Abstract
Alkylating agents such as cyclophosphamide (CPA) are commonly used in cytotoxic or immunosuppressive therapies for different types of diseases. One of the main secondary effects of such therapy is the reduction in follicular reserve, targeting the primordial follicles. The aim of this study was to investigate the antioxidant and protective effects of Plantago sempervirens extract on the follicular pool. The experiment was performed on Wistar female rats, for 21 days. They were divided into five groups according to the treatment they received (Control, CPA (200 mg/kg BW), P. sempervirens extract in three different doses—25/50/100 dw/mL ethanol extract (5 mL/kg BW)). The investigations regarded enzymatic and non-enzymatic nitro-oxidative stress, hormone levels (FSH and estrogen), TEM sections of the ovaries, and oestrus cycle monitorization. CPA strongly increased the oxidative stress (TOS—32.8 mmol H2O2/L; NO—68.60 μmol/L; OSI—6.82), alongside a depletion of FSH and a blockage of the oestrus cycle in the metestrus phase. The follicular pool was strongly depleted during CPA exposure, but FSH concentration and the TEM images of the cells revealed a significant improvement after treatment, as the dose increases (P3). Therefore, these findings reveal that P. sempervirens therapy could have a potential protective effect against CPA-induced acute damages in the ovaries, as well as maintaining a functional oestrus cycle.
1. Introduction
Chemotherapy is still the most common treatment against cancer, both for humans and animals [1,2]. Even if chemotherapeutic drugs have so many side effects that affect healthy cells, such therapies are the most effective. Chemotherapy-induced ovarian damage is the foremost cause of ovarian dysfunction, early menopause, primary ovarian insufficiency, or even loss of fertility. Therefore, scientists are still searching for alternatives that could be used for alleviating the inherent side effects [3,4,5].
Cyclophosphamide (CPA) is an alkylating agent, well-known for its gonadotoxic activity, that induces cross-links into DNA structure leading the follicular cells into apoptotic pathways [6]. In addition, reactive oxygen species (ROS) and reactive nitrogen species (RNS) are produced, which are harmful for the ovarian cells [7]. In addition, CPA activates the P13K/PTEN/Akt/mTOR pathway that leads to “the burnout effect”, which means an overactivation of the primordial follicles associated with their massive recruitment into the growing pool of follicles [8,9].
Phytotherapy has been employed multiple times during chemotherapy, with the aim of preventing or reducing the expected side effects [10,11]. From thousands of plants and numerous phytochemicals that have been investigated over the years, a particularly attractive approach in this field is to find a proper chemical formulation that could preserve fertility. The ovarian protective effect of Plantago species has not been previously investigated. According to previous studies, Plantago extracts, taken from leaves, exhibit various properties, including anticarcinogenic, anti-inflammatory, anti-hypertensive, weakly antibiotic, wound healing and other pharmacological activities [12,13,14]. Our previous studies suggest that Plantago species, such as P. major, P. lanceolata, P. cornuti, P. media, and P. sempervirens, follow a common chemical composition, rich in luteolin, apigenin, and rutin, but each of the extracts has a particular chemical pattern. With respect to their antioxidant activity, P. sempervirens stands out as being an antioxidant six times stronger than its Plantago counterparts [15,16].
Hence, this paper was designed to further evaluate the effects of P. sempervirens extract in a model of ovarian failure due to chemotherapy. The main hypothesis is based on the potential of P. sempervirens to maintain the ovarian follicles at resting stages, which are known to be more resistant to chemotherapy drugs [9]. Taking into consideration the chemical composition rich in phenolic compounds with estrogen-like activity, we aimed to investigate the protective effects of P. sempervirens at different doses on nitro-oxidative stress, oestrus cycles, FSH levels, and cell structural changes in the rat ovaries.
Our study highlights the potential protective effect of P. sempervirens alcoholic extract on the ovary oestrous cycle, histology (ultrastructure), hormones, and stress markers after the induced cytotoxic effects of CPA, in a rat model. This protective effect of P. sempervirens has not been previously demonstrated. The study covers three increasing concentrations (25/50/100 mg/kg BW) of the P. sempervirens extract and it highlights the fact that dose-dependent studies are important. This calls for more in depth studies regarding the investigation of active compounds and synergistic effects of the extract and requires more data from rat groups, larger mammals, or volunteering patients.
2. Materials and Methods
2.1. Plant Material and Extract Preparation
P. sempervirens leaves were collected from the Botanical Garden “Alexandru Borza”, Cluj-Napoca, Romania. A plant specimen was deposited at the Botanical Garden’s Herbarium (CL663373). The extract was prepared using a method described previously [15]. In short, 1 cm fragments of fresh leaves were percolated (at 21 °C, for 3 days) with 70% ethanol obtaining a 1 to 1.8 (w:v/g:mL, 55.5% extract) of P. sempervirens. In addition, the chemical composition of the extract was published in advance [15], identifying nine polyphenols including apigenin (2.34 μg/mL), rutin (2.78 μg/mL), luteolin (1.02 μg/mL), ferulic acid (0.45 μg/mL), and p-coumaric acid (0.68 μg/mL).
2.2. Chemicals
The reagents included in standard assay kits with ELISA methods (3-Nitrotirosine, Glutathione peroxidase, Estrogen, and Follicle-stimulating hormone) were obtained from SunRed Biotechnology, Shanghai, China and Elabscience, Houston, TX, USA. The reagents for colorimetric and kinetic methods (thiobarbituric acid, vanadium (III) chloride (VCl3), methanol, diethylether, xylenol orange, hydrogen peroxide, [o-cresosulfonphtalein-3,3-bis (sodium methyliminodiacetate)], ortho-dianisidinedihydrochloride (3,3′-dimethoxybenzidine), ferrous ammonium sulfate, sulphuric acid, hydrogencholride acid, glycerol, trichloracetic acid (TCA)) were obtained from Merck & Co., Bucharest, Romania. Other chemicals and solvents used in the study were of analytical grade.
