
Prospects of Computer-Aided Molecular Design of Coumarins as Ecotoxicologically Safe Plant Protection Agents

 , , ,
, , ,
Featured Application
Abstract
1. Introduction
2. Naturally Occurring Coumarins and Their Role in Plants
2.1. Biosynthesis and Distribution of Coumarins in Nature
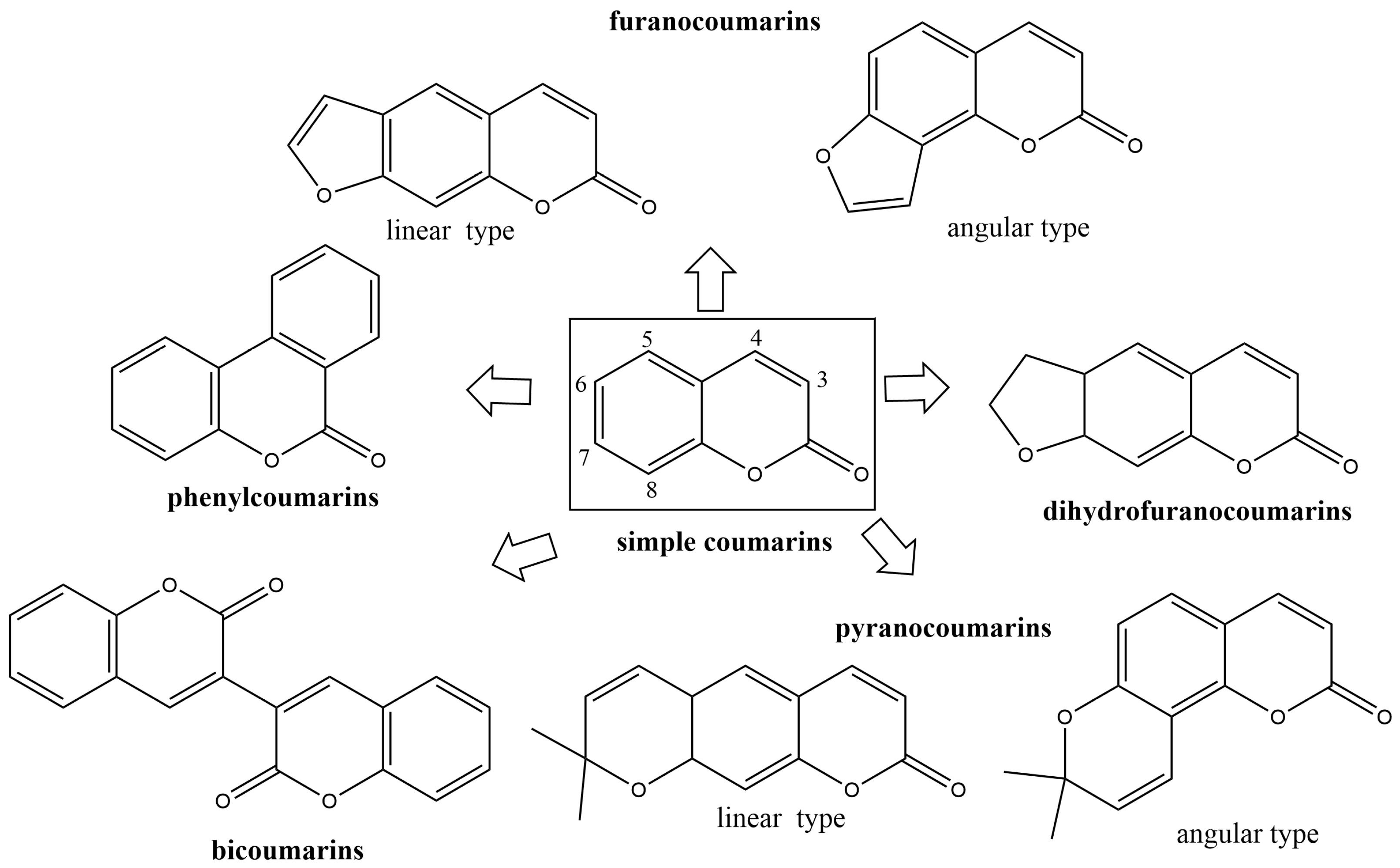
2.2. Classification of Naturally Occurring Coumarins and Their Role in Plant Protection
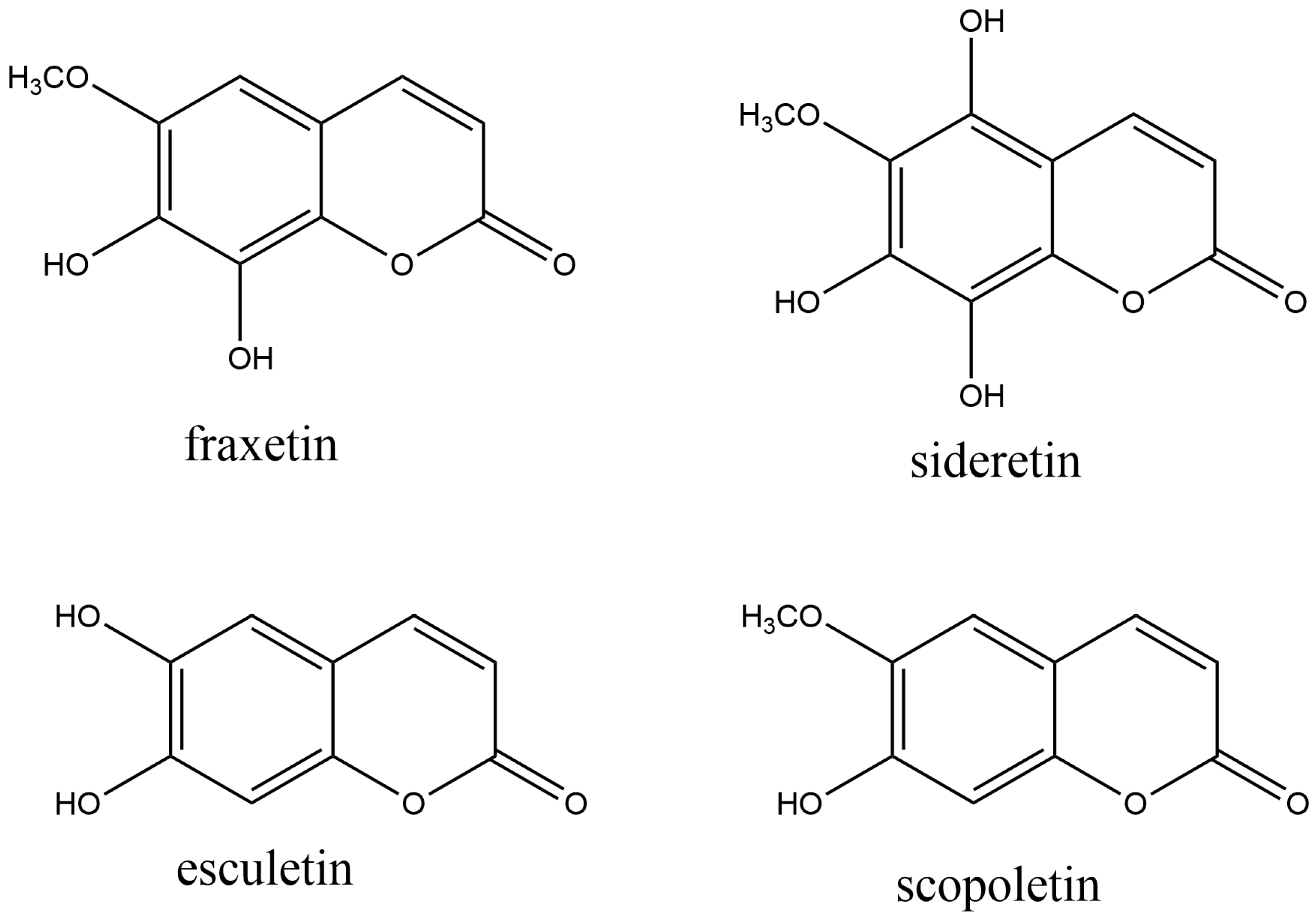
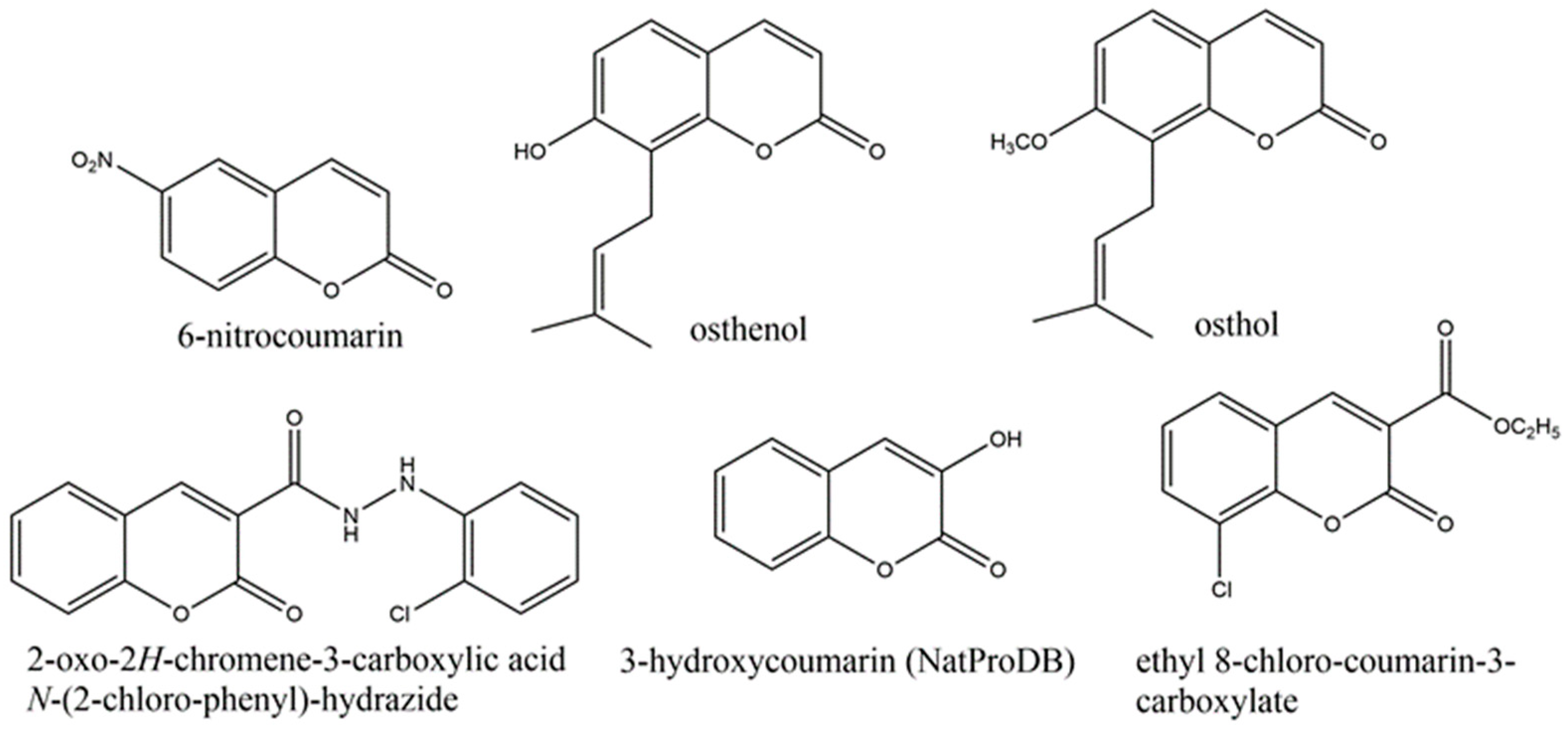
2.2.1. Simple Coumarins
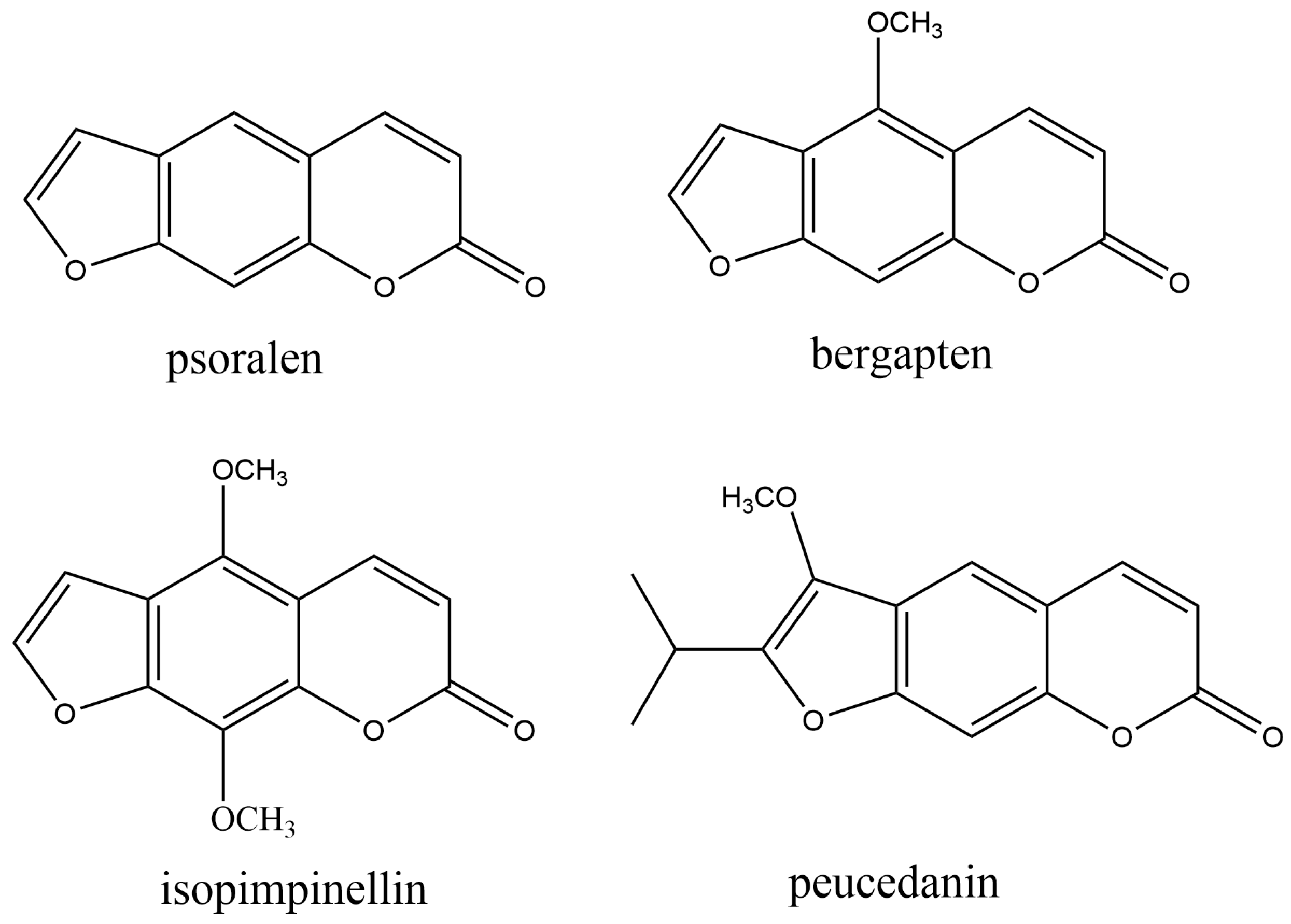
2.2.2. Furanocoumarins
2.2.3. Dihydrofuranocoumarins
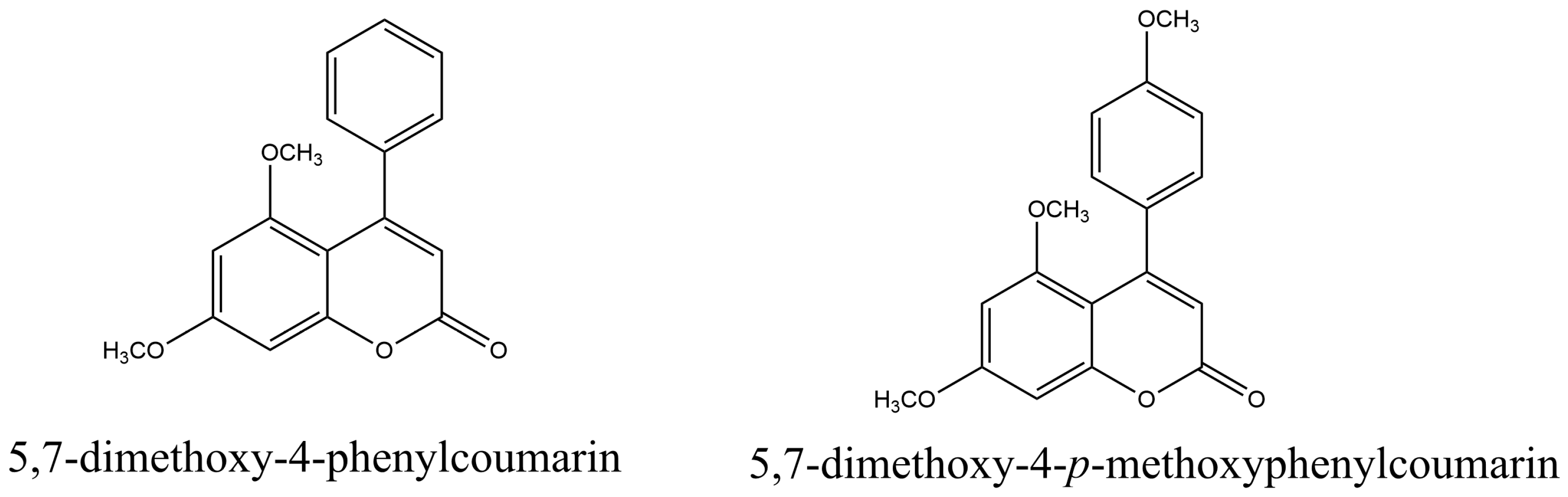
2.2.4. Phenylcoumarins
2.2.5. Pyranocoumarins
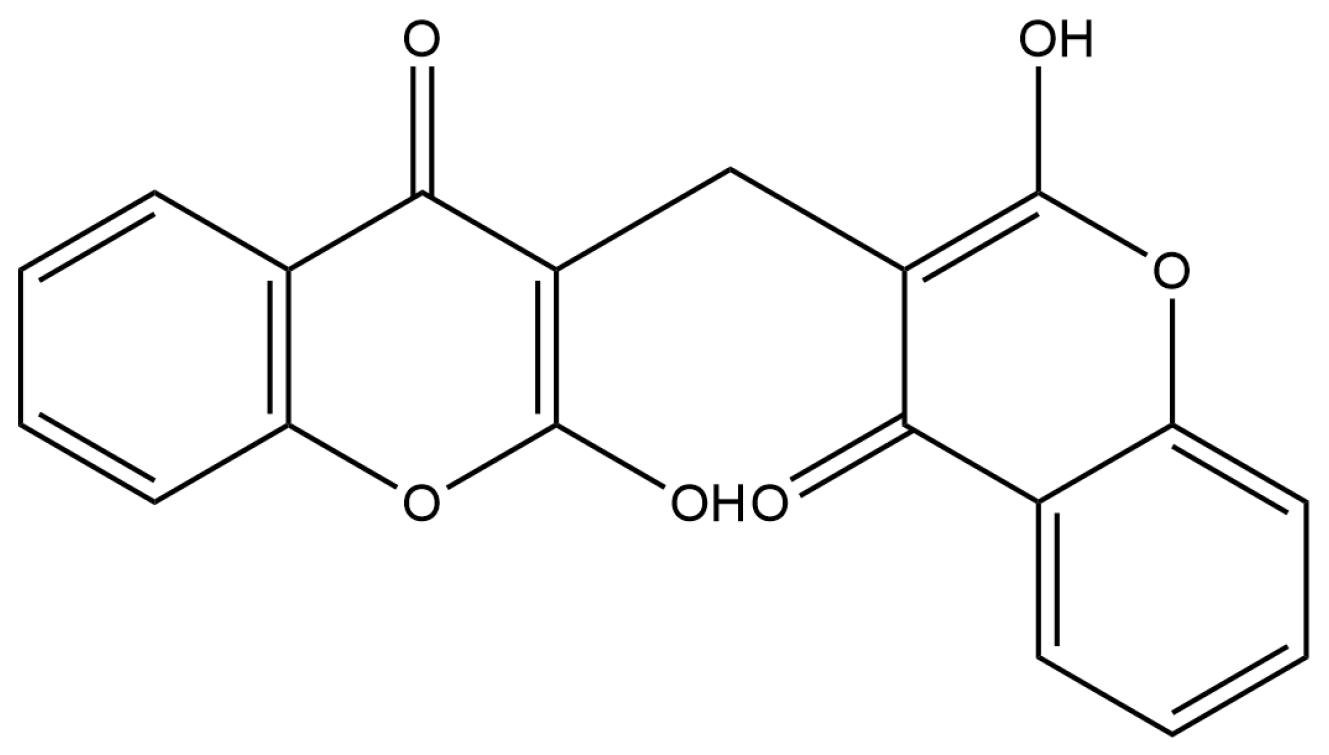
