
Changes in Oil Quality and Peroxidase Activity during Germination of Rape Seeds and Mustard Seeds


 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Materials
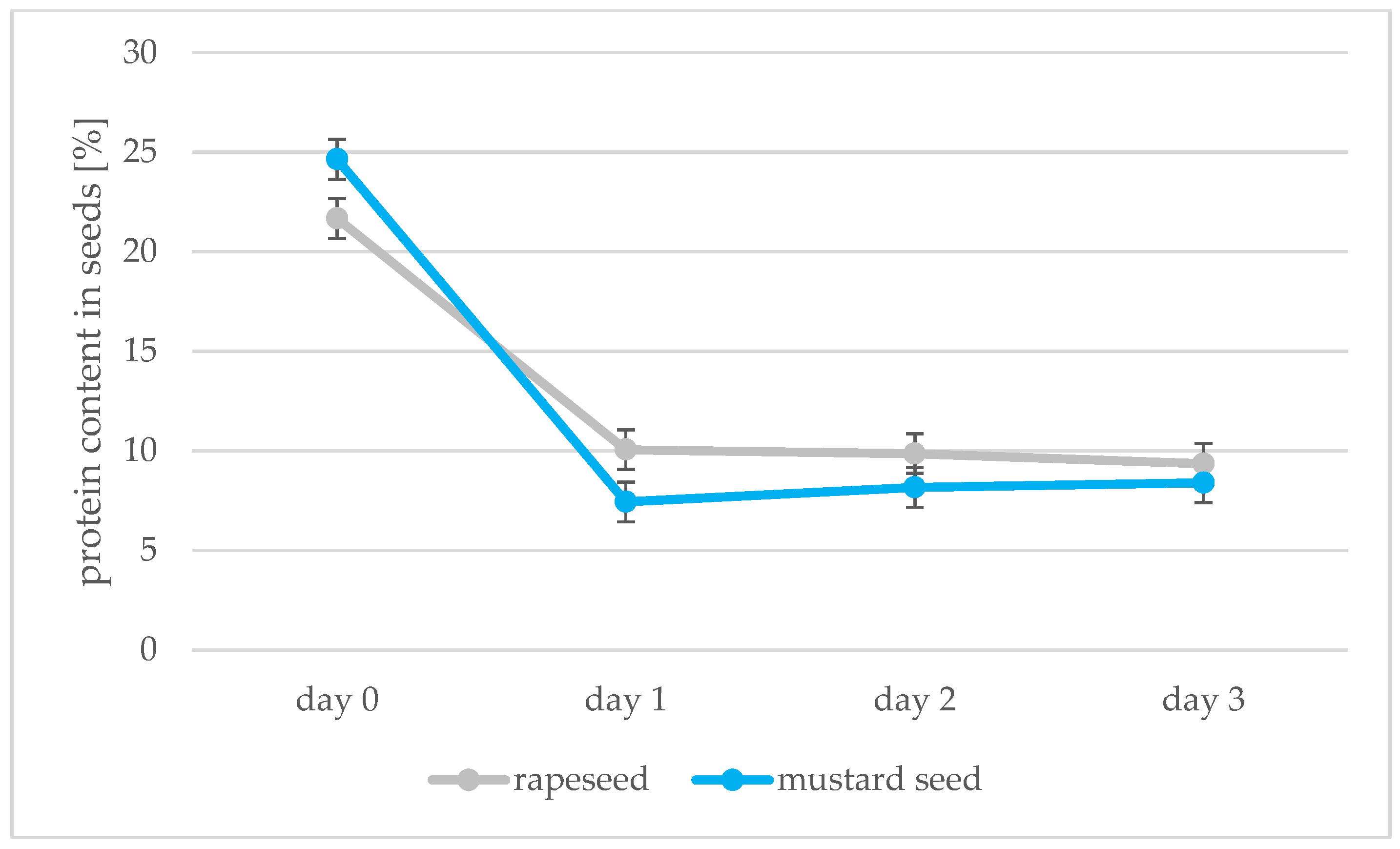
2.3. Determination of Protein Content
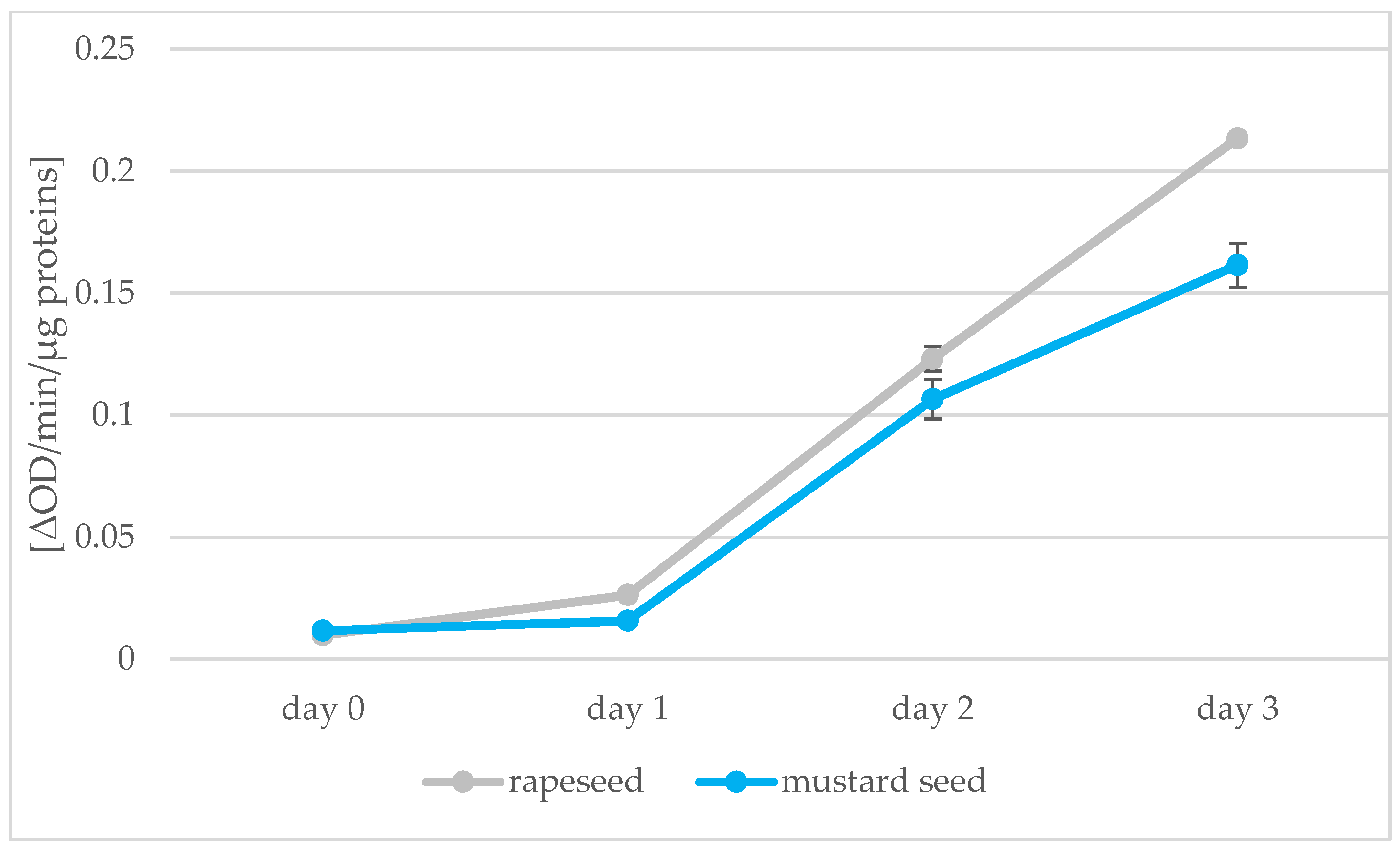
2.4. Peroxidase Enzyme Activity
2.5. Extraction of Oil from Seeds
2.6. Acid Value and Peroxide Value
2.7. Fatty acid Analysis
2.8. Sterol Analysis
2.9. Tocopherol Analysis
2.10. Statistical Analysis
3. Results and Discussion
3.1. Changes in Fat Content during Seed Germination
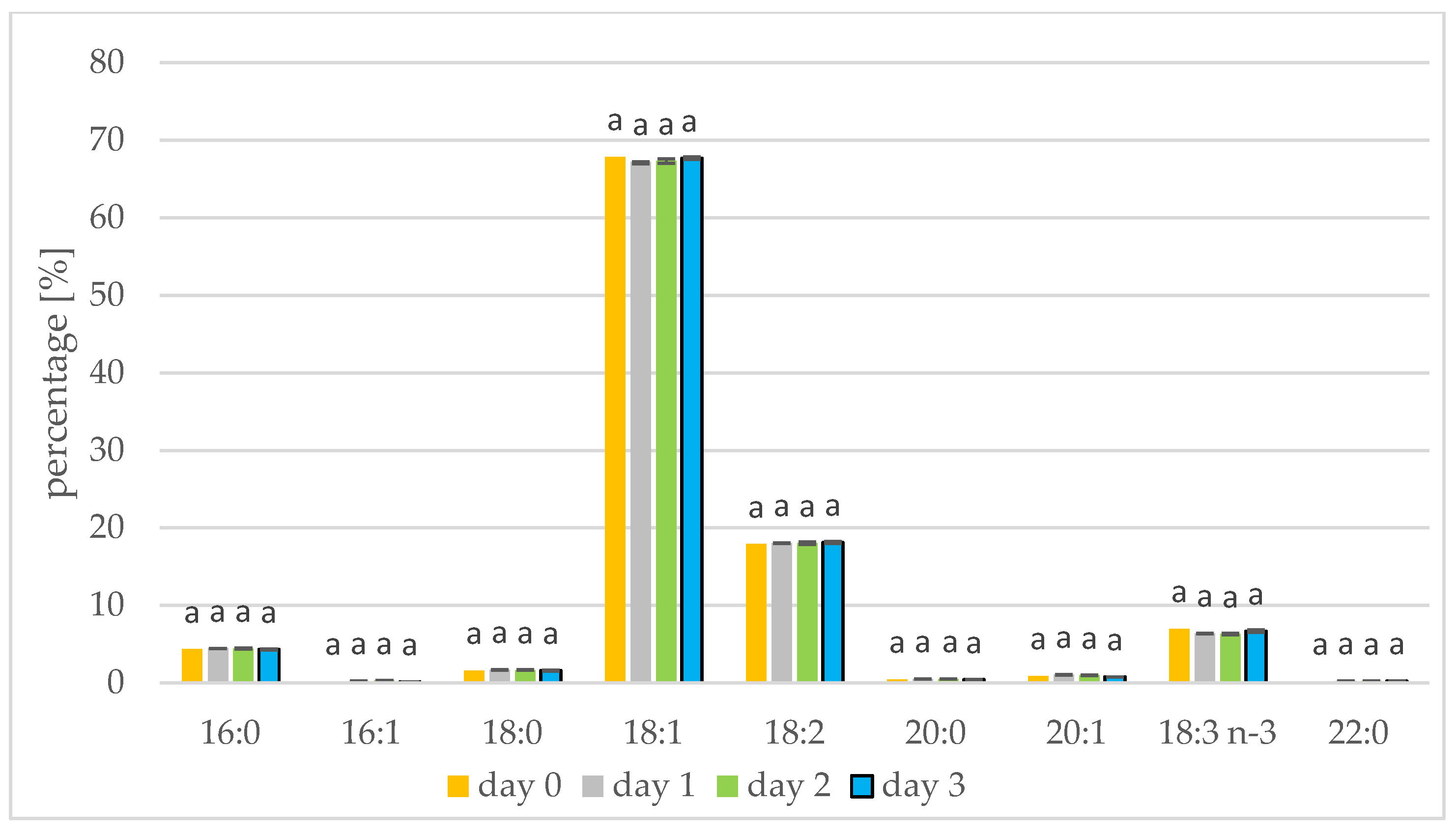
3.2. Changes in Fatty Acids during Seed Germination
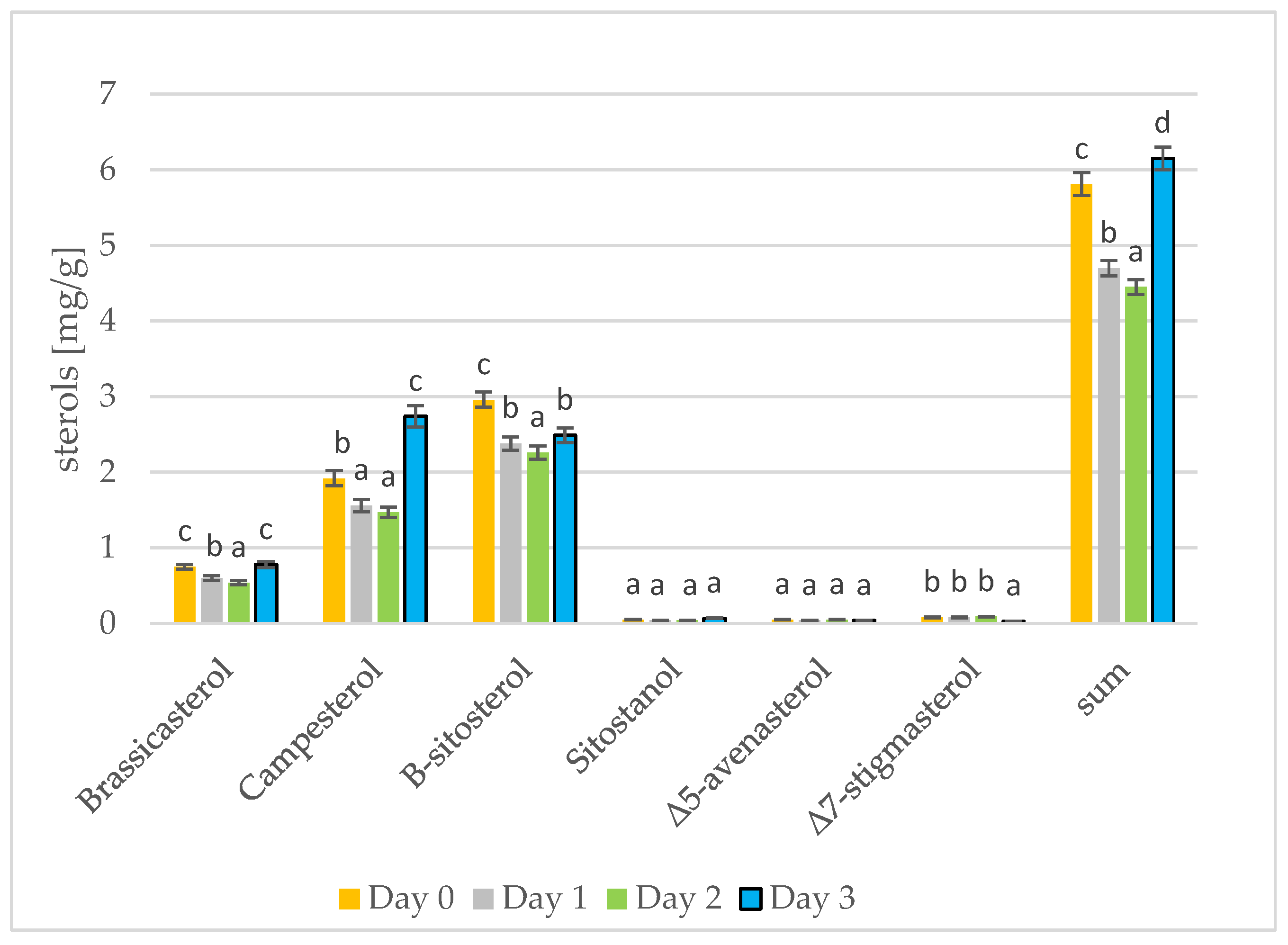
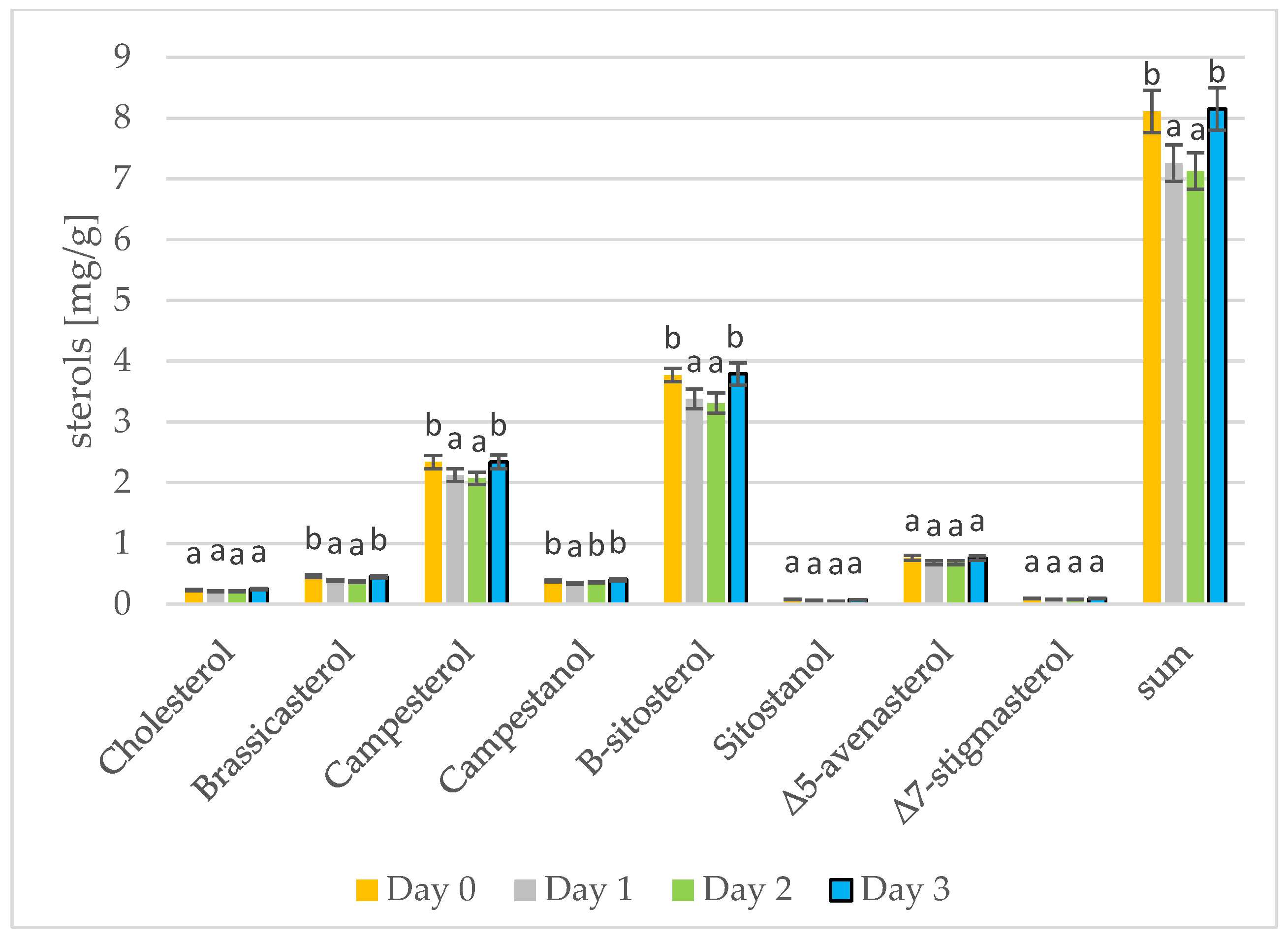
3.3. Changes in Sterols during Germination
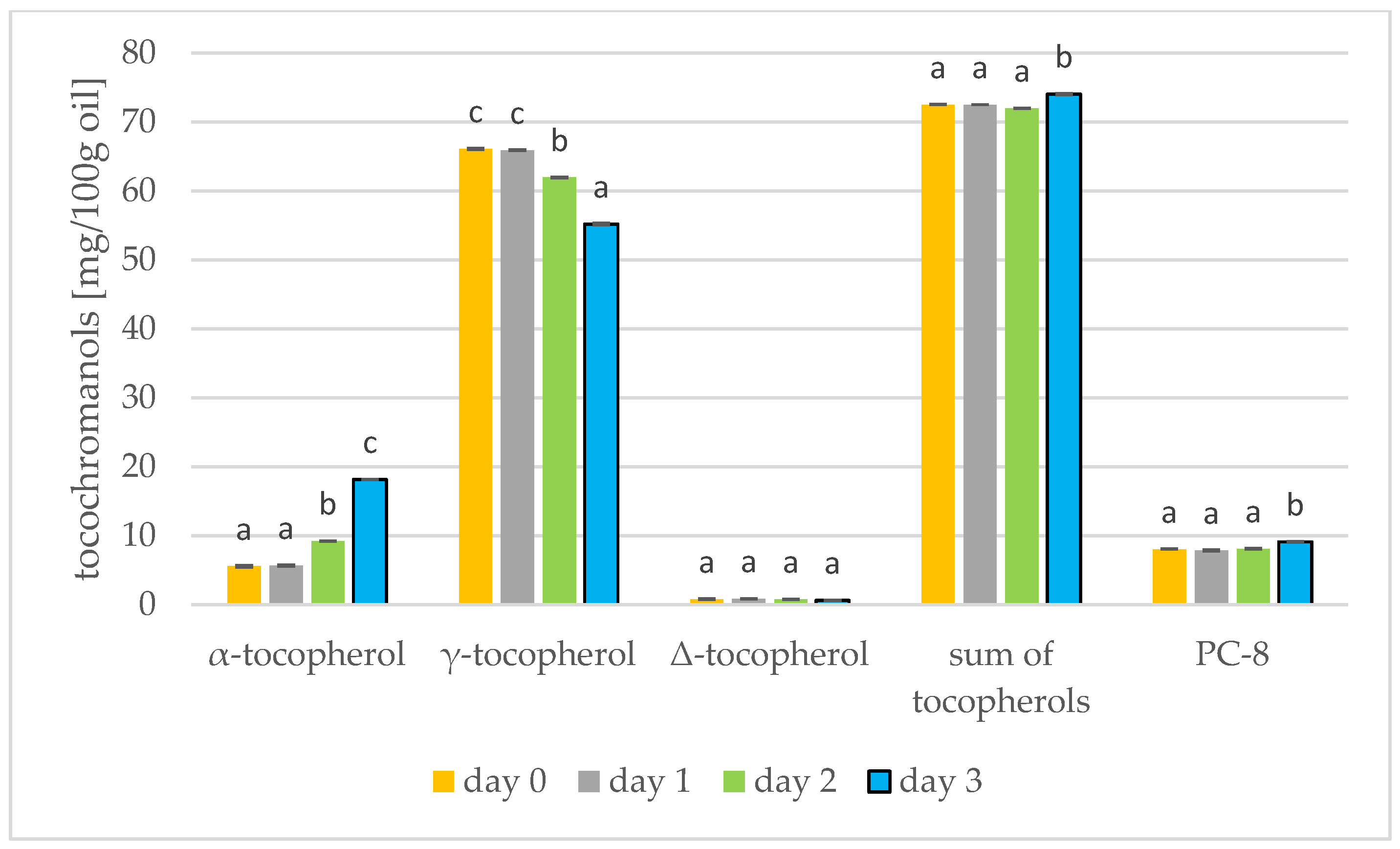
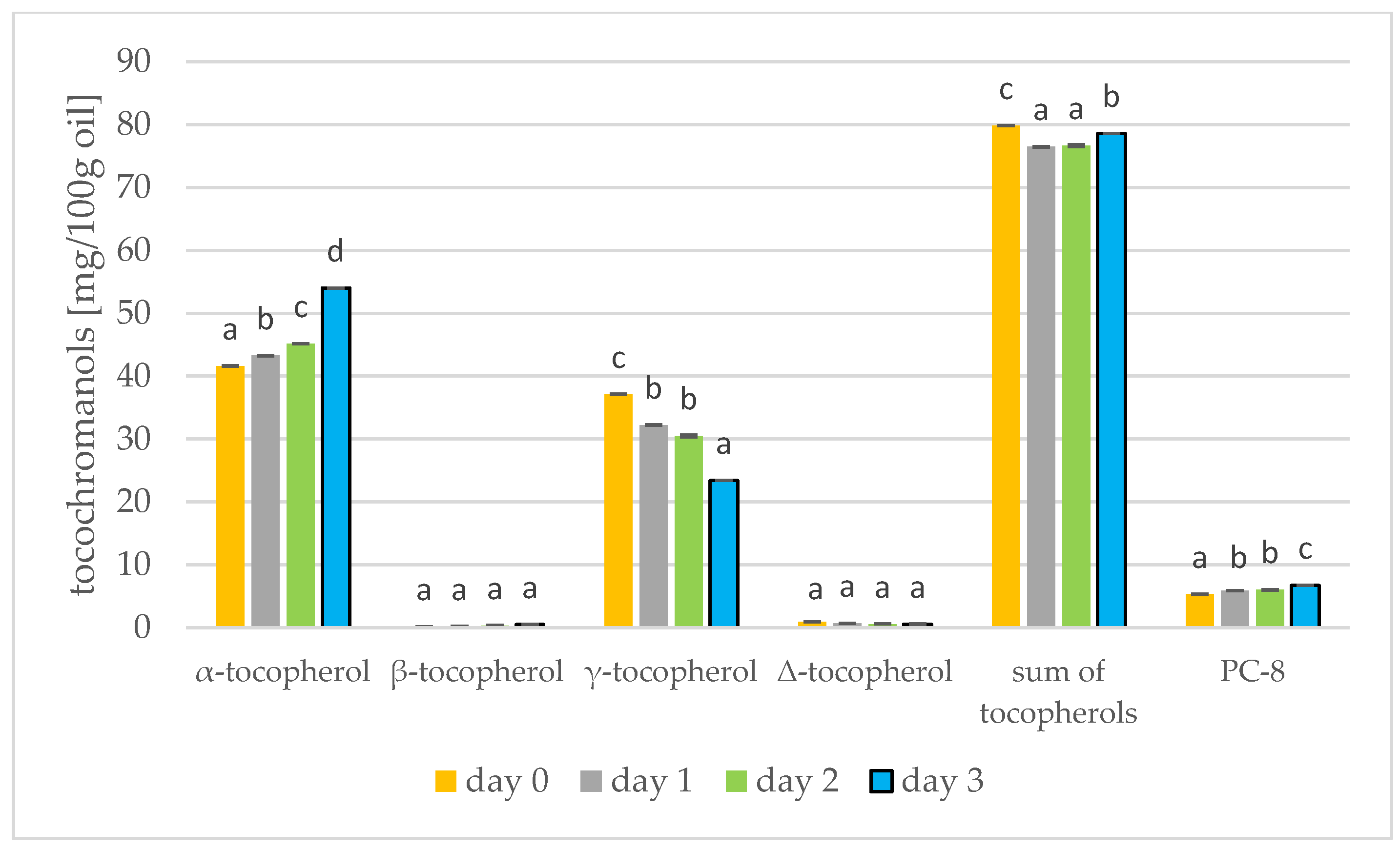
3.4. Changes in Tocochromanols during Germination
3.5. Oil Quality
3.6. Comparison of Enzymatic Activity in Seeds and Sprouts of Rape and Mustard
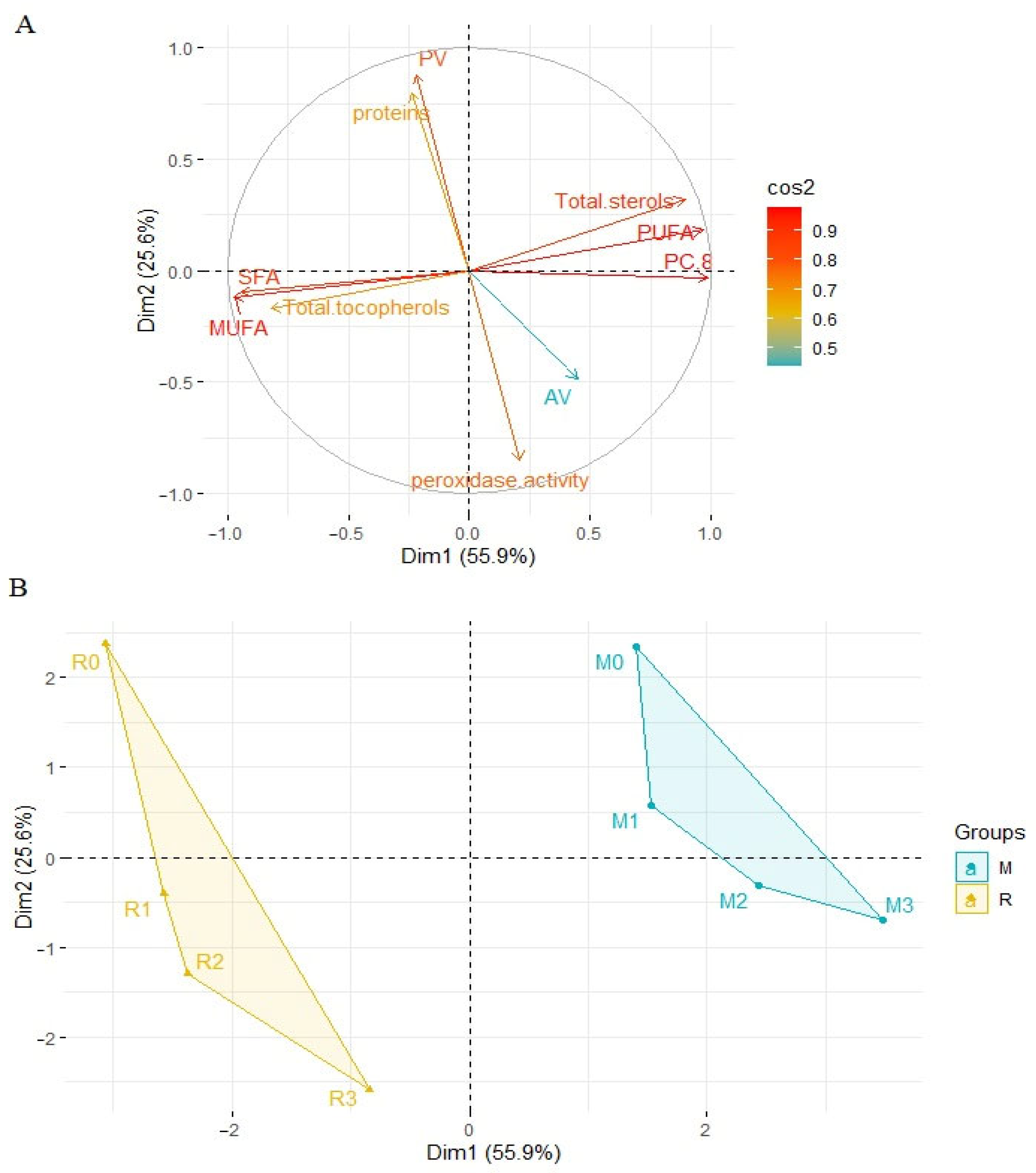
3.7. Principal Component Analysis (PCA)