2.3. Animals
The experiments were performed on female Wistar rats, weighing 150 ± 70 g. The animals were accommodated at the Animal Facility of the “Iuliu Hațieganu” University of Medicine and Pharmacy, Cluj-Napoca, Romania, in standard polypropylene cages and standard laboratory conditions (12 h light/dark cycle, temperature 25 ± 1 °C, relative humidity 55 ± 5%, water and food ad libitum). All the procedures described in the current paper comply with the Directive 2010/63/EU and Romanian national legislation (law no. 43/2014). The project was approved by the Institutional Animal Ethical Committee of “Iuliu Hațieganu” University of Medicine and Pharmacy, Cluj-Napoca, Romania (22/13.12.2016).
2.4. Study Design
Female rats were randomly divided into five groups (n = 6): control group (C), cyclophosphamide (CPA) group—a single intraperitoneal injection with 200 mg/kg BW CPA in the 15th day of the experiment, CPA+ P1 (25 mg dw/mL P. sempervirens extract—5 mL/kg BW), CPA+ P2 (50 mg dw/mL P. sempervirens extract—5 mL/kg BW), CPA+ P3 (100 mg dw/mL P. sempervirens extract—5 mL/kg BW). The extract was orally administered, daily, for 21 days. The animals were euthanized in the 21st day, by cervical dislocation, under deep narcosis with 4% isoflurane.
2.5. Measurement of Nitro-Oxidative Stress
The nitro-oxidative status of the serum was assessed using several markers, such as total antioxidant capacity (TAC), total oxidative stress (TOS), oxidative stress index (OSI), malondialdehyde (MDA), catalase (CAT), glutathione peroxidase (GPX), nitric oxide (NO), 3-nitrotirosine (3-NT), and thiol groups (SH). The first step was to remove the potential contaminant proteins by extraction with a 3:1 (v:v) solution of methanol: diethyl ether (1:9, v:v) [17]. TOS and TAC were assessed using colorimetric methods [18]. OSI represents the ratio between TOS/TAC, meaning the oxidative stress level. The NO was evaluated using the Griess reaction [19]. 3-NT and GPX were investigated via the ELISA method, using an ELISA kit mentioned above. Lipid peroxidation byproducts were assessed by MDA method [20]. CAT was determined by a method previously described [21]. The total thiols were evaluated by measuring reduced glutathione [20]. The nitro-oxidative measurements were taken at the end of the experiment.
2.6. Measurement of Sex Hormones
Both Estrogen (E2) and Follicle-stimulating hormone (FSH) were assessed using Enzyme-linked immunosorbent assay (ELISA) method anti-rat, purchased from Elabscience, Wuhan, China.
2.7. Oestrus Cycles Monitoring
Before evaluating the daily stages of the oestrus cycle, the female rats were housed with male cage beddings, in order to align them in the oestrous stage. Therefore, vaginal smears of the female rats were performed daily, starting from day 4th of the experiment, by delicately flushing the vagina with a small amount (20 μL) of phosphate buffer saline (PBS). The identification of each stage of the oestrus cycle was described using vaginal cytology according to Byers recommendations [22]. Hence, the pro-oestrus stage (P) is easily identified by the presence of nucleated cells and several cornified epithelial cells. The oestrus stage (E) is characterized by a considerable number of cornified epithelial cells. The metoestrus stage (M) and the dioestrus stage (D) are sometimes difficult to distinguish due to the presence of leukocytes, nucleated cells, and cornified epithelial cells, but usually dioestrus has a higher proportion of leukocytes. The cycle repeats at 4/5 days.
2.8. Transmission Electron Microscopy of the Ovarian Tissue
Tissue samples were fixed with 2.7% glutaraldehyde solution (Electron Microscopy Sciences, Hatfield, UK) in 0.1 M phosphate buffer, at 4 °C, followed by washing with 0.15 M PBS and a second fixation with 2% osmic acid solution. Samples were dehydrated with acetone and sections were embedded in epoxy resin (Epon 812). Samples were cut into 60 nm thick sections, using a Leica UC6 ultramicrotome (Leica Microsystems GmbH, Wetzlar, Germany) and were contrasted with uranyl acetate and lead citrate (Merck and CoBucharest, Romania) as described in [23]. The analysis was performed with a Jeol JEM 1010 (Tokyo, Japan) transmission electron microscope, coupled with a Mega View III CCD camera for image capturing (Olympus, Soft Imaging System, Münster, Germany).
2.9. Statistical Analysis
All data are expressed as mean ± SEM (standard error of mean). The Gaussian distribution was verified by Shapiro-Wilk normality test. One-way analysis of variance ANOVA, followed by Bonferroni’s Multiple Comparison test procedure, was performed. Statistical significance was at p < 0.05 (95% confidence interval). Statistical values were obtained using GraphPad Prism, version 5.0 for Windows, GraphPad Software, San Diego, CA, USA.
3. Results
3.1. Nitro-Oxidative Stress Status
Reactive oxygen species (ROS) synthesis was evaluated by TOS levels, OSI and MDA concentration. As shown in Figure 1, experimental gonadotoxicity induced significant oxidative stress, by increasing TOS (p < 0.05), OSI (p < 0.05), and MDA (p < 0.05) as compared with the control group. The inhibitory effects were most effective for the second dose, P2 (50 mg dw/mL), both for TOS (p < 0.05) and OSI (p < 0.001) and for dose P3 (100 mg dw/mL), the oxidative stress level was significantly reduced (p < 0.05) (Figure 1c). Regarding the MDA concentrations after P. sempervirens pretreatment, in Figure 1d there is an obvious decrease in the peroxidation byproducts, but our statistical tests could not confirm this, probably because of the small number of animals used per group.
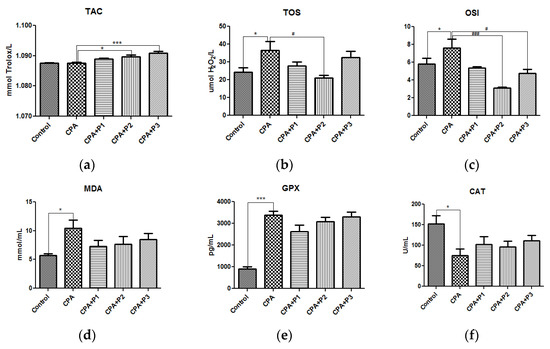
Figure 1.
