
Effects of the Germinated Glycine max and Angelica gigas Nakai Mixture on Hepatic Lipid Metabolism and Bone Turnover Balance in Ovariectomized Rats

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Cell Culture and Treatment
2.2.1. Osteoblasts
2.2.2. Osteoclasts
2.3. Cell Viability Assay
2.4. Alanine Phosphate Activity and Tartrate-Resistant Acid Phosphatase Activity
2.5. Alizarin Red Staining
2.6. Animals and Surgical Procedure
2.7. Micro-Computed Tomography Analysis
2.8. Hematoxylin and Eosin Staining
2.9. ELISA of Serum in Rats
2.10. Western Blot Analyses
2.11. Statistical Analysis
3. Results
3.1. GAM Prevents Weight Gain in the OVX-Induced Rat Model
3.2. GAM Inhibits Lipid Profile Changes in the OVX-Induced Rat Model
3.3. GAM Prevents Liver Damage in the OVX-Induced Rat Model
3.4. GAM Decreases Inflammatory Mediators in the OVX-Induced Rat Model
3.5. GAM Prevents Estrogen Deficiency-Induced Bone Loss in the OVX-Induced Rat Model
3.6. GAM Stimulated Bone Formation in the OVX-Induced Rat Model
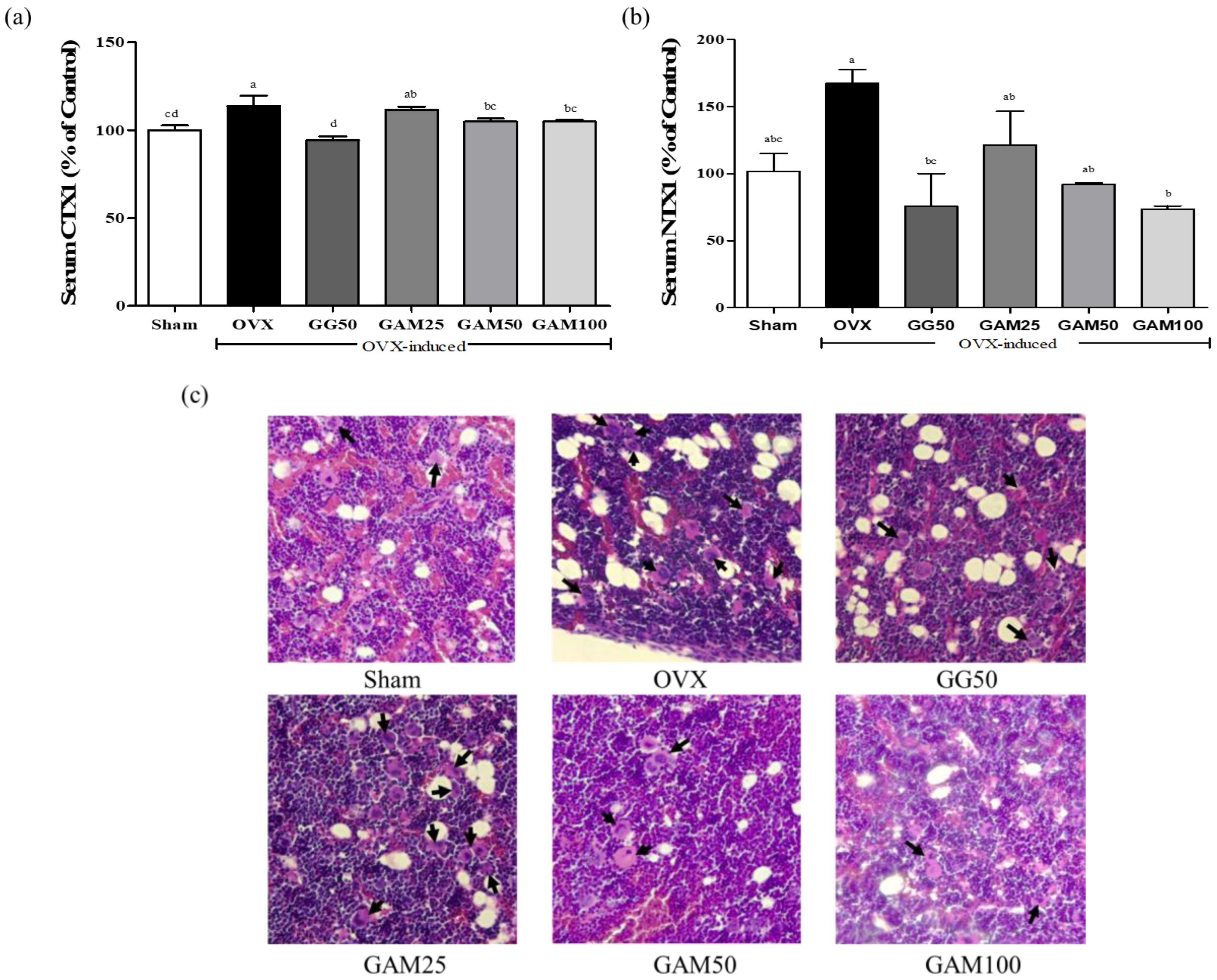
3.7. GAM Inhibits Bone Absorption in the OVX-Induced Rat Model
3.8. GAM Stimulated Osteoblast Differentiation in MC3T3-E1 Cells
3.9. GAM Inhibits TANKL-Induced Osteoclast Differentiation in RAW264.7 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Research on the Menopause in the 1990s: Report of a WHO Scientific Group; WHO: Geneva, Switzerland, 1996.