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wroniak, M.; Chlebowska-Śmigiel, A. Impact of purity of rapeseed and oil purification method on selected properties of cold-pressed oils. Zywnosc Nauka Technol. Jakosc 2013, 4, 133–149. [Google Scholar] [CrossRef]
- Zhang, N.; Li, Y.; Wen, S.; Sun, Y.; Chen, J.; Gao, Y.; Sagymbek, A.; Yu, X. Analytical methods for determining the peroxide value of edible oils: A mini-review. Food Chem. 2021, 358, 129834. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Huyan, Z.; Geng, Q.; Yu, X. Rapid Determination of Acid Value of Edible Oils via FTIR Spectroscopy Using Infrared Quartz Cuvette. J. Oleo Sci. 2019, 68, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of The United Nations. The Codex Alimentarius Fats, Oils and Related Products; World Health Organization: Rome, Italy, 2001. [Google Scholar]
- Czwartkowski, K.; Wierzbic, A.; Golimowski, W. Quality, key production factors, and consumption volume of niche edible oils marketed in the European Union. Sustainability 2022, 14, 1846. [Google Scholar] [CrossRef]
- Rękas, A.; Wroniak, M.; Siger, A.; Ścibisz, I. Chemical composition and resistance to oxidation of high-oleic rapeseed oil pressed from microwave pre-treated intact and de-hulled seeds. Grasas Aceites 2017, 68, e225. [Google Scholar] [CrossRef]
- Corrales, C.V.; Achir, N.; Forestier, N.; Lebrun, M.; Maraval, I.; Dornier, M.; Perez, A.M.; Vaillant, F.; Fliedel, G. Innovative process combining roasting and tempering to mechanically dehull jicaro seeds (Crescentia alata K.H.B). J. Food Eng. 2017, 212, 283–290. [Google Scholar] [CrossRef]
- Nederal, S.; Skevin, D.; Kraljić, K.; Obranović, M.; Papesa, S.; Bataljaku, A. Chemical Composition and Oxidative Stability of Roasted and Cold Pressed Pumpkin Seed Oils. J. Am. Oil Chem. Soc. 2012, 89, 1763–1770. [Google Scholar] [CrossRef]
- Luo, F.; Fei, X. Maillard reaction derived from oil-tea camellia seed through roasting. J. Sci. Food Agric. 2019, 99, 5000–5007. [Google Scholar] [CrossRef]
- Vo, B.V.; Siddik, M.A.B.; Fotedar, R.; Chaklader, M.R.; Foysal, M.J.; Pham, H.D. Digestibility and water quality investigations on the processed peanut (Arachis hypogaea) meal fed barramundi (Lates calcarifer) at various inclusion levels. Aquac. Rep. 2020, 18, 100474. [Google Scholar] [CrossRef]