The in vivo antioxidant response of P. sempervirens extract seen in (a) TAC, (b) TOS, (c) OSI, (d) MDA levels, (e) GPX and (f) CAT activities in serum of Control, CPA, and CPA + P1/P2/P3 exposed animals. Data represent mean ± SEM; One-way ANOVA followed by Bonferroni’s Multiple comparison test; * Significant at p < 0.05; *** Significant at p < 0.001—compared with Control group; # Significant at p < 0.05; ### Significant at p < 0.001—compared with CPA group.
The antioxidant capacity (TAC), the antioxidant enzymes (GPx and CAT) as shown in Figure 1a,d,e, and SH levels (Figure 2a) are the most reliable parameters for evaluating the antioxidant defense mechanism in an in vivo experimental model. Therefore, a single dose of chemotherapy (200 mg/kg BW) is enough to activate the antioxidant barriers as seen is GPx activity (p < 0.05). On the other side, CAT was significantly depleted (p < 0.05), as well as SH groups (p < 0.05). P. sempervirens, in turn, was able to lessen the harmful effects by returning the enzymatic activities to near-normal and increasing the glutathione level to greater values than control, in a dose dependent manner (p < 0.001) (Figure 2a).
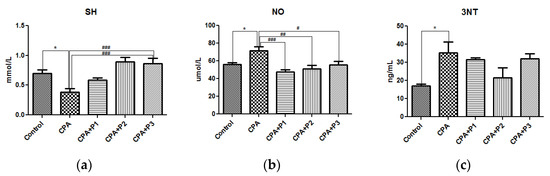
Figure 2.
(a) The SH concentration, (b) NO and (c) 3NT levels in serum of Control, CPA and CPA + P1/P2/P3 exposed animals. Data represent mean ± SEM; One-way ANOVA followed by Bonferroni’s Multiple comparison test; * Significant at p < 0.05—compared with Control group; # Significant at p < 0.05; ## Significant at p < 0.01; ### Significant at p < 0.001—compared with CPA group.
ROS react with nitrogen oxide (NO) creating reactive nitrogen species (RNS), which are known for being strong oxidants [24]. Indirectly, the RNS level was measured by determining the concentrations of NO and 3NT, as seen in Figure 2b,c. Therefore, both parameters were significantly increased (p < 0.05) following CPA administration. The P. sempervirens pretreatment significantly reduced the NO levels (p < 0.001), (p < 0.01) and (p < 0.05) in an inversely proportional mode with respect to the dose.
3.2. Sex Hormone Levels
The estradiol and FSH concentrations in control and experimental groups are shown in Figure 3a,b. The CPA group is represented, in both graphs, with the lowest values, with a p < 0.05 in FSH graph, as compared to control group. The three doses of the extract (P1/P2/P3) have shown, in a dose dependent manner, their efficacy in rebalancing the sex hormones to normal or greater values, as seen in Figure 3a—CPA + P3, p < 0.001. These observations are correlated with the oestrus cycle dynamics, illustrated in Figure 4. After CPA administration on the 15th day of the experiment, the oestrus cycle remained stuck in the metoestrus stage. The administration of the P. sempervirens extract prior to CPA injection and five days after it, the oestrus cycle was visible in a continuous dynamic, following one or two stages more after CPA injection.
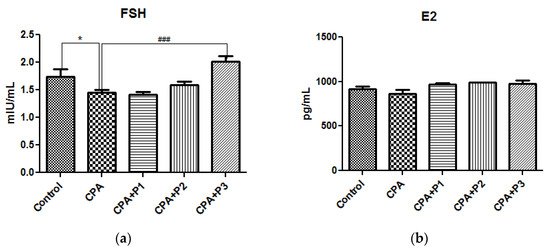
Figure 3.
Sex hormone levels in serum—(a) FSH and (b) E2 of Control, CPA, and CPA + P1/P2/P3 exposed animals. Data represent mean ± SEM; One-way ANOVA followed by Bonferroni’s Multiple comparison test; * Significant at p < 0.05—compared with Control group; ### p < 0.001—compared with CPA group.
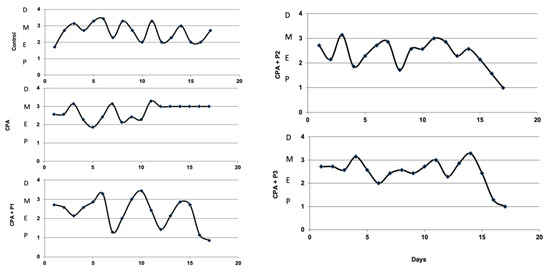
Figure 4.
The oestrus stages determined by vaginal cytology (pro-oestrus (P), oestrus (E), metoestrus (M) and dioestrus (D)) in Control, CPA, and CPA + P1/P2/P3 groups. The oestrus cycle respects a repetitive dynamics at almost every five days, except the CPA group which blocking in the metoestrus stages is evident after CPA administration.
3.3. Ultrastructure of the Ovarian Cell
The ultra-thin sections investigated by TEM support the previous biochemical observations, by which the CPA administration to experimental animals induced significant injuries. As seen in Figure 5B, the ovary exposed to CPA was poorly populated with lipidic granules, which are prominent in control animals (Figure 5A). The untreated samples showed several layers of theca cells, which are elongated and smaller as compared with the granulosa cells. The latter ones are arranged concentrically, most of them with a round nucleus and possessing many electron dense granules, that most likely represent hormone secretions. Interestingly, the extract pretreatment could prevent the hormone deprivation, as seen in Figure 5C–E with a focus on the P2 and P3 doses, that could clearly increase the number and the electron-density of the granules (P2) or only the number of the granules (P3). Besides, the structure of the granulosa cells appeared to be very well preserved after P2 and P3 doses. As the dose of the extract increases (Figure 5E) the perfectly round-shaped nuclei are more prominent.