2.2.6. Bicoumarines
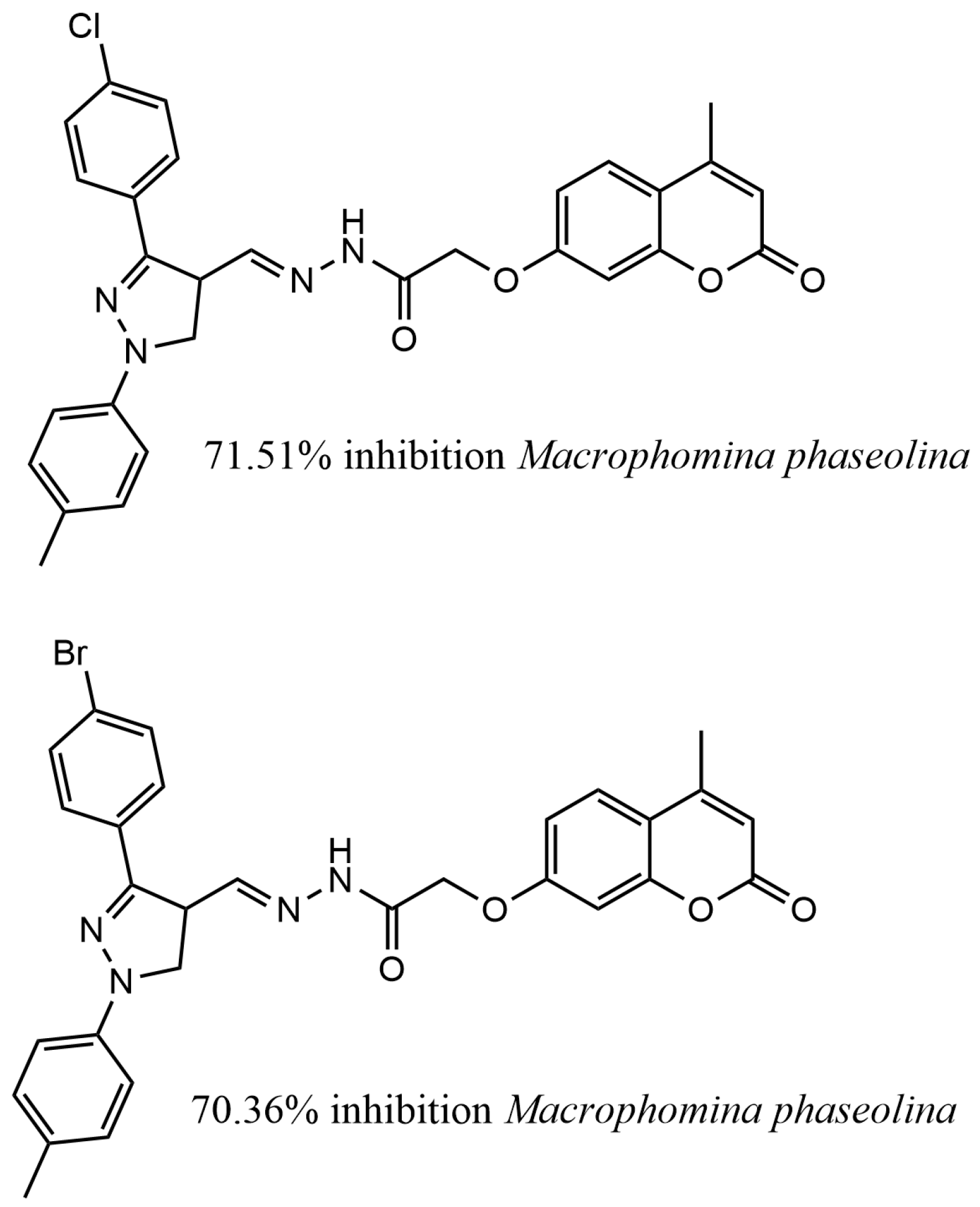
3. Synthetic Coumarin Derivatives in Plant Protection
4. Computer-Aided Molecular Design (CAMD) of Coumarins for Potential Plant Protection Application
4.1. QSAR
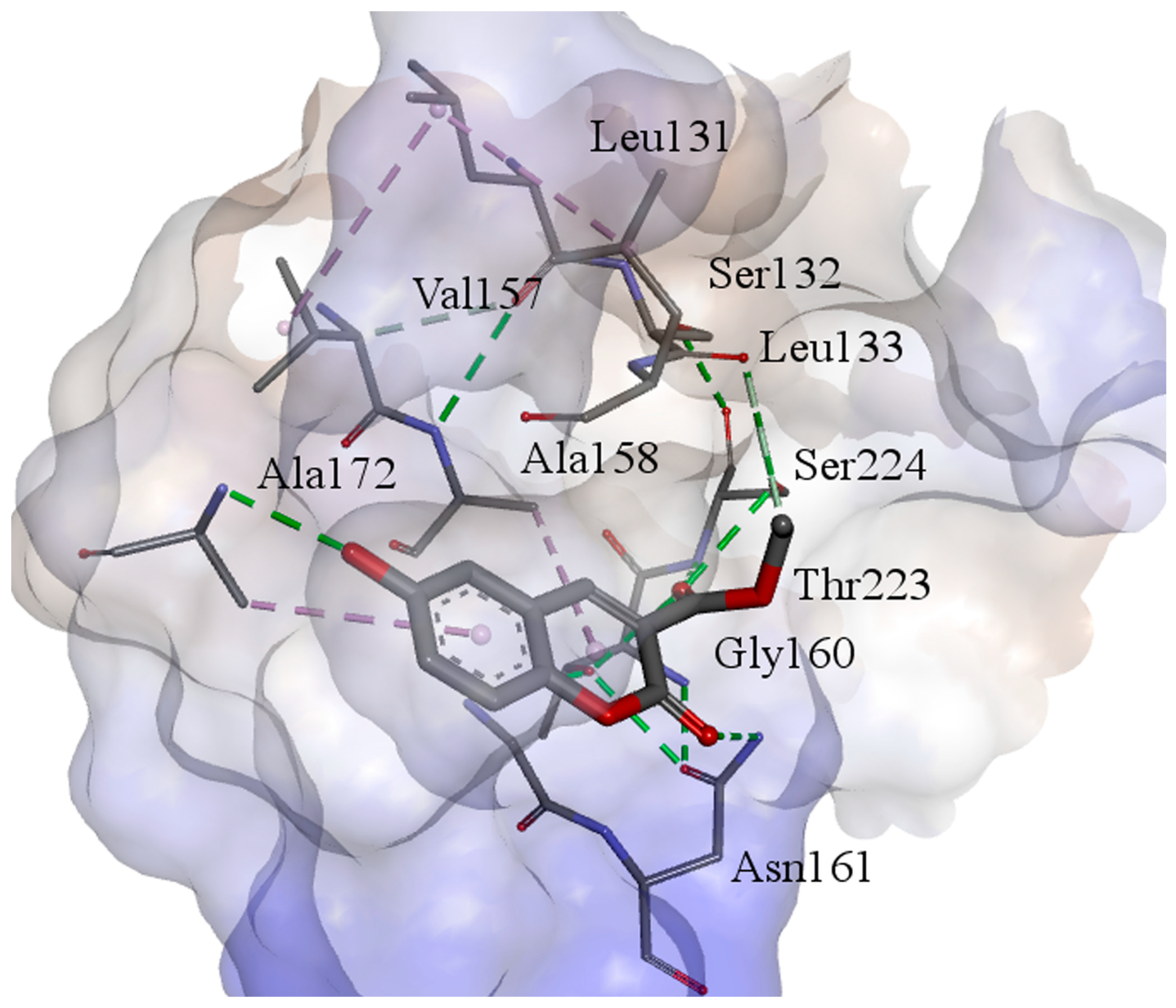
4.2. Molecular Docking, and Molecular Dynamics (MD) Simulations
4.3. Quantitative Estimation of Pesticide-Likeness Properties
4.4. Environmental and Health Hazards Properties of Coumarins
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AB | number of aromatic bonds |
| AChE | acetylcholinesterase |
| ADMET | absorption, distribution, metabolism, excretion, and toxicity |
| ADMET | absorption, distribution, metabolism, excretion, and toxicity |