- Cui, J.; Lamade, E.; Tcherkez, G. Seed Germination in Oil Palm (Elaeis guineensis Jacq.): A Review of Metabolic Pathways and Control Mechanisms. Int. J. Mol. Sci. 2020, 21, 4227. [Google Scholar] [CrossRef]
- Gu, J.; Hou, D.; Li, Y.; Chao, H.; Zhang, K.; Wang, H.; Xiang, J.; Raboanatahiry, N.; Wang, B.; Li, M. Integration of proteomic and genomic approaches to dissect seed germination vigor in Brassica napus seeds differing in oil content. BMC Plant Biol. 2019, 19, 21. [Google Scholar] [CrossRef]
- Pachari Vera, E.; Alca, J.J.; Rondón Saravia, G.; Callejas Campioni, N.; Jachmanián Alpuy, I. Comparison of the lipid profile and tocopherol content of four Peruvian quinoa (Chenopodium quinoa Willd.) cultivars (‘Amarilla de Maranganí’, ‘Blanca de Juli’, INIA 415 ‘Roja Pasankalla’, INIA 420 ‘Negra Collana’) during germination. J. Cereal Sci. 2019, 88, 132–137. [Google Scholar] [CrossRef]
- Chauhan, A.; Kumari, N.; Saxena, D.C.; Singh, S. Effect of germination on fatty acid profile, amino acid profile and minerals of amaranth (Amaranthus spp.) grain. J. Food Meas. Charact. 2022, 16, 1777–1786. [Google Scholar] [CrossRef]