- Monteleone, P.; Mascagni, G.; Genazzani, A.; Simoncini, T. Symptoms of menopause-global prevalence, physiology and implication. Nat. Rev. Endocrinol. 2018, 14, 199–215. [Google Scholar] [CrossRef]
- Tian, L.; Baek, S.H.; Jang, J.; Kim, S.Y. Imbalanced bone turnover markers and low bone mineral density in patients with osteonecrosis of the femoral head. Int. Orthop. 2018, 42, 1545–1549. [Google Scholar] [CrossRef]
- Ono, T.; Nakashima, T. Recent advances in osteoclast biology. Histochem. Cell Biol. 2018, 149, 325–341. [Google Scholar] [CrossRef]
- Wang, T.; He, C. TNF-α and IL-6: The link between immune and bone system. Curr. Drug Targets 2020, 21, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Song, X.X.; Jin, L.Y.; Li, X.F.; Yu, B.W. Substance P mediates estrogen modulation proinflammatory cytokines release in intervertebral disc. Inflammation 2021, 44, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Song, Q.; Zhang, Y.; Wang, C.; Yang, M.; Zhang, J.; Han, W.; Jiang, P. Antidepressant activity of ω-3 polyunsaturated fatty acids in ovariectomized rats: Role of neuroinflammation and microglial polarization. Lipids Health Dis. 2020, 19, 4. [Google Scholar] [CrossRef] [PubMed]
- Pfeilschifter, J.; Chenu, C.; Bird, A.; Mundy, G.R.; Roodman, G.D. Interleukin-1 and tumor necrosis factor stimulate the formation of human osteoclastlike cells in vitro. J. Bone Miner. Res. 1989, 4, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Bertolini, D.R.; Nedwin, G.E.; Bringman, T.S.; Smith, D.D.; Mundy, G.R. Stimulation of bone resorption and inhibition of bone formation in vitro by human tumour necrosis factors. Nature 1986, 319, 516–518. [Google Scholar] [CrossRef]
- Brennan, O.; Kennedy, O.D.; Lee, T.C.; Rackard, S.M.; O’brien, F.J. Effects of estrgen deficienct and bisphosphonate therapy on osteocyte viability and microdamage accumulation in an ovine model of osteoporosis. J. Orthop. Res. 2011, 29, 419–424. [Google Scholar] [CrossRef]
- Lelovas, P.P.; Xanthos, T.T.; Thoma, S.E.; Lyritis, G.P.; Dontas, I.A. The laboratory rat as an animal model for osteoporosis research. Comp Med. 2008, 58, 424–430. [Google Scholar]
- Cagnacci, A.; Venier, M. The controversial history of hormone replacement therapy. Medicina 2019, 55, 602. [Google Scholar] [CrossRef] [PubMed]
- Tit, D.M.; Bungau, S.; Lovan, C.; Cseppento, D.C.N.; Endres, L.; Sava, C.; Sabau, A.M.; Furau, G.; Furau, C. Effects of the hormone replacement therapy and of soy isoflavones on bone resorption in postmenopause. J. Clin. Med. 2018, 7, 297. [Google Scholar] [CrossRef] [PubMed]
- Gorzkiewicz, J.; Bartosz, G.; Sadowska-Bartosz, I. The potential effects of phytoestrogens: The role in neuroprotection. Molecules 2021, 26, 2954. [Google Scholar] [CrossRef] [PubMed]
- Abdi, F.; Rahnemaei, F.A.; Roozbeh, N.; Pakzad, R. Impact of phytoestrogens on treatment of urogenital menopause symptoms: A systematic review of randomized clinical trials. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 261, 222–235. [Google Scholar] [CrossRef]