| BGLU | beta-glucosidase |
| CAMD | computer-aided molecular design |
| CoA | coenzyme A |
| CoMFA | comparative molecular field analysis |
| CoMSIA | comparative molecular similarity indices analysis |
| CS | chitin synthase |
| ECHA | European Chemicals Agency |
| ECTOC | European Centre for Ecotoxicology and Toxicology of Chemicals |
| EFSA | European Food Safety Authority |
| EPA | Environmental Protection Agency |
| EPN | entomopathogenic nematodes |
| FNN | feedforward neural network |
| GEP | gene expression programming |
| HBA | hydrogen-bond acceptor |
| HBD | hydrogen-bond donor |
| HTS | high throughput screening |
| JCR | Joint Research Centre |
| k-NN | k-nearest neighbor |
| LLR | local lazy regression |
| MD | molecular dynamics |
| MIC | minimal inhibitory concentration |
| MLR | multiple linear regression |
| MoRSE | molecular representation of structures based on electronic diffraction |
| MW | molecular weight |
| NN | neural networks |
| OECD | Organisation for Economic Co-operation and Development |
| 2OGD | 2-oxoglutarate-dependent dioxygenase |
| PLS | partial least squares |
| PPN | plant parasitic nematode |
| PPR | project pursuit regression |
| QSAR | quantitative structure-activity relationships |
| RB | number of rotatable bonds |
| REACH | registration, evaluation and authorization of chemicals |
| SAR | structure-activity relationship |
| SBL | soil borne legacy |
| SVM | support vector machine |