- Chandrasekaran, U.; Liu, A. Stage-specific metabolization of triacylglycerols during seed germination of Sacha Inchi (Plukenetia volubilis L.). J. Sci. Food Agric. 2015, 95, 1764–1766. [Google Scholar] [CrossRef]
- Guo, S.; Klinkesorn, U.; Lorjaroenphon, Y.; Ge, Y.; Na Jom, K. Effects of germinating temperature and time on metabolite profiles of sunflower (Helianthus annuus L.) seed. Food Sci. Nutr. 2021, 9, 2810–2822. [Google Scholar] [CrossRef]
- Shi, H.; Nam, P.K.; Ma, Y. Comprehensive Profiling of Isoflavones, Phytosterols, Tocopherols, Minerals, Crude Protein, Lipid, and Sugar during Soybean (Glycine max) Germination. J. Agric. Food Chem. 2010, 58, 4970–4976. [Google Scholar] [CrossRef]
- Herchi, W.; Bahashwan, S.; Sebei, K.; Saleh, H.B.; Kallel, H.; Boukhchina, S. Effects of germination on chemical composition and antioxidant activity of flaxseed (Linum usitatissimum L.) oil. Grasas Aceites 2015, 66, 1–8. [Google Scholar] [CrossRef]
- Kohli, S.K.; Khanna, K.; Bhardwaj, R.; Abd_Allah, E.F.; Ahmad, P.; Corpas, F.J. Assessment of Subcellular ROS and NO Metabolism in Higher Plants: Multifunctional Signaling Molecules. Antioxidants 2019, 8, 641. [Google Scholar] [CrossRef]
- Mansoor, S.; Wani, O.A.; Lone, J.K.; Manhas, S.; Kour, N.; Alam, P.; Ahmad, A.; Ahmad, P. Reactive Oxygen Species in Plants: From Source to Sink. Antioxidants 2022, 11, 225. [Google Scholar] [CrossRef]
- Singh, K.L.; Chaudhuri, A.; Kar, R.K. Role of peroxidase activity and Ca2+ in axis growth during seed germination. Planta 2015, 242, 997–1007. [Google Scholar] [CrossRef]
- Ahmad, P.; Tripathi, D.K.; Deshmukh, R.; Singh, V.P.; Corpas, F.J. Revisiting the role of ROS and RNS in plants under changing environment. Environ. Exp. Bot. 2019, 161, 1–3. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Yamamoto, K.; Tawaratsumida, T.; Yuasa, T.; Iwaya-Inoue, M. Hydrogen peroxide scavenging regulates germination ability during wheat (Triticum aestivum L.) seed maturation. Plant Signal. Behav. 2008, 3, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Consumption of Vegetable Oils Worldwide from 2013/14 to 2021/2022, by Oil Type. Available online: https://www.statista.com/statistics/263937/vegetable-oils-global-consumption/ (accessed on 17 January 2023).