- Pan, M.; Li, Z.; Yeung, V.; Xu, R.j. Dietary supplementation of soy germ phytoestrogens or estradiol improves spatial memory performance and increases gene expression of BDNF, TrkB receptor and synaptic factors in ovariectomized rats. Nutr. Metab. 2010, 7, 75. [Google Scholar] [CrossRef]
- Nahas, E.A.P.; Nahas-Neto, J.; Orsatti, F.L.; Carvalho, E.P.; Oliveira, M.L.C.S.; Dias, R. Efficacy and safety of a soy isoflavone extract in postmenopausal women: A randomized, double-blind, and placebo-controlled study. Maturitas 2007, 58, 249–258. [Google Scholar] [CrossRef]
- Chen, K.I.; Erh, M.H.; Su, N.W.; Liu, W.H.; Chou, C.C.; Cheng, K.C. Soyfoods and soybean products: From traditional use to modern applications. Appl. Microbiol. Biotechnol. 2012, 96, 9–22. [Google Scholar] [CrossRef]
- Zhu, D.; Hettiarachchy, N.S.; Horax, R.; Chen, P. Isoflavone contents in germinated soybean seeds. Plant Foods Hum. Nutr. 2005, 60, 147–151. [Google Scholar] [CrossRef]
- Kang, J.W.; Cho, H.E.; Choi, H.M.; Lee, I.C. Anti-wrinkle properties of Angelica gigas Nakai Root extracts using mineral-rich water. J. Cosmet. Dermatol. 2023, 22, 328–334. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, K.M.; Kang, J.S. Preventive effect of Angelica gigas Nakai extract oral administration on dry eye syndrome. Asian Pac. J. Trop Med. 2021, 11, 37–43. [Google Scholar] [CrossRef]
- Ok, S.; Oh, S.R.; Jung, T.S.; Jeon, S.O.; Jung, S.O.; Ryu, D.S. Effects of Angelica gigas Nakai as an Anti-Inflammatory Agent in In Vitro and In Vivo Atopic Dermatitis Models. Evid. Based Complement. Alternat. Med. 2018, 2018, 2450712. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jeong, S.J.; Kwon, H.Y.; Park, S.Y.; Lee, H.J.; Lieske, J.C.; Kim, S.H. Decursin prevents cisplatin-induced apoptosis via the enhancement of antioxidant enzymes in human renal epithelial cells. Biol. Pharm. Bull. 2010, 33, 1279–1284. [Google Scholar] [CrossRef]
- Kouhestani, S.; Jafari, A.; Babaei, P. Kaempferol attenuates cognitive deficit via regulating oxidative stress and neuroinflammation in an ovariectomized rat model of sporadic dementia. Neural Regen. Res. 2018, 10, 1827–1832. [Google Scholar] [CrossRef]
- Lee, G.H.; Hoang, T.H.; Lee, H.Y.; Lim, Y.J.; Kim, J.H.; Jung, S.J.; Chae, S.W.; Rashid, M.M.U.; Chae, H.J.; Yoon, S.J. Ramie leaf Extract Alleviates Bone Loss in Ovariectomized Rats—The Involvement of ROS and Its Associated Signalings. Nutrients 2023, 15, 745. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.H.; Kim, H.S. Menopause-associated lipid metabolic disorders and foods beneficial for postmenopausal women. Nutrients 2020, 12, 202. [Google Scholar] [CrossRef]
- Wu, X.; Kim, M.J.; Yang, H.J.; Park, S. Chitosan alleviated menopausal symptoms and modulated the gut microbiota in estrogen-deficient rats. Eur. J. Nutr. 2021, 60, 1907–1919. [Google Scholar] [CrossRef]
- Nanashima, N.; Horie, K.; Yamanouchi, K.; Tomisawa, T.; Kitajima, M.; Oey, I.; Maeda, H. Blackcurrant (Ribes nigrum) extract prevents dyslipidemia and hepatic steatosis in ovariectomized rats. Nutrients 2020, 12, 1541. [Google Scholar] [CrossRef]
- Targher, G.; Arcaro, G. Non-alcoholic fatty liver disease and increased risk of cardiovascular disease. Atherosclerosis 2007, 191, 235–240. [Google Scholar] [CrossRef]
