
Virulence-Linked Mutations in Rubredoxin Reductase and Glutaredoxin: Impact on Antibiotic Susceptibility and Phage Therapy in Pseudomonas aeruginosa
 ,
,  , ,
, ,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Strains and Maintenance
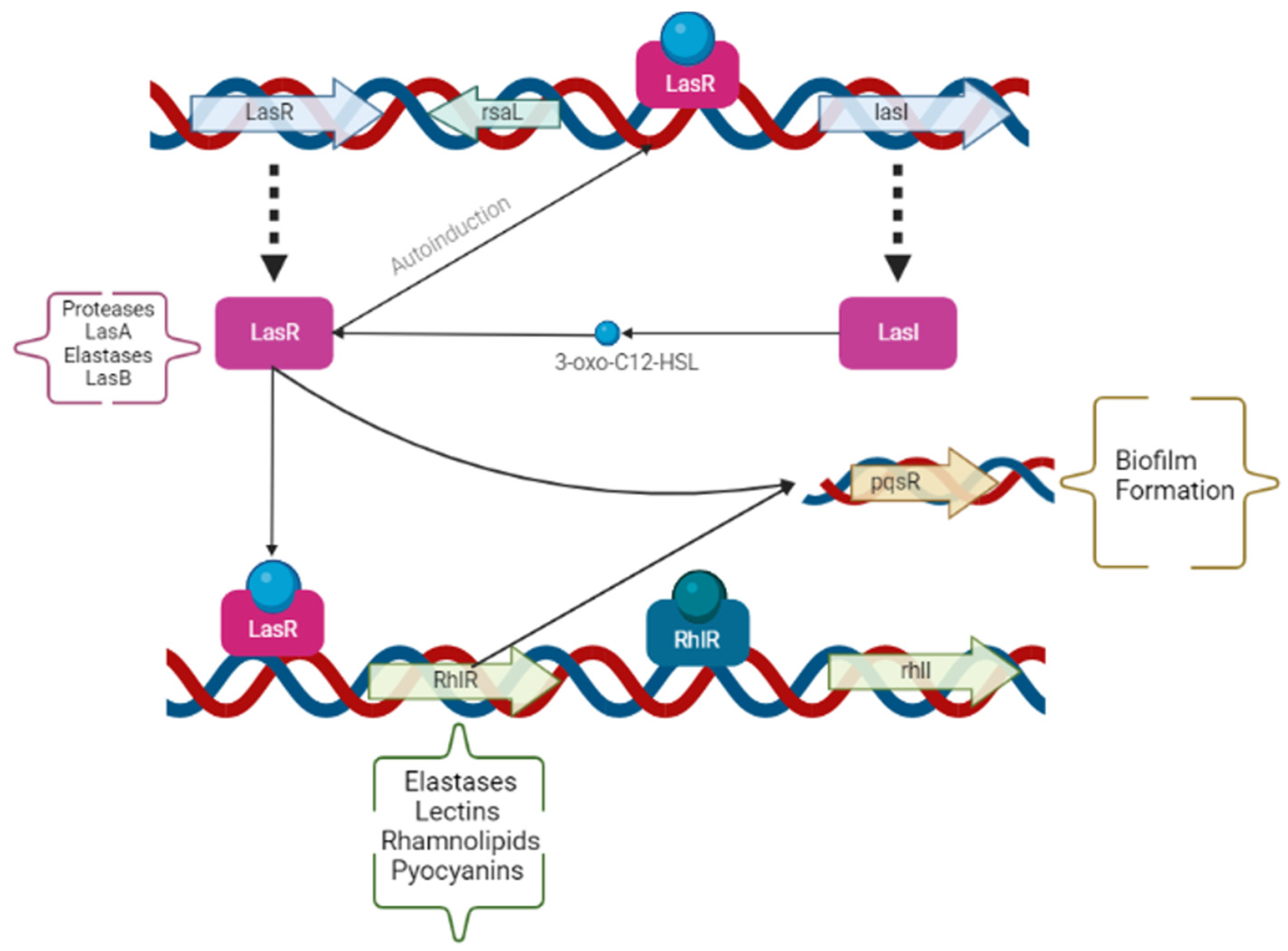
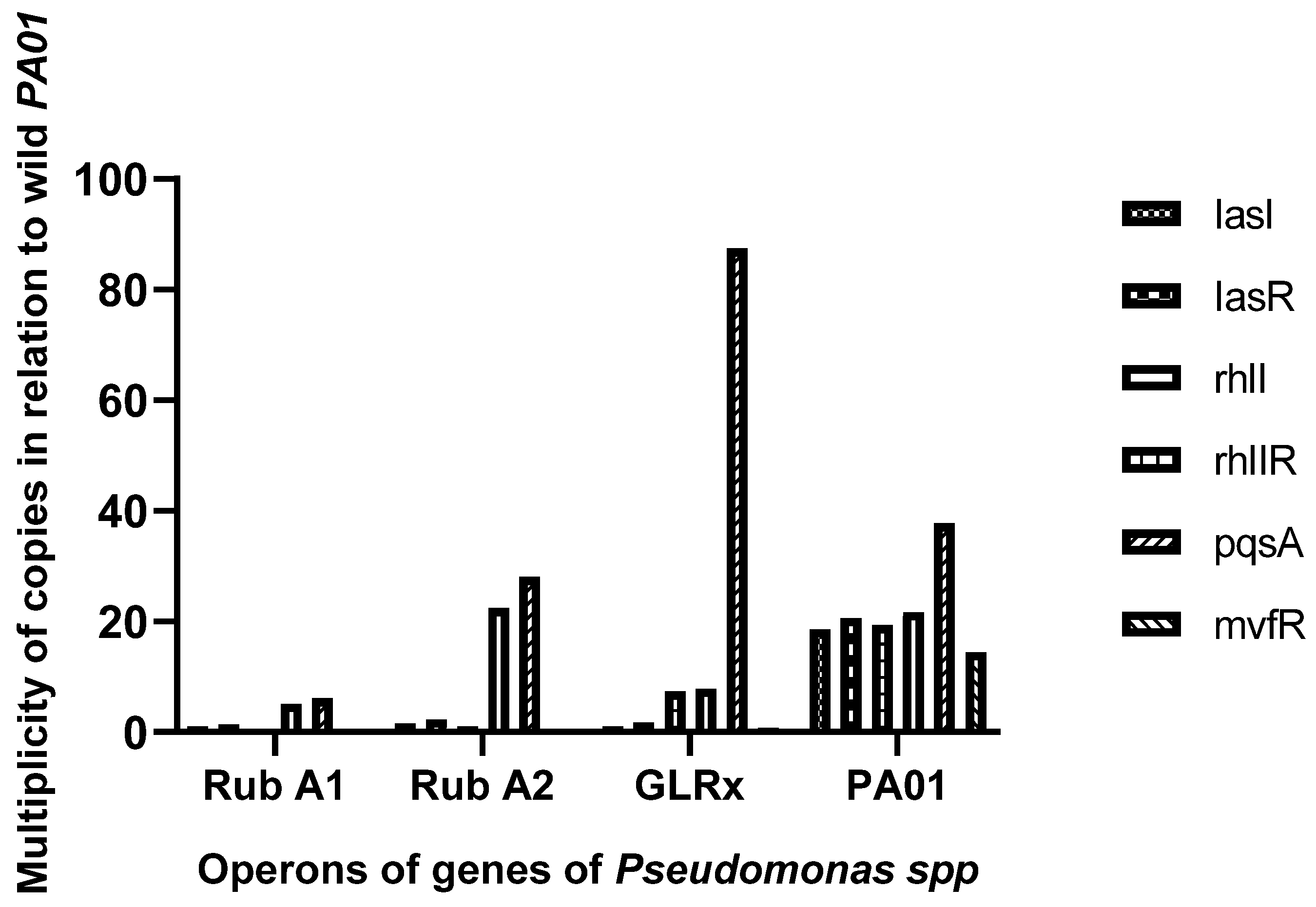
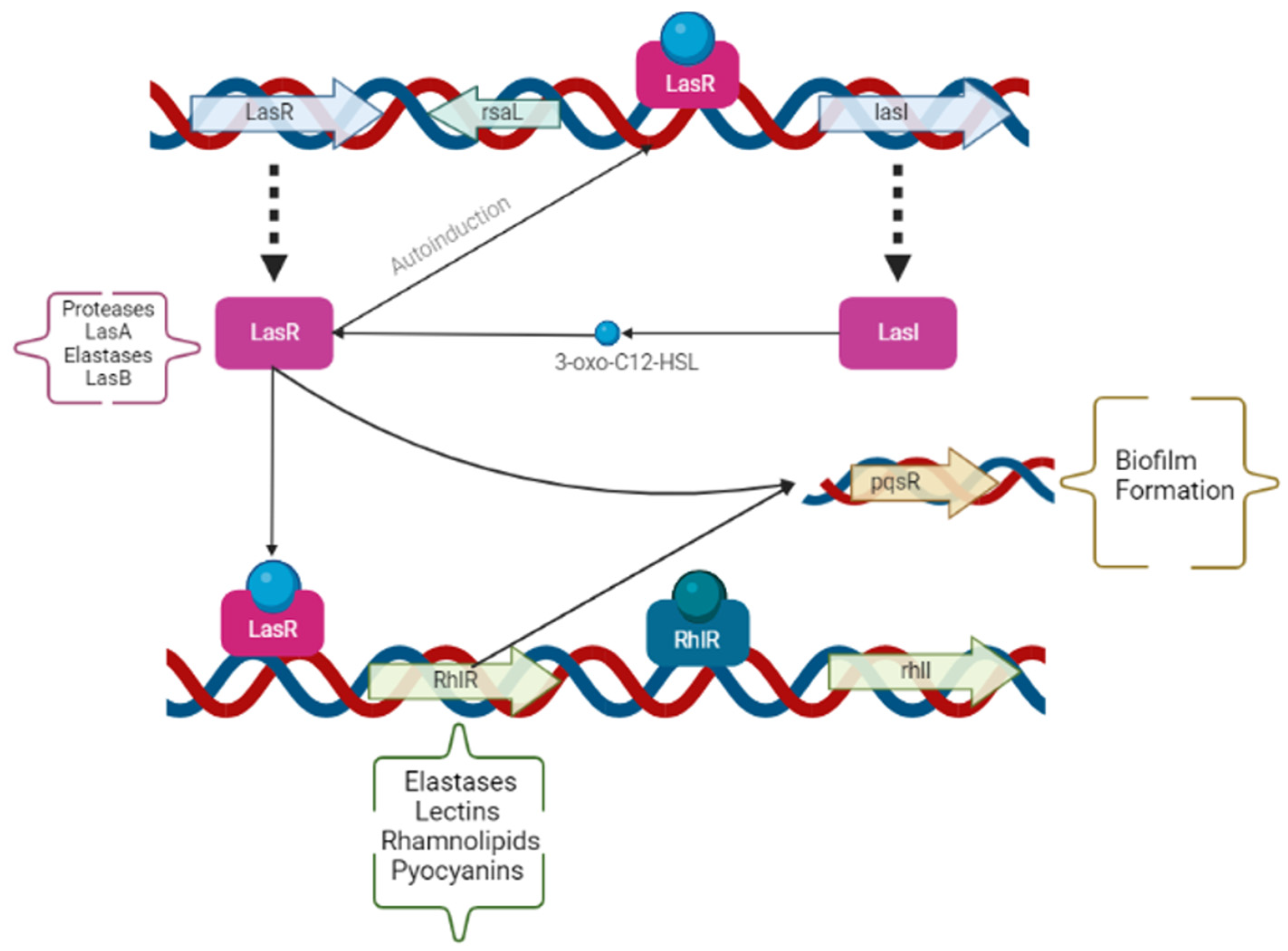
2.3. Quorum Sensing Operons and Gene Expression
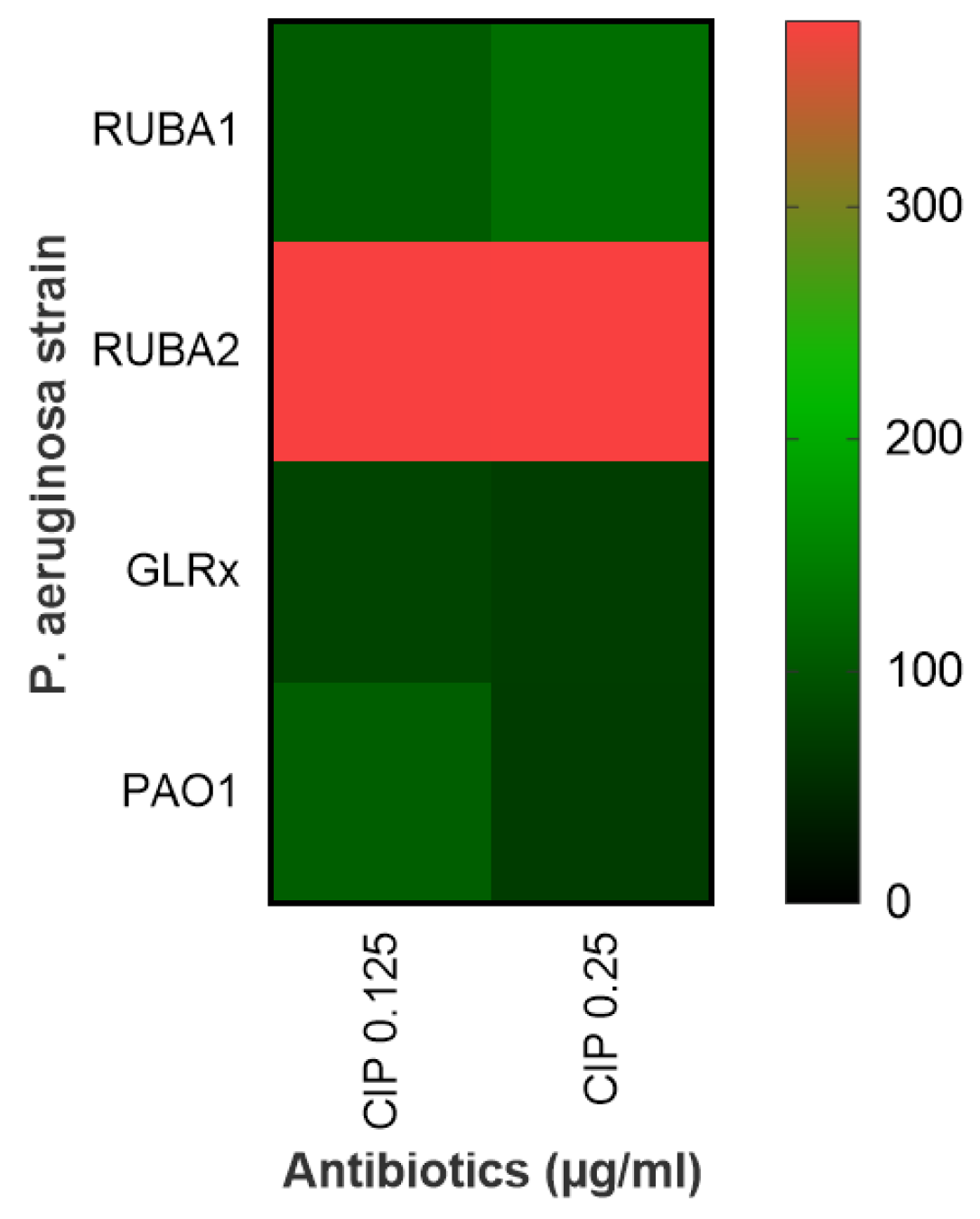
2.4. Viability Assay of Rub A1, Rub A2, GLRX and PAO1 under CIP Concentrations
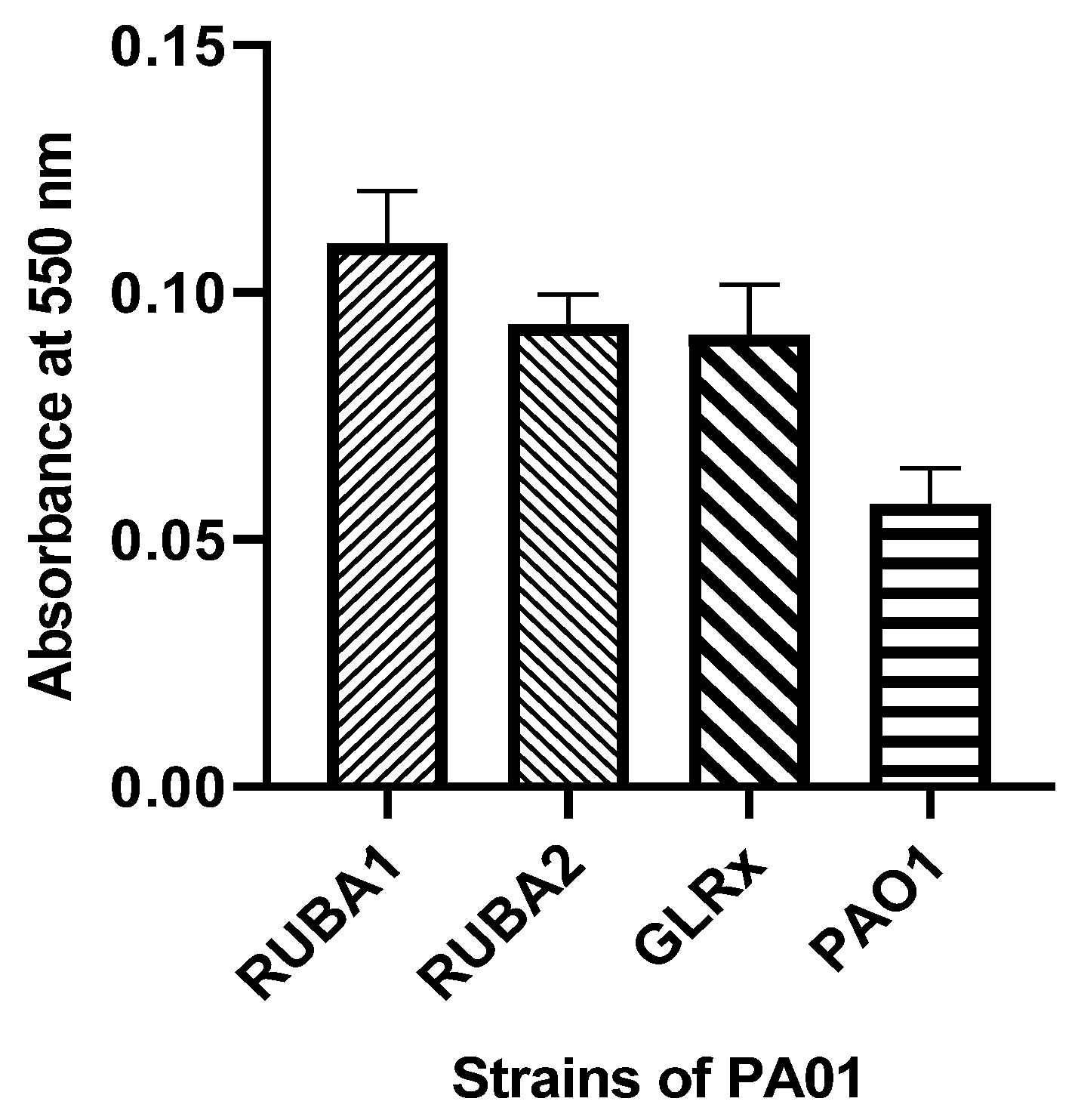
2.5. Biofilm Formation of Rub A1, Rub A2, GLRX and PAO1 under Normal Conditions
2.6. Evaluation of the Production of Proteases LasA in the Strains Rub A1, Rub A2, GLRX and PAO1 under CIP Concentrations
2.7. Rhamnolipids Production in the Strains Rub A1, Rub A2, GLRX and PAO1 under CIP Concentrations
2.8. Pyocyanin Comparison between the Strains Rub A1, Rub A2, GLRX and PAO1 under CIP Concentrations
2.9. Bacteriophage Assay
2.10. Statistics, Data Treatment and Analysis
3. Results
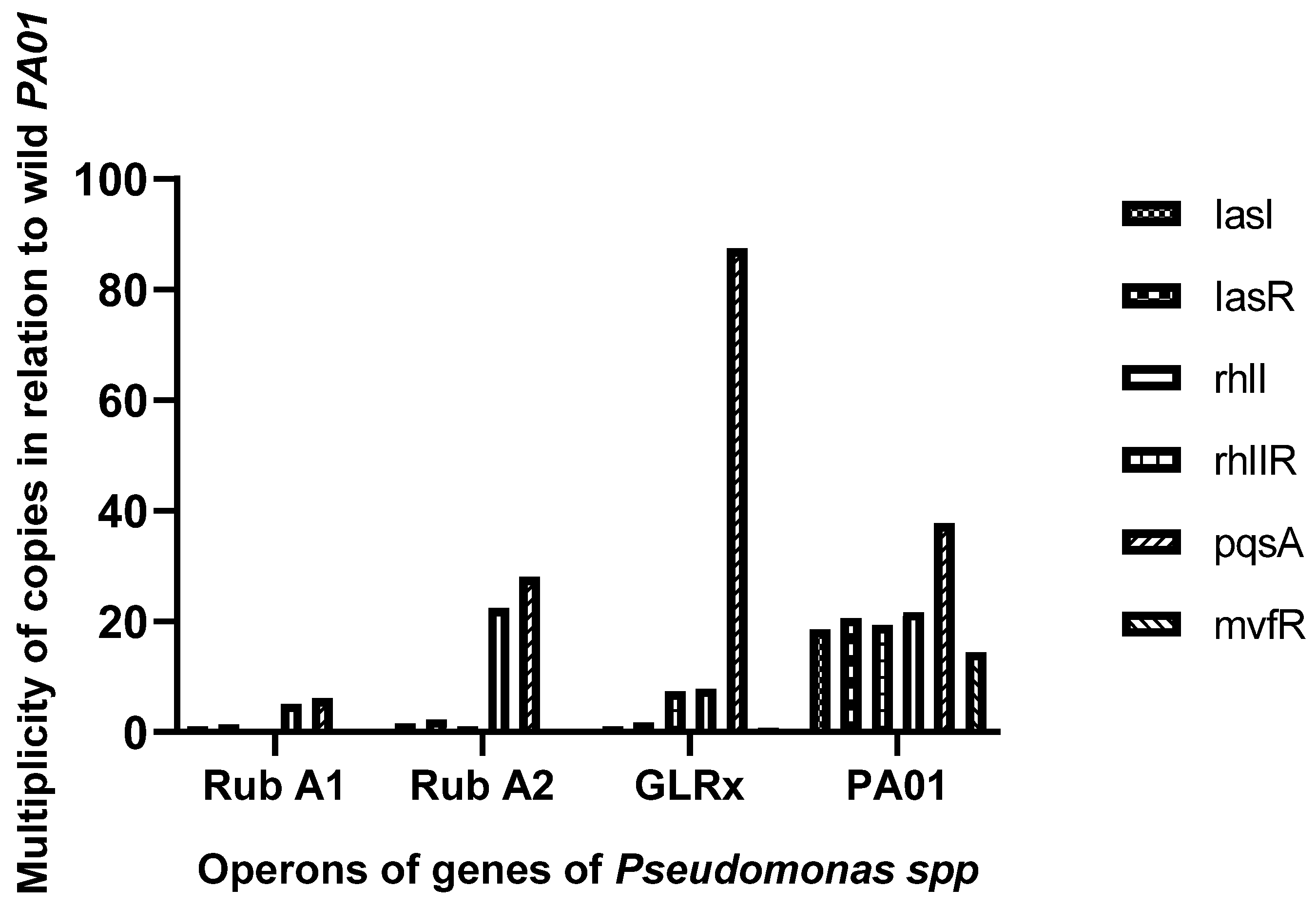
3.1. QS Gene Expression
3.2. Viability Assay of Rub A1, Rub A2, GLRX and PAO1 under CIP Concentrations
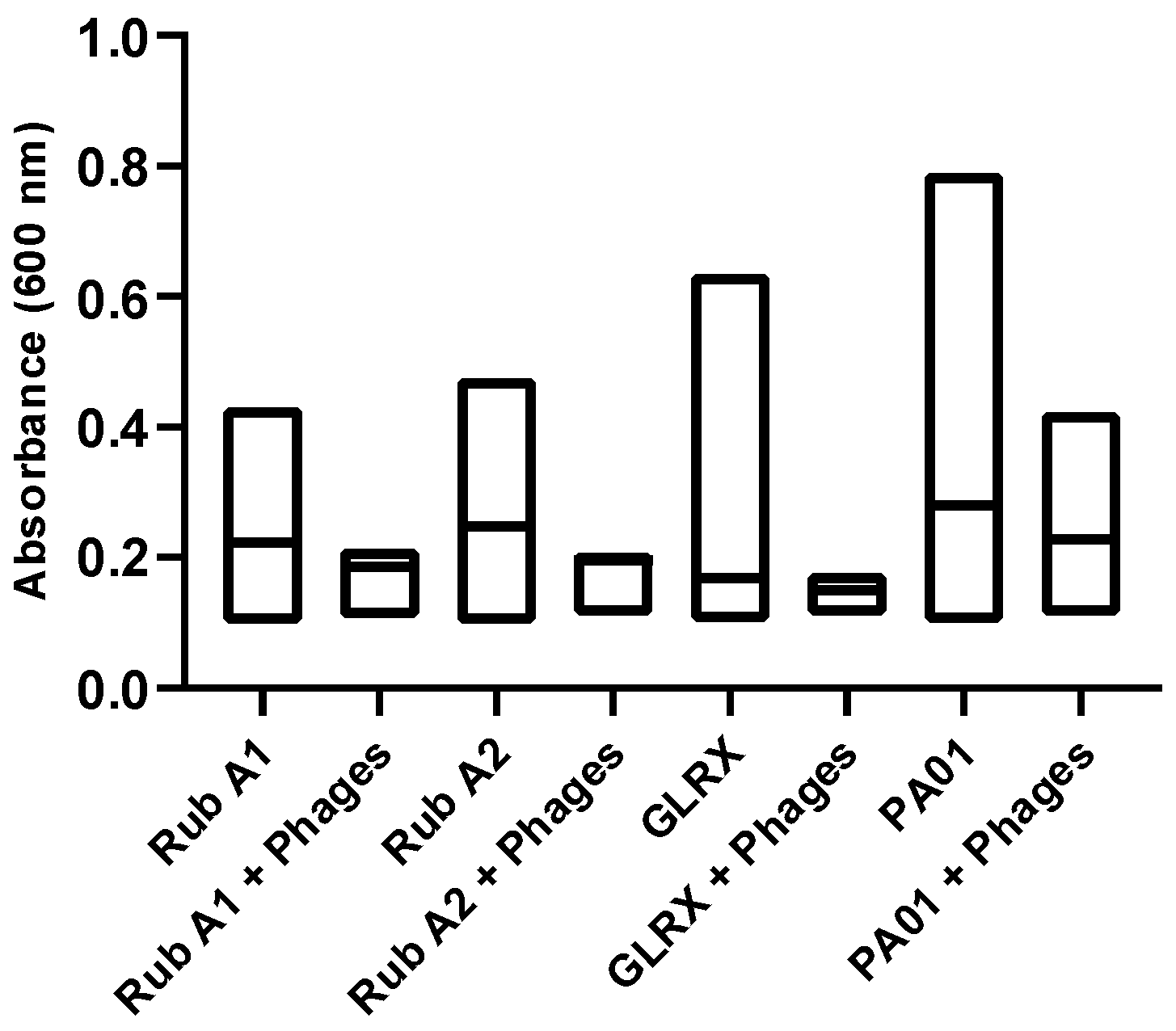
3.3. Biofilm Formation of Rub A1, Rub A2, GLRX and PAO1 under Normal Conditions
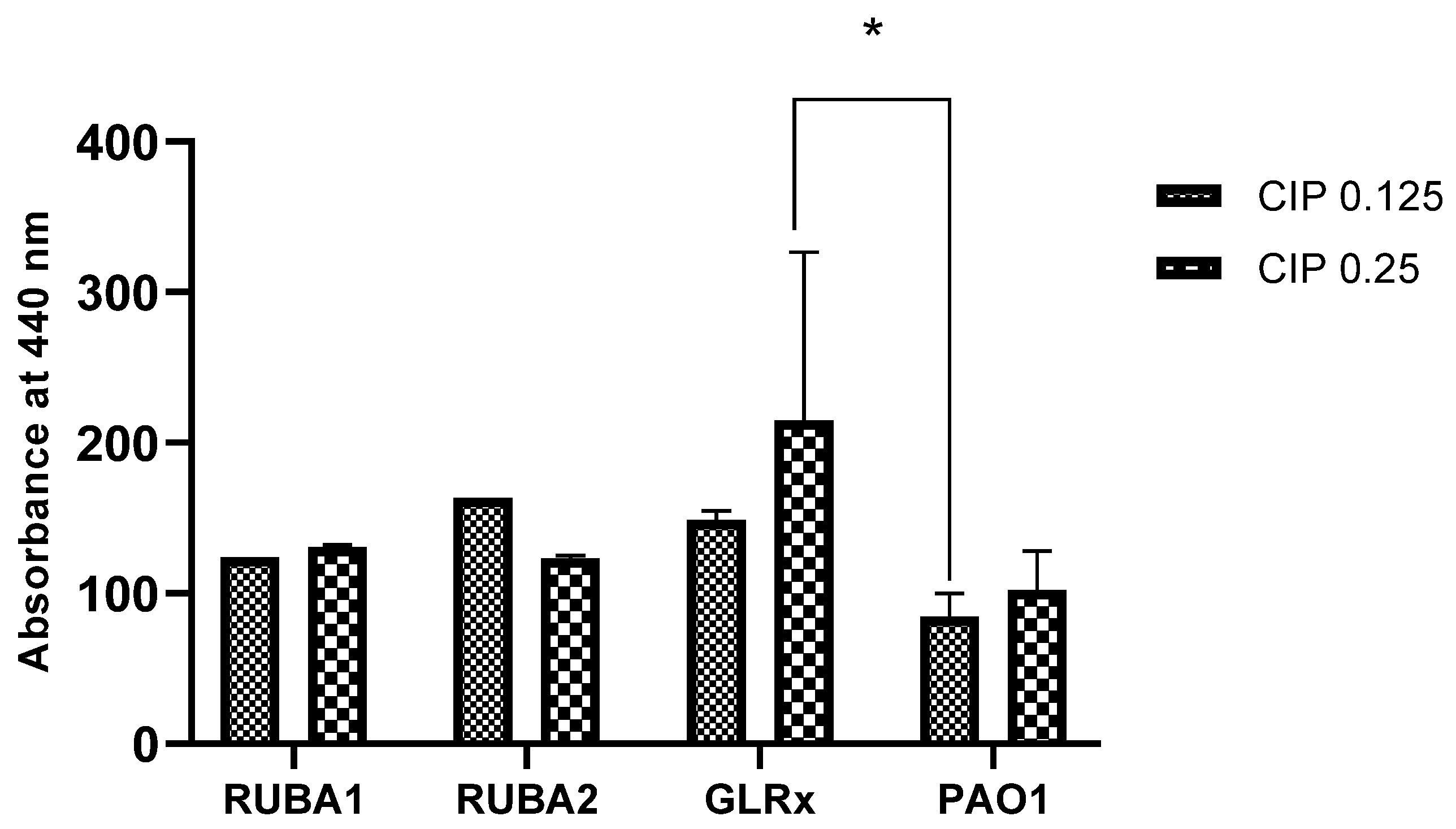
3.4. Evaluation of the Production of Proteases LasA in the Strains Rub A1, Rub A2, GLRX and PAO1 at Two CIP Concentrations
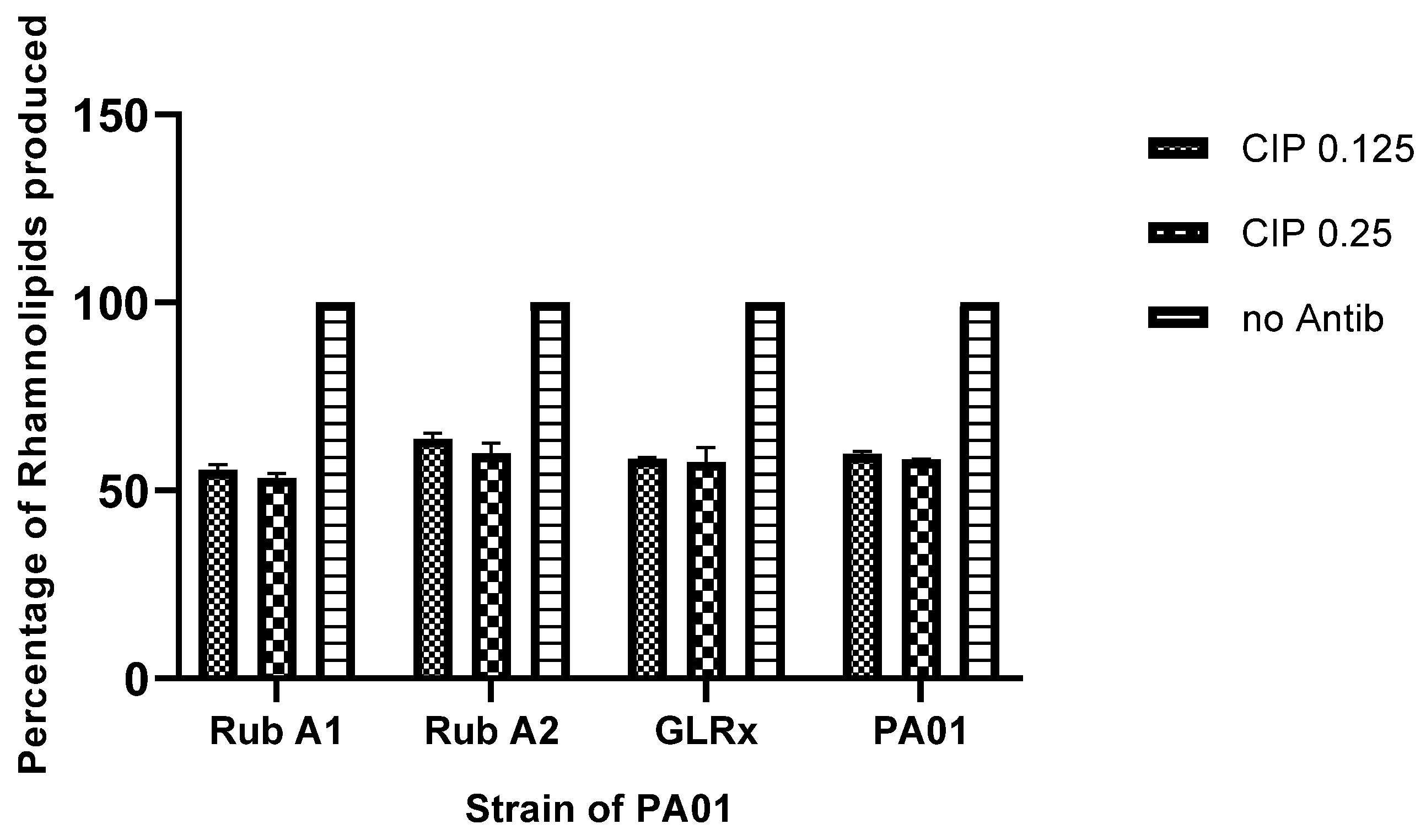
3.5. Rhamnolipids Production in the Strains Rub A1, Rub A2, GLRX and PAO1 under CIP Concentrations
3.6. Pyocyanin Comparison between the Strains Rub A1, Rub A2, GLRX and PAO1 under CIP Concentrations
3.7. Phage Assay
4. Discussion
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Laar, T.A.; Esani, S.; Birges, T.J.; Hazen, B.; Thomas, J.M.; Rawata, M. Pseudomonas aeruginosa gshA Mutant Is Defective in Biofil Formation, Swarming, and Pyocyanin Production. Am. Soc. Microbiol. MSpere 2018, 3, e00155-18. [Google Scholar] [CrossRef]