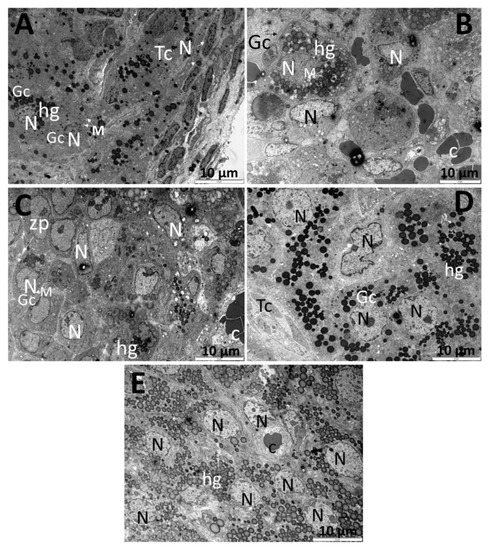
Figure 5.
Transmission electron microscopy images (10 μm) of the ovary tissue of Control rats (A) and rats exposed to CPA (B), CPA + P1 (C), CPA + P2 (D), and CPA+ P3 (E). Image (A) indicates normal, undamaged cells (granulosa and theca cells), with clear round-shaped nuclei a with numerous high electron dense lipidic granules representing hormone depositions. Panel (B) represents the ovary exposed to 200 mg/kg BW CPA, showing several round-shaped oocytes with a smaller number of lipidic granules and significant less electron density. Panel (C) reveals a multi-oocyte follicle belonging to CPA + P1 group, having round nuclei and several lipidic granules. Image (D) is representative for CPA + P2 with a remarkable number and higher electron density of the lipidic structures. Image (E), from CPA + P3 female rats, shows a multi-oocyte follicle with clear round nuclei and a great number of granules that are not very intense in terms of electron density. N = nucleus, M = mitochondria, hg = hormonal granules/lipidic granules, c = capillary, Tc = theca cells, Gc = granulosa cells, zp = zona pellucida.
4. Discussion
It has been demonstrated that chemotherapy alone reduced the ovarian reserve in the growing follicles by activating the apoptosis in the granulosa cells [25,26,27,28]. Besides, oxidative and nitrosative stress are known to damage cellular macromolecules during follicle growth leading to follicular atresia and other damages in the intraovarian microenvironment, developing the so called premature ovarian insufficiency [29,30]. Granulosa cells are easily affected by ROS, mostly by H2O2 [27]. Therefore, in this paper we provided new insights regarding the oxidative stress effects after cyclophosphamide treatments and P. sempervirens therapy of preserving the ovarian physiological function.
In accordance with our previous work [15], P. sempervirens hidroalcoholic extract has showed a strong antioxidant effect in vitro against DPPH and ABTS radicals, as well as in the in vitro cell migration assay. In addition, the same extract was investigated in an in vivo turpentine oil-induced inflammation model, where three different dilutions of the P. sempervirens extract have decreased the oxidative and nitrosative stress markers, such as TOS, OSI, MDA, and NO; meanwhile TAR, SH, CAT, and superoxide dismutase enzyme (SOD) were significantly enhanced in a dose-dependent manner [15].
Here we showed that 200 mg/kg CPA in a unique injection disturbed the homeostasis leading to oxidative and nitrosative stress, as seen in Figure 1, in TOS, OSI, and MDA levels. While it is assumed that CPA metabolic byproducts, such as phosphoramide mustard, create such oxidative injuries in the ovarian reserve, antioxidants are more than required to defend cells and allow them to function in that environment. As such, employing P. sempervirens as an adjuvant pretherapy, did manage to counteract the excess of ROS/RNS by decreasing values of TOS, OSI, and MDA to near-normal or even higher. The same was observed for NO and 3NT. The dose-dependent effect was not clearly noticed, but an interesting observation in TOS and OSI levels was that P2 dose induced a greater inhibition as compared with P1 and P3 doses, meaning that P. sempervirens chemical composition has a high concentration of polyphenols, which can easily become pro-oxidants above a certain dose [15]. It is known and well investigated that polyphenols, mostly flavonoids, can manifest a pro-oxidant reactivity under certain conditions, leading to undesirable effects, many of them related to oxidative stress [31]. Other studies have shown that in vivo dietary polyphenols can interact with transition metal ions in the gastrointestinal tract and since these ions are often unabsorbed, pro-oxidant effects could most likely occur, and among these changes, plasma ROS markers are most prominent to reflect them [32].
With respect to antioxidants, these not only serve as ROS or RNS neutralizers, but other biological effects have been demonstrated over the years, including the inhibition of cycloxygenase, lipoxygenase activity, metaloproteases, and also regulating many signaling pathways (MAPK, P13K, NF-kB) [32,33,34,35]. The antioxidant defense markers investigated in our in vivo experimental model (TAC, CAT, GPx and SH), revealed a contradictory situation. As seen in the results section, TAC is not significantly influenced by CPA, meanwhile the P2 and P3 doses of P. sempervirens extract managed to significantly enhance the TAC levels as compared with CPA group.
On the other hand, the endogenous antioxidant enzymes, GPx and CAT, behaved opposite to each other, after the injection with 200 mg/kg BW, namely CAT significantly decreased, while GPx significantly increased. This is an unusual situation, if compared with previous studies, where endogenous antioxidant enzymes are strongly depleted in various in vivo models of oxidative injuries [36,37,38,39,40]. It is intriguing that GPx reacted in such a significant up-rising way, while the literature has confirmed so many times that this enzyme behaves similarly to its counterparts during stress. GPx has a higher degree of affinity for H2O2 and it also reduces lipidperoxides, meaning it is more resistant as compared with CAT. This could be a possible explanation regarding the differences between these two enzymes, when exposed to CPA. It is worth mentioning the fact that the GPx-CAT cooperativity was noticed by other scientists, who discovered that GPx protects CAT from H2O2 toxicity [41]. In order to elucidate the CAT/GPx enigma during CPA exposure, further studies with different concentrations of CPA are required. However, P. sempervirens pretreatment, in particular in CPA + P3 group, significant changes have been registered as compared to CPA group, irrespective to CAT/GPx abnormalities. Regarding SH concentration, the highest dose (P3) of the extract reestablished the equilibria of the endogenous non-enzymatic antioxidant species to greater values than the control group.
The dose-dependent suppression was also observed in the nitric oxide levels, comparable with 3-NT levels, after pretreatment with all three doses of P. sempervirens extract, as seen in the results section. Our observations support the idea that CPA significantly increased RNS species, while P. sempervirens extract turned out to be a great scavenger for RNS, rather than for oxygen-based species. NO and 3-NT are well known as nitrosative stress markers, but these parameters are essential indicators in inflammatory processes, also common during CPA treatments [42,43]. Therefore, we can argue the anti-inflammatory effect of the extract in the present experimental model. This observation aligns with our previous research regarding P. sempervirens anti-inflammatory potential [15].