| WHIM | weighted holistic invariant molecular |
| TcPMCA1 | Tetranychus cinnabarinus plasma membrane Ca2+-ATPase 1 |
References
- Sánchez-Bayo, F.; Tennekes, H.A. Environmental Risk Assessment of Agrochemicals—A Critical Appraisal of Current Approaches. In Toxicity and Hazard of Agrochemicals; Larramendy, M.L., Soloneski, S., Eds.; InTech: London, UK, 2015. [Google Scholar] [CrossRef]
- Li, Z. A disease-specific screening-level modeling approach for assessing the cancer risks of pesticide mixtures. Chemosphere 2022, 286, 131811. [Google Scholar] [CrossRef]
- Li, Z. Prioritizing agricultural pesticides to protect human health: A multi-level strategy combining life cycle impact and risk assessments. Ecotoxicol. Environ. Saf. 2022, 242, 113869. [Google Scholar] [CrossRef] [PubMed]
- Polyxeni, N.-S.; Sotirios, M.; Chrysanthi, K.; Panagiotis, S.; Luc, H. Pesticides and human health: The urgent need for a new concept in agriculture. Front. Public Health 2016, 4, 148. [Google Scholar] [CrossRef]
- Smith, K.; Evans, D.A.; El-Hiti, G.A. Role of modern chemistry in sustainable arable crop protection. Philos. Trans. R. Soc. B 2008, 363, 623–637. [Google Scholar] [CrossRef]
- European Parliament. Directive 2009/128/EC of the European Parliament and of the Council of Establishing a Framework for Community Action to Achieve the Sustainable Use of Pesticides. 21 October 2009. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=celex%3A32009L0128 (accessed on 4 November 2022).