- Tam, M.; Thi, T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef]
- Thai, T.; Salisbury, B.H.; Zito, P.M. Ciprofloxacin. StatPearls Publishing. 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK535454/ (accessed on 23 June 2022).
- Bassetti, M.; Vena, A.; Croxatto, A.; Righi, E.; Guery, B. How to manage Pseudomonas aeruginosa infections. Drugs Context 2018, 7, 212527. [Google Scholar] [CrossRef]
- Lee, K.; Yoon, S.S. Pseudomonas aeruginosa Biofilm, a Programmed Bacterial Life for Fitness. J. Microbiol. Biotechnol. 2017, 27, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Kostylev, M.; Kim, D.Y.; Smalley, N.E.; Salukhe, I.; Peter Greenberg, E.; Dandekar, A.A. Evolution of the Pseudomonas aeruginosa quorum-sensing hierarchy. Proc. Natl. Acad. Sci. USA 2019, 116, 7027–7032. [Google Scholar] [CrossRef] [PubMed]
- Prateeksha; Bajpai, R.; Yusuf, M.A.; Upreti, D.K.; Gupta, V.K.; Singh, B.N. Endolichenic fungus, Aspergillus quandricinctus of Usnea longissima inhibits quorum sensing and biofilm formation of Pseudomonas aeruginosa PAO1. Microb. Pathog. 2020, 140, 103933. [Google Scholar] [CrossRef]
- Singh, B.N.; Prateeksha; Upreti, D.K.; Singh, B.R.; Defoirdt, T.; Gupta, V.K.; De Souza, A.O.; Singh, H.B.; Barreira, J.C.M.; Ferreira, I.C.F.R.; et al. Bactericidal, quorum quenching and anti-biofilm nanofactories: A new niche for nanotechnologists. Crit. Rev. Biotechnol. 2017, 37, 525–540. [Google Scholar] [CrossRef]
- Williams, P.; Cámara, M. Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: A tale of regulatory networks and multifunctional signal molecules. Curr. Opin. Microbiol. 2009, 12, 182–191. [Google Scholar] [CrossRef]