Cyclophosphamide is the first chemotherapeutic agent that was demonstrated to induce ovarian toxicity in mouse models and nowadays CPA is used as an experimental model for studying the physiology of the ovaries devoid of follicles [44]. 200 mg/kg BW CPA was proved to suppress half of the ovarian follicle pool [9]. Our experiment was performed with a 200 mg/kg CPA for a mild level of toxicity and still preserving the ovarian function. Thus, the E2 concentrations are slightly decreased in CPA group, as seen in Figure 3b, while FSH were similarly lowered, but with a greater significance. In rodents, several studies have shown a reduction in E2 and an increase in FSH levels, during chemotherapy [45,46]. This unusual change must be because we used a single unique dose with a mild amount of CPA that aimed to reduce, not to destroy, the ovarian cycle. Hence, 200 mg of CPA, one single time, may induce negative feedback on the pituitary gland with a consequence of reduction in its serum level. P. sempervirens extract has shown a dose-dependent increase in FSH and E2 levels, near normal control values. In this situation, our hypothesis is that the chemical composition of the extract is rich in flavonoids, which are well-known for their estrogen-like activity, interacting with endogenous estrogen receptors (ERα,β). In addition, our study provides data on folliculogenesis dynamics. From the 15th day of the experiment, when animals received the CPA injection, the oestrus cycles remained in the metoestrus stage, at least until the end of the experiment, which was the 21st day. This observation confirms the fact that folliculogenesis could have been blocked for a few days, with an inhibition of the pituitary-gonadal axis. More studies need to be done for clarifying the exact mechanisms and hormonal changes with a focus on their dynamics in time. P. sempervirens extract managed to place back the oestrus cycles in a normal dynamic, which increases its therapeutic potential and consideration for pharmaceutical formulations in this field.
There is a significant lack of information in the literature concerning the transmission electron microscopy of the ovaries after chemotherapy. The ultra-thin sections presented in our study revealed several lipidic granules that we assumed to be hormonal granules in almost every group, with a lesser intensity in CPA group and with a numerous number and intensity after administration of P. sempervirens. The grey (less dark) aspect of the lipid droplets is thought to be directly linked to lipolysis and increased ROS accumulation in oocytes [47]. An aspect that may be preserved to all ovarian cells. It is noteworthy that the CPA + P2 group had the most electron-dense granules, but the CPA + P3 group had an impressive number of those structures. In the control group, there is a group of theca cells, nicely layered, as compared to the granulosa cells, which are concentrically disposed. A single unique dose of cyclophosphamide (CPA group) induced several morphologic changes as seen in the chromatin aggregated pattern in the perinuclear area and the swelling of several mitochondria, events which mark the beginning of the apoptosis process. Similar changes were suggested by other scientists [48] and related to most chemotherapy treatment effects on the ovaries [49,50]. Moreover, the structure of the granulosa cells appeared to be very well preserved after P. sempervirens treatment.
Similar results were obtained in vitro with doxorubicine and Cimicifuga racemosa (L.) nutt extract, where the chemotherapy induced apoptosis in follicular and stromal cells, depleted the lipid droplets, but the ethanolic extract of C. racemosa has reduced those effects [51]. The common phenolic compound of C. racemosa and P. sempervirens is ferulic acid, a compound that was previously shown to help oocyte development in vitro, reduce ovarian oxidative stress [52], and protect against irradiation-induced apoptosis [53,54].
5. Conclusions
In summary, cyclophosphamide generates oxidative and nitrosative stress, reduced levels of endogenous antioxidants with imbalances in GPx/CAT systems, low levels of sex hormones, a temporarily suppression of follicular-pituitary dynamics, and ultrastructural changes in granulosa cells with clear indicators of apoptotic events. However, a xerophyte such as, P. sempervirens, administered for 21 days to rats could significantly reduce the deleterious effects of CPA by up-regulating the antioxidant status (SH, CAT, TAC) and down-regulating the nitro-oxidative stress markers (TOS, OSI, NO, 3-NT). Therefore, our main results suggest that P. sempervirens has significant protective effects against nitrosative stress, along with folliculogenesis regulation (oestrus cycle keeper).
Further studies should be employed for investigating the cooperativity of GPx-CAT during CPA exposure, as well as FSH dynamics in different moments of the experiment. The main limitation of our study is the reduced number of animals; therefore future experiments with several more groups, and different doses of cyclophosphamide, could reveal important data regarding its aggressiveness and also potential mechanisms of the extract protection.
Our findings could represent an important base for future investigations in this field that might lead to developing drugs or food supplements with an ovarian protective role useful for many patients that are facing ovarian cancer.