- Hussain, M.I.; Qamar Abbas, S.; Reigosa, M.J. Activities and novel applications of secondary metabolite coumarins. Planta Daninha 2017, 35, e017174040. [Google Scholar] [CrossRef]
- Kadhum, A.A.H.; Al-Amiery, A.A.; Musa, A.Y.; Mohamad, A.B. The antioxidant activity of new coumarin derivatives. Int. J. Mol. Sci. 2011, 12, 5747–5761. [Google Scholar] [CrossRef]
- Molnar, M.; Komar, M.; Brahmbhatt, H.; Babić, J.; Jokić, S.; Rastija, V. Deep eutectic solvents as convenient media for synthesis of novel coumarinyl schiff bases and their QSAR studies. Molecules 2017, 22, 1482. [Google Scholar] [CrossRef]
- Al-Majedy, Y.K.; Al-Duhaidahawi, D.L.; Al-Azawi, K.F.; Al-Amiery, A.A.; Kadhum, A.A.H.; Mohamad, A.B. Coumarins as potential antioxidant agents complemented with suggested mechanisms and approved by molecular modeling studies. Molecules 2016, 21, 135. [Google Scholar] [CrossRef]
- Sahoo, C.R.; Sahoo, J.; Mahapatra, M.; Lenka, D.; Sahu, P.K.; Dehury, B.; Padhy, R.N.; Paidesetty, S.K. Coumarin derivatives as promising antibacterial agent(s). Arab. J. Chem. 2021, 14, 102922. [Google Scholar] [CrossRef]
- Bhagat, K.; Bhagat, J.; Gupta, M.K.; Singh, J.V.; Gulati, H.K.; Singh, A.; Kaur, K.; Kaur, G.; Sharma, S.; Rana, A.; et al. Design, synthesis, antimicrobial evaluation, and molecular modeling studies of novel indolinedione–coumarin molecular hybrids. ACS Omega 2019, 4, 8720–8730. [Google Scholar] [CrossRef]
- Cheke, R.S.; Patel, H.M.; Patil, V.M.; Ansari, I.A.; Ambhore, J.P.; Shinde, S.D.; Kadri, A.; Snoussi, M.; Adnan, M.; Kharkar, P.S.; et al. Molecular insights into coumarin analogues as antimicrobial agents: Recent developments in drug discovery. Antibiotics 2022, 11, 566. [Google Scholar] [CrossRef] [PubMed]
- Završnik, D.; Špirtović-Halilović, S.; Softić, D. Synthesis, structure and antibacterial activity of 3-substituted derivatives of 4-hydroxycoumarin. Period. Biol. 2011, 133, 93–97. Available online: https://hrcak.srce.hr/67271 (accessed on 2 January 2023).
- Eustáquio, A.S.; Gust, B.; Luft, T.; Li, S.-M.; Chater, K.F.; Heide, L. Clorobiocin biosynthesis in Streptomyces: Identification of the halogenase and generation of structural analogs. Chem. Biol. 2003, 10, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Süzgeç-Selçuk, S.; Dikpınar, T. Phytochemical evaluation of the Ferulago genus and the pharmacological activities of its coumarin constituents. J. Herb. Med. 2021, 21, 100415. [Google Scholar] [CrossRef]
- Deryabin, D.; Inchagova, K.; Rusakova, E.; Duskaev, G. Coumarin’s anti-quorum sensing activity can be enhanced when combined with other plant-derived small molecules. Molecules 2021, 26, 208. [Google Scholar] [CrossRef]
- De Araújo, R.S.A.; Guerra, F.Q.S.; de O Lima, E.; De Simone, C.A.; Tavares, J.F.; Scotti, L.; Scotti, M.T.; De Aquino, T.M.; De Moura, R.O.; Mendonça, F.J.B.; et al. Synthesis, structure-activity relationships (SAR) and in silico studies of coumarin derivatives with antifungal activity. Int. J. Mol. Sci. 2013, 14, 1293–1309. [Google Scholar] [CrossRef]
- Lemos, A.S.O.; Florêncio, J.R.; Pinto, N.C.C.; Campos, L.M.; Silva, T.P.; Grazul, R.M.; Pinto, P.F.; Tavares, G.D.; Scio, E.; Apolônio, A.C.M.; et al. Antifungal activity of the natural coumarin scopoletin against planktonic cells and biofilms from a multidrug-resistant Candida tropicalis strain. Front. Microbiol. 2020, 11, 1525. [Google Scholar] [CrossRef]
- Ayine-Tora, D.M.; Kingsford-Adaboh, R.; Asomaning, W.A.; Harrison, J.J.E.K.; Mills-Robertson, F.C.; Bukari, Y.; Sakyi, P.O.; Kaminta, S.; Reynisson, J. Coumarin Antifungal lead compounds from Millettia thonningii and their predicted mechanism of action. Molecules 2016, 21, 1369. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Chen, Q.; Zhang, Y.; Liang, C. Coumarin-based derivatives with potential anti-HIV activity. Fitoterapia 2021, 150, 104863. [Google Scholar] [CrossRef] [PubMed]