- Jeong, S.J.; Yoon, S.S.; Bae, I.K.; Jeong, S.H.; Kim, J.M.; Lee, K. Risk factors for mortality in patients with bloodstream infections caused by carbapenem-resistant Pseudomonas aeruginosa: Clinical impact of bacterial virulence and strains on outcome. Diagn. Microbiol. Infect. Dis. 2014, 80, 130–135. [Google Scholar] [CrossRef]
- Malešević, M.; Di Lorenzo, F.; Filipić, B.; Stanisavljević, N.; Novović, K.; Senerovic, L.; Polović, N.; Molinaro, A.; Kojić, M.; Jovčić, B. Pseudomonas aeruginosa quorum sensing inhibition by clinical isolate Delftia tsuruhatensis 11304: Involvement of N-octadecanoylhomoserine lactones. Sci. Rep. 2019, 9, 16465. [Google Scholar] [CrossRef]
- Ahmed, S.A.K.S.; Rudden, M.; Smyth, T.J.; Dooley, J.S.G.; Marchant, R.; Banat, I.M. Natural quorum sensing inhibitors effectively downregulate gene expression of Pseudomonas aeruginosa virulence factors. Appl. Microbiol. Biotechnol. 2019, 103, 3521–3535. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Li, X.-H.; Kim, S.-K.; Lee, J.-H. Post-secretional activation of Protease IV by quorum sensing in Pseudomonas aeruginosa. Sci. Rep. 2017, 7, 4416. [Google Scholar] [CrossRef] [PubMed]
- Lichtenberg, M.; Jakobsen, T.H.; Kühl, M.; Bjarnsholt, T. The structure—Function relationship of Pseudomonas aeruginosa in infections and its influence on the microenvironment. FEMS Microbiol. Rev. 2022, 46, fuac018. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, A.A.; Kamer, A.M.A.; Al-Monofy, K.B.; Al-Madboly, L.A. Pseudomonas aeruginosa’s greenish-blue pigment pyocyanin: Its production and biological activities. Microb. Cell Factories 2023, 22, 110. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Hao, S.; Zhao, L.; Shi, F.; Ye, G.; Zou, Y.; Song, X.; Li, L.; Yin, Z.; He, X.; et al. Paeonol Attenuates Quorum-Sensing Regulated Virulence and Biofilm Formation in Pseudomonas aeruginosa. Front. Microbiol. 2021, 12, 692474. [Google Scholar] [CrossRef]