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Guha, P.; Mandal, A.K. Deteriorative changes in enzyme activity of non-invigorated and invigorated soybean seeds (Glycine max [l.] merrill, cv. soyamax). Legum. Res. 2019, 42, 633–639. [Google Scholar] [CrossRef]
- Górnaś, P.; Siger, A.; Rudzińska, M.; Grygier, A.; Marszałkiewicz, S.; Ying, Q.; Sobieszczańska, N.; Segliņa, D. Impact of the Extraction Technique and Genotype on the Oil Yield and Composition of Lipophilic Compounds in the Oil Recovered from Japanese Quince (Chaenomeles japonica) Seeds. Eur. J. Lipid Sci Technol. 2019, 121, 1–10. [Google Scholar] [CrossRef]
- ISO 660:2010; Animal and Vegetable Fats and Oils—Determination of Acid Value and Acidity. Polish Committee of Standardization International: Warsaw, Poland, 2010. Available online: https://www.iso.org/standard/75594.html (accessed on 17 January 2023).
- ISO 3960:2017; Animal and Vegetable Fats and Oils—Determination of Peroxide Value—Iodometric (Visual) Endpoint Determination. Polish Committee for Standardization: Warsaw, Poland, 2017. Available online: https://www.iso.org/standard/71268.html (accessed on 17 January 2023).
- Raczyk, M.; Polanowska, K.; Kruszewski, B.; Grygier, A.; Michałowska, D. Effect of spirulina (Arthrospira platensis) supplementation on physical and chemical properties of semolina (Triticum durum) Based Fresh Pasta. Molecules 2022, 27, 355. [Google Scholar] [CrossRef]
- Sahu, P.K.; Chakradhari, S.; Sipeniece, E.; Mišina, I.; Qian, Y.; Grygier, A.; Rudzińska, M.; Patel, K.S.; Górnaś, P. Fatty Acids, Tocopherols, Tocotrienols, Phytosterols, Carotenoids, and Squalene in Seed Oils of Hyptis suaveolens, Leonotis nepetifolia, and Ocimum sanctum. Eur. J. Lipid Sci. Technol. 2020, 122, 1–5. [Google Scholar] [CrossRef]
- Xu, M.; Jin, Z.; Simsek, S.; Hall, C.; Rao, J.; Chen, B. Effect of germination on the chemical composition, thermal, pasting, and moisture sorption properties of flours from chickpea, lentil, and yellow pea. Food Chem. 2019, 295, 579–587. [Google Scholar] [CrossRef]
- Borek, S.; Kubala, S.; Kubala, S.; Ratajczak, L. Comparative study of storage compound breakdown in germinating seeds of three lupine species. Acta Physiol. Plant 2011, 33, 1953–1968. [Google Scholar] [CrossRef]
- Sakiroh, S.; Taryono, T.; Purwanti, S. Dynamics of storage materials in cotyledon during cocoa seed germination. Ilmu Pertan. (Agric. Sci.) 2018, 3, 12–20. [Google Scholar] [CrossRef]
- Hahm, T.; Park, S.; Lo, M.Y. Effects of germination on chemical composition and functional properties of sesame (Sesamum indicum L.) seeds. Bioresour. Technol. 2009, 100, 1643–1647. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Marangoni, F.; Poli, A. Phytosterols and cardiovascular health. Pharmacol. Res. 2010, 61, 193–199. [Google Scholar] [CrossRef]
- Bush, P.B.; Grunwald, C. Sterol Changes during Germination of Nicotiana tabacum Seeds. Plant Physiol. 1972, 50, 69–72. [Google Scholar] [CrossRef][Green Version]
- Rękas, A.; Siger, A.; Wroniak, M.; Ścibisz, I.; Derewiaka, D.; Anders, A. Influence of de-hulled rapeseed roasting on the physicochemical composition and oxidative state of oil. Grasas Aceites 2017, 68, e176. [Google Scholar] [CrossRef]
- Piironen, V.; Lindsay, D.; Miettinen, T.; Toivo, J.; Lampi, A.M. Plant sterols: Biosynthesis, biological function and their importance to human nutrition. J. Sci. Food Agric. 2020, 80, 939–966. [Google Scholar] [CrossRef]
- Valitova, J.N.; Sulkarnayeva, A.G.; Minibayeva, F.V. Plant Sterols: Diversity, Biosynthesis, and Physiological Functions. Biochem. Biokhimiia 2016, 81, 819–834. [Google Scholar] [CrossRef]
- Shu, X.L.; Frank, T.; Shu, Q.Y.; Engel, K.H. Metabolite profiling of germinating rice seeds. J. Agric. Food Chem. 2008, 56, 11612–11620. [Google Scholar] [CrossRef]