Author Contributions
Conceptualization, A.D.S. and A.E.P.; methodology, A.D.S., B.S. and M.S.; investigation, A.D.S., B.S., I.R. and V.A.T.; resources, M.P.; data curation, C.D.; writing—original draft preparation, A.D.S.; writing—review and editing, B.S., C.D., A.E.P., M.S., M.P., I.R. and V.A.T.; supervision, M.P.; project administration, A.D.S.; funding acquisition, A.D.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a grant from the Ministry of Research, Innovation and Digitization, CNCS-UEFISCDI, project number PN-III-P1-1.1-PD-2021-0286, within PNCDI III.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee of Iuliu Hațieganu University of Medicine and Pharmacy (22/13.12.2016).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chi, Y.-N.; Yang, J.-M.; Liu, N.; Cui, Y.-H.; Ma, L.; Lan, X.-B.; Ma, W.-Q.; Liu, Y.-J.; Yu, J.-Q.; Du, J. Development of Protective Agents against Ovarian Injury Caused by Chemotherapeutic Drugs. Biomed. Pharmacother. 2022, 155, 113731. [Google Scholar] [CrossRef]
- Behranvand, N.; Nasri, F.; Zolfaghari Emameh, R.; Khani, P.; Hosseini, A.; Garssen, J.; Falak, R. Chemotherapy: A Double-Edged Sword in Cancer Treatment. Cancer Immunol. Immunother. 2022, 71, 507–526. [Google Scholar] [CrossRef]
- Li, G.; Zhou, J.; Sun, M.; Cen, J.; Xu, J. Role of Luteolin Extracted from Clerodendrum Cyrtophyllum Turcz Leaves in Protecting HepG2 Cells from TBHP-Induced Oxidative Stress and Its Cytotoxicity, Genotoxicity. J. Funct. Foods 2020, 74, 104196. [Google Scholar] [CrossRef]
- Chen, Q.; Xu, Z.; Li, X.; Du, D.; Wu, T.; Zhou, S.; Yan, W.; Wu, M.; Jin, Y.; Zhang, J.; et al. Epigallocatechin Gallate and Theaflavins Independently Alleviate Cyclophosphamide-Induced Ovarian Damage by Inhibiting the Overactivation of Primordial Follicles and Follicular Atresia. Phytomedicine 2021, 92, 153752. [Google Scholar] [CrossRef]
- Lins, T.L.B.G.; Gouveia, B.B.; Barberino, R.S.; Silva, R.L.S.; Monte, A.P.O.; Pinto, J.G.C.; Campinho, D.S.P.; Palheta, R.C., Jr.; Matos, M.H.T. Rutin Prevents Cisplatin-Induced Ovarian Damage via Antioxidant Activity and Regulation of PTEN and FOXO3a Phosphorylation in Mouse Model. Reprod. Toxicol. 2020, 98, 209–217. [Google Scholar] [CrossRef]
- Liu, X.; Song, Y.; Zhou, F.; Zhang, C.; Li, F.; Hu, R.; Ma, W.; Song, K.; Tang, Z.; Zhang, M. Network and Experimental Pharmacology on Mechanism of Si-Wu-Tang Improving Ovarian Function in a Mouse Model of Premature Ovarian Failure Induced by Cyclophosphamide. J. Ethnopharmacol. 2023, 301, 115842. [Google Scholar] [CrossRef]
- Barberino, R.S.; Lins, T.L.B.G.; Monte, A.P.O.; Silva, R.L.S.; Andrade, K.O.; Campinho, D.S.P.; Palheta Junior, R.C.; Smitz, J.E.J.; Matos, M.H.T. Epigallocatechin-3-Gallate Attenuates Cyclophosphamide-Induced Damage in Mouse Ovarian Tissue via Suppressing Inflammation, Apoptosis, and Expression of Phosphorylated Akt, FOXO3a and RpS6. Reprod. Toxicol. 2022, 113, 42–51. [Google Scholar] [CrossRef]
- Masciangelo, R.; Hossay, C.; Chiti, M.C.; Manavella, D.D.; Amorim, C.A.; Donnez, J.; Dolmans, M.-M. Role of the PI3K and Hippo Pathways in Follicle Activation after Grafting of Human Ovarian Tissue. J. Assist. Reprod. Genet. 2020, 37, 101–108. [Google Scholar] [CrossRef]
- Horicks, F.; Van Den Steen, G.; Houben, S.; Englert, Y.; Demeestere, I. Folliculogenesis Is Not Fully Inhibited during GnRH Analogues Treatment in Mice Challenging Their Efficiency to Preserve the Ovarian Reserve during Chemotherapy in This Model. PLoS ONE 2015, 10, e0137164. [Google Scholar] [CrossRef]
- Rampogu, S.; Balasubramaniyam, T.; Lee, J.-H. Phytotherapeutic Applications of Alkaloids in Treating Breast Cancer. Biomed. Pharmacother. 2022, 155, 113760. [Google Scholar] [CrossRef]
- Liu, Y.-Q.; Wang, X.-L.; He, D.-H.; Cheng, Y.-X. Protection against Chemotherapy- and Radiotherapy-Induced Side Effects: A Review Based on the Mechanisms and Therapeutic Opportunities of Phytochemicals. Phytomedicine 2021, 80, 153402. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M.; Nybom, H.; Lindholm, C.; Rumpunen, K. Major Polyphenols in Aerial Organs of Greater Plantain (Plantago major L.), and Effects of Drying Temperature on Polyphenol Contents in the Leaves. Sci. Hortic. 2011, 128, 523–529. [Google Scholar] [CrossRef]
- Beara, I.N.; Lesjak, M.M.; Orčić, D.Z.; Simin, N.Đ.; Četojević-Simin, D.D.; Božin, B.N.; Mimica-Dukić, N.M. Comparative Analysis of Phenolic Profile, Antioxidant, Anti-Inflammatory and Cytotoxic Activity of Two Closely-Related Plantain Species: Plantago altissima L. and Plantago lanceolata L. LWT—Food Sci. Technol. 2012, 47, 64–70. [Google Scholar] [CrossRef]
- Gonçalves, S.; Romano, A. The Medicinal Potential of Plants from the Genus Plantago (Plantaginaceae). Ind. Crops Prod. 2016, 83, 213–226. [Google Scholar] [CrossRef]