- Hagelueken, G.; Wiehlmann, L.; Adams, T.M.; Kolmar, H.; Heinz, D.W.; Tu, B. Crystal structure of the electron transfer complex rubredoxin—Rubredoxin reductase of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2007, 104, 12276–12281. [Google Scholar] [CrossRef]
- Wiehlmann, L.; Urbanke, C.; Adams, T.M.; Munder, A.; Tümmler, B. The ancient rubredoxin system: An efficient defense mechanism of pseudomonas aeruginosa during infections. Eur. Respir. J. 2014, 44, 484. [Google Scholar]
- Saninjuk, K.; Romsang, A.; Duang-Nkern, J.; Wongsaroj, L.; Leesukon, P.; Dubbs, J.M.; Vattanaviboon, P.; Mongkolsuk, S. Monothiol Glutaredoxin Is Essential for Oxidative Stress Protection and Virulence in Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2023, 89, e0171422. [Google Scholar] [CrossRef]
- Napper, K.R.; Leeper, T.C.; Khattri, R.; Morris, D.; Davis, C. Drug Interactions with Glutaredoxin Orthologues. Bachelor’s Thesis, The Honors College, University of Akron, Akron, OH, USA, 2015. [Google Scholar]
- Chegini, Z.; Khoshbayan, A.; Moghadam, M.T.; Farahani, I.; Jazireian, P. Bacteriophage therapy against Pseudomonas aeruginosa biofilms: A review. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 45. [Google Scholar] [CrossRef]
- Liu, R.; Han, G.; Li, Z.; Cun, S.; Hao, B.; Zhang, J.; Liu, X. Bacteriophage therapy in aquaculture: Current status and future challenges. Folia Microbiol. 2022, 67, 573–590. [Google Scholar] [CrossRef]
- Uyttebroek, S.; Chen, B.; Onsea, J.; Ruythooren, F.; Debaveye, Y.; Devolder, D.; Spriet, I.; Depypere, M.; Wagemans, J.; Lavigne, R.; et al. Safety and efficacy of phage therapy in difficult-to-treat infections: A systematic review. Lancet 2022, 22, e208–e220. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, S.N.; Manohar, P.; Sukumaran, S.; Sadagopan, S.; Loh, B.; Leptihn, S.; Nachimuthu, R. Antibacterial efficacy of lytic phages against multidrug-resistant Pseudomonas aeruginosa infections in bacteraemia mice models. BMC Microbiol. 2022, 22, 187. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, M.A.; Alwood, A.; Thaipisuttikul, I.; Spencer, D.; Haugen, E.; Ernst, S.; Will, O.; Kaul, R.; Raymond, C.; Levy, R.; et al. Comprehensive transposon mutant library of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2003, 100, 14339–14344. [Google Scholar] [CrossRef] [PubMed]
- Malgaonkar, A.; Nair, M. Quorum sensing in Pseudomonas aeruginosa mediated by RhlR is regulated by a small RNA PhrD. Sci. Rep. 2019, 9, 11. [Google Scholar] [CrossRef]
- Franke, J.D.; Braverman, A.L.; Cunningham, A.M.; Eberhard, E.E.; Perry, G.A. Erythrosin B: A versatile colorimetric and fluorescent vital dye for bacteria. BioTechniques 2019, 68, 7–13. [Google Scholar] [CrossRef]
- Gambello, M.J.; Kaye, S.; Iglewski, B.H. LasR of Pseudomonas aeruginosa is a transcriptional activator of the alkaline protease gene (apr) and an enhancer of exotoxin A expression. Infect. Immun. 1993, 61, 1180–1184. [Google Scholar] [CrossRef]
- Ochsner, U.A.; Koch, A.K.; Fiechter, A.; Reiser, J. Isolation and characterization of a regulatory gene affecting rhamnolipid biosurfactant synthesis in Pseudomonas aeruginosa. J. Bacteriol. 1994, 176, 2044–2054. [Google Scholar] [CrossRef]
- Adamski, M.G.; Gumann, P.; Baird, A.E. A Method for Quantitative Analysis of Standard and High-Throughput qPCR Expression Data Based on Input Sample Quantity. PLoS ONE 2014, 9, e103917. [Google Scholar] [CrossRef]
- Balasubramanian, D.; Mathee, K. Comparative transcriptome analyses of Pseudomonas aeruginosa. Hum. Genom. 2009, 3, 349. [Google Scholar] [CrossRef]
- Shome, S.; Talukdar, A.D.; Nath, R.; Tewari, S. Curcumin-ZnO nanocomposite mediated inhibition of Pseudomonas aeruginosa biofilm and its mechanism of action. J. Drug Deliv. Sci. Technol. 2023, 81, 104301. [Google Scholar] [CrossRef]
