hybrid system to evaluate adaptability of an insect. Robot. Auton. Syst. 2012, 60, 692–699. [Google Scholar] [CrossRef]
- Sponberg, S.; Daniel, T.L. Abdicating power for control: A precision timing strategy to modulate function of flight power muscles. Proc. R. Soc. B Biol. Sci. 2012, 279, 3958–3966. [Google Scholar] [CrossRef]
- Cao, F.; Zhang, C.; Choo, H.Y.; Sato, H. Insect–computer hybrid legged robot with user-adjustable speed, step length and walking gait. J. R. Soc. Interface 2016, 13, 20160060. [Google Scholar] [CrossRef]
- Owaki, D.; Dürr, V.; Schmitz, J. A hierarchical model for external electrical control of an insect, accounting for inter-individual variation of muscle force properties. eLife 2023, 12, e85275. [Google Scholar] [CrossRef]
- Carlsson, M.A.; Hansson, B.S. Dose–response characteristics of glomerular activity in the moth antennal lobe. Chem. Senses 2003, 28, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, M.A.; Chong, K.Y.; Daniels, W.; Hansson, B.S.; Pearce, T.C. Component information is preserved in glomerular responses to binary odor mix-tures in the moth Spodoptera littoralis. Chem. Senses 2007, 32, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Seelig, J.D.; Jayaraman, V. Feature detection and orientation tuning in the Drosophila central complex. Nature 2013, 503, 262–266. [Google Scholar] [CrossRef]
- Mizunami, M.; Okada, R.; Li, Y.; Strausfeld, N.J. Mushroom bodies of the cockroach: Activity and identities of neurons recorded in freely moving animals. J. Comp. Neurol. 1998, 402, 501–519. [Google Scholar] [CrossRef]
- Martin, J.P.; Guo, P.; Mu, L.; Harley, C.M.; Ritzmann, R.E. Central-complex control of movement in the freely walking cockroach. Curr. Biol. 2015, 25, 2795–2803. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shigaki, S.; Ando, N. Usage of a Sensory-Motor Intervention System for Understanding the Adaptive Behavior of Insects. Appl. Sci. 2024, 14, 1139. https://doi.org/10.3390/app14031139
Shigaki S, Ando N. Usage of a Sensory-Motor Intervention System for Understanding the Adaptive Behavior of Insects. Applied Sciences. 2024; 14(3):1139. https://doi.org/10.3390/app14031139
Chicago/Turabian StyleShigaki, Shunsuke, and Noriyasu Ando. 2024. "Usage of a Sensory-Motor Intervention System for Understanding the Adaptive Behavior of Insects" Applied Sciences 14, no. 3: 1139. https://doi.org/10.3390/app14031139
APA StyleShigaki, S., & Ando, N. (2024). Usage of a Sensory-Motor Intervention System for Understanding the Adaptive Behavior of Insects. Applied Sciences, 14(3), 1139. https://doi.org/10.3390/app14031139