- Farcaș, A.D.; Moț, A.C.; Pârvu, A.E.; Toma, V.A.; Popa, M.A.; Mihai, M.C.; Sevastre, B.; Roman, I.; Vlase, L.; Pârvu, M. In Vivo Pharmacological and Anti-Inflammatory Evaluation of Xerophyte Plantago sempervirens Crantz. Oxid. Med. Cell Longev. 2019, 2019, 5049643. [Google Scholar] [CrossRef]
- Farcaș, A.D.; Zăgrean-Tuza, C.; Vlase, L.; Gheldiu, A.-M.; Pârvu, M.; Moț, A.C. EPR Fingerprinting and Antioxidant Response of Four Selected Plantago Species. Stud. Univ. Babeș-Bolyai Chem. 2020, 65, 209–220. [Google Scholar] [CrossRef]
- Harma, M.; Harma, M.; Erel, O. Increased Oxidative Stress in Patients with Hydatidiform Mole. Swiss Med. Wkly. 2003, 133, 563–566. [Google Scholar] [CrossRef]
- Erel, O. A Novel Automated Method to Measure Total Antioxidant Response against Potent Free Radical Reactions. Clin. Biochem. 2004, 37, 112–119. [Google Scholar] [CrossRef]
- Miranda, K.M.; Espey, M.G.; Wink, D.A. A Rapid, Simple Spectrophotometric Method for Simultaneous Detection of Nitrate and Nitrite. Nitric Oxide 2001, 5, 62–71. [Google Scholar] [CrossRef]
- Mitev, D.; Gradeva, H.; Stoyanova, Z.; Petrova, N.; Karova, N.; Dimov, D.; Iliev, V.; Koychev, A.; Prakova, G.; Vlaykova, T. Evaluation of Thiol Compounds and Lipid Peroxidative Products in Plasma of Patients With COPD. Trakia J. Sci. 2010, 8, 306–314. [Google Scholar]
- Li, Y.; Schellhorn, H.E. Rapid Kinetic Microassay for Catalase Activity. J. Biomol. Tech. JBT 2007, 18, 185. [Google Scholar] [PubMed]
- Byers, S.L.; Wiles, M.V.; Dunn, S.L.; Taft, R.A. Mouse Estrous Cycle Identification Tool and Images. PLoS ONE 2012, 7, e35538. [Google Scholar] [CrossRef] [PubMed]
- Suciu, M.; Mirescu, C.; Crăciunescu, I.; Macavei, S.G.; Leoștean, C.; Ştefan, R.; Olar, L.E.; Tripon, S.-C.; Ciorîță, A.; Barbu-Tudoran, L. In Vivo Distribution of Poly(Ethylene Glycol) Functionalized Iron Oxide Nanoclusters: An Ultrastructural Study. Nanomaterials 2021, 11, 2184. [Google Scholar] [CrossRef] [PubMed]
- Toiu, A.; Mocan, A.; Vlase, L.; Pârvu, A.E.; Vodnar, D.C.; Gheldiu, A.-M.; Moldovan, C.; Oniga, I. Comparative Phytochemical Profile, Antioxidant, Antimicrobial and In Vivo Anti-Inflammatory Activity of Different Extracts of Traditionally Used Romanian Ajuga genevensis L. and A. Reptans L. (Lamiaceae). Molecules 2019, 24, 1597. [Google Scholar] [CrossRef]
- Roness, H.; Kashi, O.; Meirow, D. Prevention of Chemotherapy-Induced Ovarian Damage. Fertil. Steril. 2016, 105, 20–29. [Google Scholar] [CrossRef]
- Melekoglu, R.; Ciftci, O.; Eraslan, S.; Cetin, A.; Basak, N. Beneficial Effects of Curcumin and Capsaicin on Cyclophosphamide-Induced Premature Ovarian Failure in a Rat Model. J. Ovarian Res. 2018, 11, 33. [Google Scholar] [CrossRef]
- Yang, H.; Xie, Y.; Yang, D.; Ren, D. Oxidative Stress-Induced Apoptosis in Granulosa Cells Involves JNK, P53 and Puma. Oncotarget 2017, 8, 25310–25322. [Google Scholar] [CrossRef]
- Shen, M.; Lin, F.; Zhang, J.; Tang, Y.; Chen, W.-K.; Liu, H. Involvement of the Up-Regulated FoxO1 Expression in Follicular Granulosa Cell Apoptosis Induced by Oxidative Stress. J. Biol. Chem. 2012, 287, 25727–25740. [Google Scholar] [CrossRef]
- Yang, L.; Chen, Y.; Liu, Y.; Xing, Y.; Miao, C.; Zhao, Y.; Chang, X.; Zhang, Q. The Role of Oxidative Stress and Natural Antioxidants in Ovarian Aging. Front. Pharmacol. 2021, 11, 617843. [Google Scholar] [CrossRef]
- Dumesic, D.A.; Meldrum, D.R.; Katz-Jaffe, M.G.; Krisher, R.L.; Schoolcraft, W.B. Oocyte Environment: Follicular Fluid and Cumulus Cells Are Critical for Oocyte Health. Fertil. Steril. 2015, 103, 303–316. [Google Scholar] [CrossRef]
- Moţ, A.C.; Coman, C.; Miron, C.; Damian, G.; Sarbu, C.; Silaghi-Dumitrescu, R. An Assay for Pro-Oxidant Reactivity Based on Phenoxyl Radicals Generated by Laccase. Food Chem. 2014, 143, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Are Polyphenols Antioxidants or Pro-Oxidants? What Do We Learn from Cell Culture and in Vivo Studies? Arch. Biochem. Biophys. 2008, 476, 107–112. [Google Scholar] [CrossRef] [PubMed]
- De Torre, M.P.; Cavero, R.Y.; Calvo, M.I.; Vizmanos, J.L. A Simple and a Reliable Method to Quantify Antioxidant Activity In Vivo. Antioxidants 2019, 8, 142. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, P.; Shikha, D.; Thakur, M.; Aneja, A. Functionality of Apigenin as a Potent Antioxidant with Emphasis on Bioavailability, Metabolism, Action Mechanism and in Vitro and in Vivo Studies: A Review. J. Food Biochem. 2022, 46, e13950. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.; Almeida, A.; López-Yerena, A.; Pinto, S.; Sarmento, B.; Lamuela-Raventós, R.; Vallverdú-Queralt, A.; Delerue-Matos, C.; Rodrigues, F. Appraisal of a New Potential Antioxidants-Rich Nutraceutical Ingredient from Chestnut Shells through in-Vivo Assays—A Targeted Metabolomic Approach in Phenolic Compounds. Food Chem. 2023, 404, 134546. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Pan, J.-H.; Huang, X.-F.; Liao, Y.-Q.; Ling, Y.-J.; Luo, J.-Y. Effect of Melatonin on Oxidative Stress Indicators in Animal Models of Fibrosis: A Systematic Review and Meta-Analysis. Free Radic. Biol. Med. 2023, 195, 158–177. [Google Scholar] [CrossRef] [PubMed]