- Rao, V.A. Iron Chelators with Topoisomerase-Inhibitory Activity and Their Anticancer Applications. Antioxid. Redox Signal. 2013, 18, 930–955. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.; Turner, K.E.; Kirienko, N.V. PqsA Promotes Pyoverdine Production via Biofilm Formation. Pathogens 2018, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Cheng, J.; Wang, Y.; Shen, X. The Pseudomonas Quinolone Signal (PQS): Not Just for Quorum Sensing Anymore. Front. Cell. Infect. Microbiol. 2018, 8, 230. [Google Scholar] [CrossRef]
- Shahab, M.; Danial, M.; Khan, T.; Liang, C.; Duan, X.; Wang, D.; Gao, H.; Zheng, G. In Silico Identification of Lead Compounds for Pseudomonas Aeruginosa PqsA Enzyme: Computational Study to Block Biofilm Formation. Biomedicines 2023, 11, 961. [Google Scholar] [CrossRef] [PubMed]
- Storz, M.P.; Maurer, C.K.; Zimmer, C.; Wagner, N.; Brengel, C.; de Jong, J.C.; Lucas, S.; Müsken, M.; Häussler, S.; Steinbach, A.; et al. Validation of PqsD as an Anti-biofilm Target in Pseudomonas aeruginosa by Development of Small-Molecule Inhibitors. J. Am. Chem. Soc. 2012, 134, 16143–16146. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.C.; Ohman, D.E. Efficient production and processing of elastase and LasA by Pseudomonas aeruginosa require zinc and calcium ions. J. Bacteriol. 1992, 174, 4140–4147. [Google Scholar] [CrossRef]
- Wittgens, A.; Kovacic, F.; Müller, M.M.; Gerlitzki, M.; Santiago-schübel, B.; Hofmann, D.; Tiso, T.; Blank, L.M.; Henkel, M.; Hausmann, R.; et al. Novel insights into biosynthesis and uptake of rhamnolipids and their precursors. Appl. Microbiol. Biotechnol. 2017, 101, 2865–2878. [Google Scholar] [CrossRef]
- Kart, D.; Reçber, T.; Nemutlu, E.; Sagiroglu, M. Sub-Inhibitory Concentrations of Ciprofloxacin Alone and Combinations with Plant-Derived Compounds against P. aeruginosa Biofilms and Their Effects on the Metabolomic Profile of P. aeruginosa Biofilms. Antibiotics 2021, 10, 414. [Google Scholar] [CrossRef]