- Jao, H.Y.; Hsu, J.D.; Lee, Y.R.; Lo, C.S.; Lee, H.J. Mulberry water extract regulates the osteoblast/osteoclast balance in an ovariectomic rat model. Food Funct. 2016, 7, 4753–4763. [Google Scholar] [CrossRef]
- Sapir-Koren, R.; Livshits, G. Postmenopausal osteoporosis in rheumatoid arthritis: The estrogen deficiency-immune mechanisms link. Bone 2017, 103, 102–115. [Google Scholar] [CrossRef]
- Udagawa, N.; Koide, M.; Nakamura, M.; Nakamichi, Y.; Yamashita, T.; Uehara, S.; Kobayashi, Y.; Guruya, Y.; Yasuda, H.; Fukada, C.; et al. Osteoclast differentiation by RANKL and OPG signaling pathways. J. Bone Miner. Metab. 2021, 39, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Pandijaitan, N.; Hettiarachchy, N.; Ju, Z.Y.; Crandall, P.; Sneller, C.; Dombek, D. Evaluation of genistin and genistein contents in soybean varieties and soy protein concentrate prepared with 3 basic methods. J. Food Sci. 2000, 65, 399–402. [Google Scholar] [CrossRef]
- McGrain, A.K.; Chen, J.C.; Wilson, K.A.; Tan-Wilson, A.L. Degradation of trypsin inhibitors during soybean germination. Phytochemistry 1989, 28, 1013–1017. [Google Scholar] [CrossRef]
- Sattar, A.; Neelofar; Akhtar, M.A. Irradiation and germination effects on phytate, protein and amino acids of soybean. Plant Food Hum. Nutr. 1990, 40, 185–194. [Google Scholar] [CrossRef]
- Rhy, S.B.; Zheng, L.; Wang, X. Changes in phospholipase D experessiopn in soybeans during seed development and germination. J. Am. Oil Chem. Soc. 1996, 73, 1171–1176. [Google Scholar]
- Ohanenye, I.C.; Tsopmo, A.; Ejike, C.E.C.C.; Udenigwe, C.C. Germination as bioprocess for enhancing the quality and nutritional prospects of legume proteins. Trands Food Sci. Technol. 2022, 101, 213–222. [Google Scholar] [CrossRef]
- Choi, C.W.; Choi, S.W.; Kim, H.J.; Lee, K.S.; Kim, S.H.; Kim, S.L.; Do, S.H.; Seo, W.D. Germinated soy germ with increased soyasaponin Ab improves BMP-2-induced bone formation and protects against in vivo bone loss in osteoporosis. Sci. Rep. 2018, 8, 12970. [Google Scholar] [CrossRef]
- Feresin, R.G.; Johnson, S.A.; Elam, M.L.; Jo, E.; Arjmandi, B.H.; Hakkak, R. Effects of obesity on bone mass and quality in ovariectomized female Zucker rats. J. Obes. 2014, 2014, 690123. [Google Scholar] [CrossRef]
- Dalvand, S.; Gheshlagh, R.G.; Kurdi, A. Prevalence of poststroke depression in Iranian patients: A systematic review and meta-analysis. Neuropsychiatr. Dis. Treat. 2018, 14, 3073–3080. [Google Scholar] [CrossRef] [PubMed]
- Orekhov, A.N.; Myasoedova, V.A. Low density lipoprotein-induced lipid accumulation is a key phenomenon of atherogenesis at the arterial cell level. Vessel Plus 2019, 3, 3. [Google Scholar] [CrossRef]
- Kang, S.J.; Choi, B.R.; Kim, S.H.; Yi, H.Y.; Park, H.R.; Song, C.H.; Ku, S.K.; Lee, Y.J. Anti-climacterium effects of pomegranate concentrated solutions in ovariectomized ddY mice. Exp. Ther. Med. 2017, 13, 1249–1266. [Google Scholar] [CrossRef]
- Yadav, S.; Jangra, R.; Sharma, B.R.; Sharma, M. Current Advancement in Biosensing techniques for determination of Alanine aminotransferase and Aspartate aminotransferase-a Mini Review. Process. Biochem. 2022, 114, 71–76. [Google Scholar] [CrossRef]