- García-Navarro, E.; Pérez-Vich, B.; Velasco, L. Changes in plastochromanol-8 and tocopherols during germination in Ethiopian mustard lines with contrasting tocopherol levels. Seed Sci. Res. 2014, 24, 101–112. [Google Scholar] [CrossRef]
- Zhang, H.; Vasanthan, T.; Wettasinghe, M. Enrichment of tocopherols and phytosterols in canola oil during seed germination. J. Agric. Food Chem. 2007, 55, 355–359. [Google Scholar] [CrossRef]
- Azzi, A. Many tocopherols, one vitamin E. Mol. Aspects Med. 2018, 61, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, H.A.; Ahmad, E.E.M.; Mariod, A.A.; Matthäus, B.; Salaheldeen, M. Effect of pretreatment on the proximate composition, physicochemical characteristics and stability of Moringa peregrina oil. Grasas Aceites 2017, 68, e227. [Google Scholar] [CrossRef]
- Lewicki, P.P. Kiełki nasion jako źródło cennych składników odżywczych. Zywn. Nauka Technol. Jakosc 2010, 6, 18–33. [Google Scholar]
- Wang, K.H.; Lai, Y.H.; Chang, J.C.; Ko, T.F.; Shyu, S.L.; Chiou, R.Y. Germination of Peanut Kernels to Enhance Resveratrol Biosynthesis and Prepare Sprouts as a Functional Vegetable. Agric. Food Chem. 2005, 53, 242–246. [Google Scholar] [CrossRef]
- Scilabba, A.; Bellani, L.M.; Dell’Aquila, A. Effects of ageing on peroxidase activity and localization in radish (Raphanus sativus L.) seeds. Eur. J. Histochem. 2002, 46, 351–358. [Google Scholar] [CrossRef]
- Palmiano, E.P.; Juliano, B.O. Changes in the activity of some hydrolases, peroxidase, and catalase in the rice seed during germination. Plant Physiol. 1973, 52, 274–277. [Google Scholar] [CrossRef]
- Omidiji, O.; Okpuzor, J.; Otubu, O. Peroxidase activity of germinating Sorghum bicolor grains: Effect of some cations. J. Sci. Food Agric. 2002, 82, 1881–1885. [Google Scholar] [CrossRef]
- Ramaiah, P.K.; Durzan, D.J.; Mia, A.J. Amino acids, soluble proteins, and isoenzyme patterns of peroxidase during the germination of jack pine. Can. J. Plant Sci. 1971, 49, 2151–2161. [Google Scholar] [CrossRef]










{kind=link}
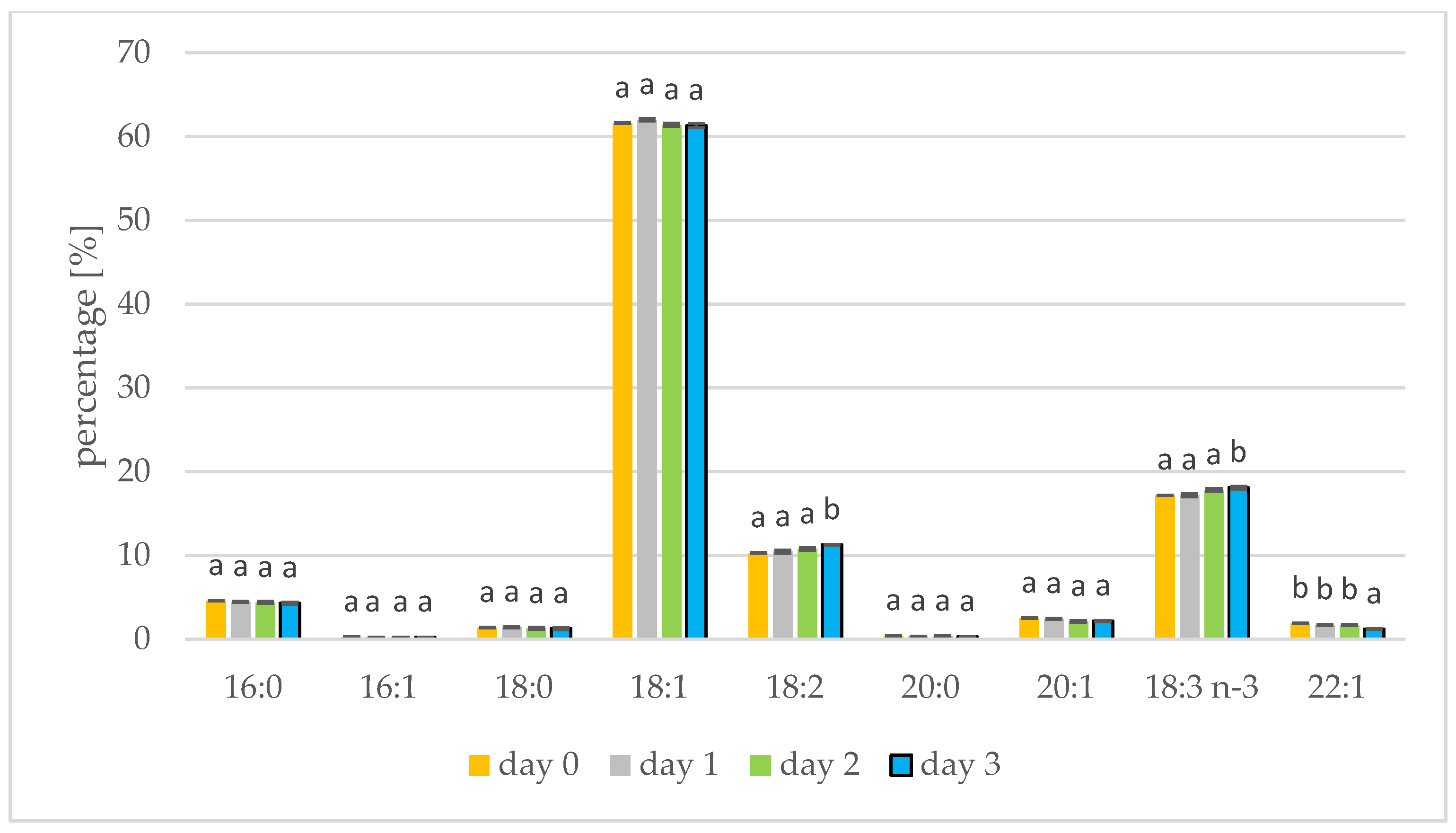{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Content [%] | |
|---|---|
| R0 | 41.5 d ± 0.2 |
| R1 | 32.0 c ± 0.2 |
| R2 | 20.7 a ± 0.1 |
| R3 | 22.1 b ± 0.2 |
| M0 | 26.0 a ± 0.2 |
| M1 | 27.1 b ± 0.2 |
| M2 | 27.1 b ± 0.1 |
| M3 | 27.0 b ± 0.0 |
| Peroxide Value [meq O2/kg] | Acid Value [mg KOH/g] | |||
|---|---|---|---|---|
| Rapeseed Oil | Mustard Oil | Rapeseed Oil | Mustard Oil | |
| Day 0 | 11.53 | 7.47 | 0.67 | 5.61 |
| Day 1 | 1.56 | 4.54 | 0.99 | 3.80 |
| Day 2 | 1.58 | 2.18 | 2.24 | 4.13 |
| Day 3 | 1.58 | 2.14 | 10.31 | 5.61 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belt, D.; Grygier, A.; Siger, A.; Kmiecik, D.; Spasibionek, S.; Rudzińska, M. Changes in Oil Quality and Peroxidase Activity during Germination of Rape Seeds and Mustard Seeds. Appl. Sci. 2023, 13, 2196. https://doi.org/10.3390/app13042196
Belt D, Grygier A, Siger A, Kmiecik D, Spasibionek S, Rudzińska M. Changes in Oil Quality and Peroxidase Activity during Germination of Rape Seeds and Mustard Seeds. Applied Sciences. 2023; 13(4):2196. https://doi.org/10.3390/app13042196
Chicago/Turabian StyleBelt, Dorota, Anna Grygier, Aleksander Siger, Dominik Kmiecik, Stanisław Spasibionek, and Magdalena Rudzińska. 2023. "Changes in Oil Quality and Peroxidase Activity during Germination of Rape Seeds and Mustard Seeds" Applied Sciences 13, no. 4: 2196. https://doi.org/10.3390/app13042196
APA StyleBelt, D., Grygier, A., Siger, A., Kmiecik, D., Spasibionek, S., & Rudzińska, M. (2023). Changes in Oil Quality and Peroxidase Activity during Germination of Rape Seeds and Mustard Seeds. Applied Sciences, 13(4), 2196. https://doi.org/10.3390/app13042196