- Alatraktchi, F.A.; Svendsen, W.E.; Molin, S. Electrochemical Detection of Pyocyanin as a Biomarker for Pseudomonas aeruginosa: A Focused Review. Sensors 2020, 20, 5218. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Abbreviation | Sources | Preparation Specifications |
|---|---|---|---|
| Tryptic soy broth | TSB | Liofilchem | 30 g in 1000 mL distilled water |
| Ciprofloxacin | CIP | TCI (8572-33-1) | 0.25; 0.50 µg of antibiotic for one mL of sterile water |
| Crystal violet | CV | Reagentkit | 0.1% of the dye crystal violet diluted in water |
| Acetic acid 30% | AC | Chem-Lab (64-19-7) | 30% de pure acetic acid diluted in sterile water |
| Erythrosine B | EB | TCI (16423-68-0) | 0.1% (w/v) in 10 mM of TRIS, pH 7.5 |
| Phosphate-buffered saline | PBS | OXOID | Ready to use |
| Azocasein | AZO | SIGMA (102110-74-7) | 0.8% of azocasein in K2HPO4 50 mM and pH adjusted to 7 |
| Hydrogen chloride | HCL | Chem-Lab (7627-01-0) | 0.2 M, 1 N and 1.5 M in sterile water |
| Sodium hydroxide | NaOH | JMGS (1310-73-2) | 1 N diluted in sterile water |
| Bovine serum albumin | BSA | FMUP | Ready to use solution |
| Reaction tampon | - | Fisher bioreagents BP152-1 Fisher bioreagents (77-86-1) | Tris-HCl 10 mM with CaCl2 0.1 mM solution |
| Congo red | CR | SIGMA | 2 mg in 1000 mL of water |
| Ethylenediamine tetra acetic acid | EDTA | Chem-Lab (6381-92-6) | 0.5 M dissolved in water |
| Chloroform | CHCl3 | MERCK | Ready to use |
| Methylene blue | MB | Chem-Lab NV (61-73-4) | 1 g in 1000 mL of methylene blue prepared in 10 mM borax buffer pH 10.5 and stabilized by adjusting pH to 5.5 |
| Gene | Primer Direction | Sequence (5′–3′) | Amplicon Size (bp) |
|---|---|---|---|
| lasI | Forward | 5-GCGTGCTCAAGTGTTCAAGG-3 | 125 |
| Reverse | 5-GGGCTTCAGGAGTATCTTCCTGG-3 | ||
| lasR | Forward | 5-GGAGTGGAGCGCCATCCTGCAG-3 | 127 |
| Reverse | 5-GGCGGCCGGGTAGTTGCCGACG-3 | ||
| rhlI | Forward | 5-CCATCCGCAAACCCGCTACATC-3 | 151 |
| Reverse | 5-CTCCCAGACCGACGGATCGCTCGGC-3 | ||
| rhlR | Forward | 5-GGGCGTGTTCGCCGTCCTGG-3 | 143 |