- Kameda, T.; Mano, H.; Yuasa, T.; Mori, Y.; Miyazawa, K.; Shiokawa, M.; Nakamaru, Y.; Hiroi, E.; Hiura, K.; Kameda, A.; et al. Estrogen Inhibits Bone Resorption by Directly Inducing Apoptosis of the Bone-resorbing Osteoclasts. J. Exp. Med. 1997, 186, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Lim, E.; Yeo, S.; Yong, Y.; Yang, J.; Jeong, S.Y. Anti-Menopausal effects of Cornus officinalis and Ribes fasciculatum extract in vitro and in vivo. Nutrients 2020, 12, 369. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.W.; Ma, B.; Zi, Y.; Xiang, L.B.; Han, T.Y. Effects of rutin on osteoblast MC3T3-E1 differentiation, ALP activity and Runx2 protein expression. Eur. J. Histochem. 2021, 65, 3195. [Google Scholar] [CrossRef]
- Mukaiyama, K.; Kamimura, M.; Uchiyama, S.; Ikegami, S.; Nakamura, Y.; Kato, H. Elevation of serum alkaline phosphatase (ALP) level in postmenopausal women is caused by high bone turnover. Aging Clin. Exp. Res. 2015, 27, 413–418. [Google Scholar] [CrossRef]
- Lee, H.H.; Jang, J.W.; Lee, J.K.; Park, C.K. Rutin improves bone histomorphometric values by reduction of osteoclastic activity in osteoporosis mouse model induced by bilateral ovariectomy. J. Korean Neurosurg. Soc. 2020, 63, 433–443. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sham | OVX | GG50 | GAM25 | GAM50 | GAM100 | |
|---|---|---|---|---|---|---|
| Initial body weight (g) | 174.5 ± 9.0 NS | 170.7 ± 8.8 | 173.5 ± 8.3 | 171.6 ± 8.1 | 171.9 ± 8.2 | 171.1 ± 8.9 |
| Final body weight (g) | 241.4 ± 5.7 b | 274.0 ± 8.7 a | 260.6 ± 8.6 ab | 266.4 ± 5.4 ab | 265.8 ± 3.9 ab | 249.7 ± 6.3 ab |
| Organs (% of body weight) | ||||||
| Liver (%) | 3.5 ± 0.4 NS | 3.5 ± 0.4 | 3.7 ± 0.4 | 3.7 ± 0.4 | 3.6 ± 0.2 | 3.9 ± 0.3 |
| Uterus (%) | 0.26 ± 0.03 a | 0.08 ± 0.07 b | 0.1 ± 0.09 b | 0.05 ± 0.01 b | 0.06 ± 0.06 b | 0.08 ± 0.05 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Cho, Y.; Choi, J.; Choi, H.; Jung, T.S.; Kwon, J. Effects of the Germinated Glycine max and Angelica gigas Nakai Mixture on Hepatic Lipid Metabolism and Bone Turnover Balance in Ovariectomized Rats. Appl. Sci. 2023, 13, 5084. https://doi.org/10.3390/app13085084
Lee S, Cho Y, Choi J, Choi H, Jung TS, Kwon J. Effects of the Germinated Glycine max and Angelica gigas Nakai Mixture on Hepatic Lipid Metabolism and Bone Turnover Balance in Ovariectomized Rats. Applied Sciences. 2023; 13(8):5084. https://doi.org/10.3390/app13085084
Chicago/Turabian StyleLee, Sangmin, Yunsang Cho, Jihye Choi, Hwal Choi, Tae Sung Jung, and Jungkee Kwon. 2023. "Effects of the Germinated Glycine max and Angelica gigas Nakai Mixture on Hepatic Lipid Metabolism and Bone Turnover Balance in Ovariectomized Rats" Applied Sciences 13, no. 8: 5084. https://doi.org/10.3390/app13085084
APA StyleLee, S., Cho, Y., Choi, J., Choi, H., Jung, T. S., & Kwon, J. (2023). Effects of the Germinated Glycine max and Angelica gigas Nakai Mixture on Hepatic Lipid Metabolism and Bone Turnover Balance in Ovariectomized Rats. Applied Sciences, 13(8), 5084. https://doi.org/10.3390/app13085084