- Issac, P.K.; Guru, A.; Velayutham, M.; Pachaiappan, R.; Arasu, M.V.; Al-Dhabi, N.A.; Choi, K.C.; Harikrishnan, R.; Arockiaraj, J. Oxidative Stress Induced Antioxidant and Neurotoxicity Demonstrated in Vivo Zebrafish Embryo or Larval Model and Their Normalization Due to Morin Showing Therapeutic Implications. Life Sci. 2021, 283, 119864. [Google Scholar] [CrossRef]
- Beken, A.T.; Saka, Ş.; Aydın, İ.; Fırat, K.; Suzer, C.; Benzer, F.; Erişir, M.; Özden, O.; Hekimoğlu, M.A.; Engin, S.; et al. In Vivo and in Vitro Evolution of the Effects of Cypermethrin on Turbot (Scophthalmus maximus, Linnaeus, 1758) Spermatozoa. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2022, 256, 109298. [Google Scholar] [CrossRef]
- Rashid, S.; Sameti, M.; Alqarni, M.H.; Abdel Bar, F.M. In Vivo Investigation of the Inhibitory Effect of Peganum Harmala L. and Its Major Alkaloids on Ethylene Glycol-Induced Urolithiasis in Rats. J. Ethnopharmacol. 2023, 300, 115752. [Google Scholar] [CrossRef]
- Grover, P.; Khanna, K.; Bhatnagar, A.; Purkayastha, J. In Vivo-Wound Healing Studies of Sodium Thiosulfate Gel in Rats. Biomed. Pharmacother. 2021, 140, 111797. [Google Scholar] [CrossRef]
- Baud, O. Glutathione Peroxidase-Catalase Cooperativity Is Required for Resistance to Hydrogen Peroxide by Mature Rat Oligodendrocytes. J. Neurosci. 2004, 24, 1531–1540. [Google Scholar] [CrossRef] [PubMed]
- Erdal, N.; Gürgül, S.; Tamer, L.; Ayaz, L. Effects of Long-Term Exposure of Extremely Low Frequency Magnetic Field on Oxidative/Nitrosative Stress in Rat Liver. J. Radiat. Res. 2008, 49, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Ricciardolo, F.L.M.; Sorbello, V.; Benedetto, S.; Paleari, D. Effect of Ambroxol and Beclomethasone on Lipopolysaccharide-Induced Nitrosative Stress in Bronchial Epithelial Cells. Respiration 2015, 89, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Spears, N.; Lopes, F.; Stefansdottir, A.; Rossi, V.; De Felici, M.; Anderson, R.A.; Klinger, F.G. Ovarian Damage from Chemotherapy and Current Approaches to Its Protection. Hum. Reprod. Update 2019, 25, 673–693. [Google Scholar] [CrossRef]
- Yuksel, A.; Bildik, G.; Senbabaoglu, F.; Akin, N.; Arvas, M.; Unal, F.; Kilic, Y.; Karanfil, I.; Eryılmaz, B.; Yilmaz, P.; et al. The Magnitude of Gonadotoxicity of Chemotherapy Drugs on Ovarian Follicles and Granulosa Cells Varies Depending upon the Category of the Drugs and the Type of Granulosa Cells. Hum. Reprod. 2015, 30, 2926–2935. [Google Scholar] [CrossRef]
- Fu, X.; He, Y.; Xie, C.; Liu, W. Bone Marrow Mesenchymal Stem Cell Transplantation Improves Ovarian Function and Structure in Rats with Chemotherapy-Induced Ovarian Damage. Cytotherapy 2008, 10, 353–363. [Google Scholar] [CrossRef]
- Isachenko, V.; Isachenko, E.; Michelmann, H.W.; Alabart, J.L.; Vazquez, I.; Bezugly, N.; Nawroth, F. Lipolysis and Ultrastructural Changes of Intracellular Lipid Vesicles after Cooling of Bovine and Porcine GV-Oocytes. Anat. Histol. Embryol. J. Vet. Med. Ser. C 2001, 30, 333–338. [Google Scholar] [CrossRef]
- Zhao, X.; Huang, Y.; Yu, Y.; Xin, X. GnRH Antagonist Cetrorelix Inhibits Mitochondria-Dependent Apoptosis Triggered by Chemotherapy in Granulosa Cells of Rats. Gynecol. Oncol. 2010, 118, 69–75. [Google Scholar] [CrossRef]
- Sonigo, C.; Beau, I.; Binart, N.; Grynberg, M. The Impact of Chemotherapy on the Ovaries: Molecular Aspects and the Prevention of Ovarian Damage. Int. J. Mol. Sci. 2019, 20, 5342. [Google Scholar] [CrossRef]
- Xiong, J.; Xue, L.; Li, Y.; Tang, W.; Chen, D.; Zhang, J.; Dai, J.; Zhou, S.; Lu, Z.; Wu, M.; et al. Therapy of Endocrine Disease: Novel Protection and Treatment Strategies for Chemotherapy-Associated Ovarian Damage. Eur. J. Endocrinol. 2021, 184, R177–R192. [Google Scholar] [CrossRef]
- De Assis, E.I.T.; Azevedo, V.A.N.; De Lima Neto, M.F.; Costa, F.C.; Paulino, L.R.F.M.; Barroso, P.A.A.; Donato, M.A.M.; Peixoto, C.A.; Do Monte, A.P.O.; Matos, M.H.T.; et al. Protective Effect of Cimicifuga racemosa (L.) Nutt Extract on Oocyte and Follicle Toxicity Induced by Doxorubicin during In Vitro Culture of Mice Ovaries. Animals 2022, 13, 18. [Google Scholar] [CrossRef] [PubMed]
- Tanihara, F.; Hirata, M.; Nhien, N.T.; Hirano, T.; Kunihara, T.; Otoi, T. Effect of Ferulic Acid Supplementation on the Developmental Competence of Porcine Embryos during in Vitro Maturation. J. Vet. Med. Sci. 2018, 80, 1007–1011. [Google Scholar] [CrossRef] [PubMed]
- Abdel Fattah, S.M.; Mohmed, H.K.; Mohamed, M.A.E.H. The Potential Protective Effect of Ferulic Acid against Gamma Irradiation Induced Ovarian Failure in Rats. Egypt. J. Radiat. Sci. Appl. 2019, 32, 1–12. [Google Scholar] [CrossRef]
- Mun, G.-I.; Kim, S.; Choi, E.; Kim, C.S.; Lee, Y.-S. Pharmacology of Natural Radioprotectors. Arch. Pharm. Res. 2018, 41, 1033–1050. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).