| Reverse | 5-GGTATCGCTCCAGCCAGGCCTTG-3 | ||
| pqsA | Forward | 5-CCGGACCTACATTCTCTCCC-3 | 182 |
| Reverse | 5-CGATATCGGCCAGGGCCTGC-3 | ||
| mvfR | Forward | 5-GTCGGGACGGCTACAAGGTCG-3 | 129 |
| Reverse | 5-GATTGCGCGGACCCTTGTTGAG-3 | ||
| rpsL | Forward | 5-GCAACTATCAACCAGCTGGTG-3 | 231 |
| Reverse | 5-GCTGTGCTCTTGCAGGTTGTG-3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sá, S.; Silva, C.; Dias, M.C.; Veiga, M.; Lopes, S.; Fernandes, R.; Rocha, A.C.; Pais, P.J.; Oliveira, M.; Mendes, J.; et al. Virulence-Linked Mutations in Rubredoxin Reductase and Glutaredoxin: Impact on Antibiotic Susceptibility and Phage Therapy in Pseudomonas aeruginosa. Appl. Sci. 2023, 13, 11918. https://doi.org/10.3390/app132111918
Sá S, Silva C, Dias MC, Veiga M, Lopes S, Fernandes R, Rocha AC, Pais PJ, Oliveira M, Mendes J, et al. Virulence-Linked Mutations in Rubredoxin Reductase and Glutaredoxin: Impact on Antibiotic Susceptibility and Phage Therapy in Pseudomonas aeruginosa. Applied Sciences. 2023; 13(21):11918. https://doi.org/10.3390/app132111918
Chicago/Turabian StyleSá, Sara, Carina Silva, Maria Clara Dias, Marlene Veiga, Sofia Lopes, Ruben Fernandes, Ana Catarina Rocha, Patrick J. Pais, Marco Oliveira, João Mendes, and et al. 2023. "Virulence-Linked Mutations in Rubredoxin Reductase and Glutaredoxin: Impact on Antibiotic Susceptibility and Phage Therapy in Pseudomonas aeruginosa" Applied Sciences 13, no. 21: 11918. https://doi.org/10.3390/app132111918
APA StyleSá, S., Silva, C., Dias, M. C., Veiga, M., Lopes, S., Fernandes, R., Rocha, A. C., Pais, P. J., Oliveira, M., Mendes, J., Novais, G., Luís, C., Gestoso, Á., Macedo, J. M., Martins-Mendes, D., Pereira, A. C., & Baylina, P. (2023). Virulence-Linked Mutations in Rubredoxin Reductase and Glutaredoxin: Impact on Antibiotic Susceptibility and Phage Therapy in Pseudomonas aeruginosa. Applied Sciences, 13(21), 11918. https://doi.org/10.3390/app132111918