- Hamoda, A.M.; Fayed, B.; Ashmawy, N.S.; El-Shorbagi, A.A.; Hamdy, R.; Soliman, S.S.M. Marine sponge is a promising natural source of anti-SARS-CoV-2 scaffold. Front. Pharmacol. 2021, 12, 666664. [Google Scholar] [CrossRef]
- Hu, Y.-Q.; Xu, Z.; Zhang, S.; Wu, X.; Ding, J.-W.; Lv, Z.-S.; Feng, L.-S. Recent developments of coumarin-containing derivatives and their anti-tubercular activity. Eur. J. Med. Chem. 2017, 136, 122–130. [Google Scholar] [CrossRef]
- Olaharski, A.J.; Rine, J.; Marshall, B.L.; Babiarz, J.; Zhang, L.; Verdin, E.; Smith, M.T. The flavoring agent dihydrocoumarin reverses epigenetic silencing and inhibits sirtuin deacetylases. PLoS Genet. 2005, 1, e77. [Google Scholar] [CrossRef]
- Mohamed, T.K.; Batran, R.Z.; Elseginy, S.A.; Ali, M.M.; Mahmoud, A.E. Synthesis, anticancer effect and molecular modeling of new thiazolylpyrazolyl coumarin derivatives targeting VEGFR-2 kinase and inducing cell cycle arrest and apoptosis. Bioorg. Chem. 2019, 85, 253–273. [Google Scholar] [CrossRef]
- Wu, X.-Q.; Huang, C.; Jia, Y.-M.; Song, B.-A.; Li, J.; Liu, X.-H. Novel coumarin-dihydropyrazole thio-ethanone derivatives: Design, synthesis and anticancer activity. Eur. J. Med. Chem. 2014, 74, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Tang, J.; Gu, Z.; Sun, L.; Wei, H.; Wang, Y.; Yang, S.; Chi, X.; Xu, L. Crystal structure, in vitro cytotoxicity, DNA binding and DFT calculations of new copper (II) complexes with coumarin-amide ligand. J. Inorg. Biochem. 2023, 238, 112030. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C. Ruta angustifolia Pers. (Narrow-Leaved Fringed Rue): Pharmacological Properties and Phytochemical Profile. Plants 2023, 12, 827. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-C.; Huang, C.-P.; Lin, C.-P.; Yang, K.-C.; Lei, Y.-J.; Wang, H.-P.; Kuo, Y.-H.; Chen, Y.-J. Naturally occurring bicoumarin compound daphnoretin inhibits growth and induces megakaryocytic differentiation in human chronic myeloid leukemia cells. Cells 2022, 11, 3252. [Google Scholar] [CrossRef]
- Razavi, S.M. Plants coumarins as allelopathic agents. Int. J. Org. Chem. 2011, 5, 86–90. Available online: https://scialert.net/abstract/?doi=ijbc.2011.86.90 (accessed on 2 January 2023). [CrossRef]
- Sondhia, S.; Duke, S.O.; Green, S.; Gemejiyeva, N.G.; Mamonov, L.K.; Cantrell, C.L. Phytotoxic furanocoumarins from the shoots of Semenovia transiliensis. Nat. Prod. Commun. 2012, 7, 1327–1330. [Google Scholar] [CrossRef]
- Song, P.P.; Zhao, J.; Liu, Z.-L.; Duan, Y.B.; Hu, Y.-P.; Zhao, C.-Q.; Wu, M.; Wei, M.; Wang, N.-H.; Lv, Y.; et al. Evaluation of antifungal activities and structure–activity relationships of coumarin derivatives. Pest Manag. Sci. 2017, 73, 94–101. [Google Scholar] [CrossRef]
- De Andrade Gonçalves, P.; dos Santos Junior, M.C.; do Sacramento Sousa, C.; Góes-Neto, A.; Luz, E.D.M.N.; Damaceno, V.O.; Niella, A.R.R.; Filho, J.M.B.; de Assis, S. A Study of sodium 3-hydroxycoumarin as inhibitors in vitro, in vivo and in silico of Moniliophthora perniciosa fungus. Eur. J. Plant Pathol. 2019, 153, 15–27. [Google Scholar] [CrossRef]
- Wei, Y.; Peng, W.; Wanf, D.; Hao, S.-H.; Li, W.-W.; Ding, F. Design, synthesis, antifungal activity, and 3D-QSAR of coumarin derivatives. J. Pestic. Sci. 2018, 43, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Rastija, V.; Vrandečić, K.; Ćosić, J.; Majić, I.; Kanižai Šarić, G.; Agić, D.; Karnaš, M.; Lončarić, M.; Molnar, M. Biological activities related to plant protection and environmental effects of coumarin derivatives: QSAR and molecular docking studies. Int. J. Mol. Sci. 2021, 22, 7283. [Google Scholar] [CrossRef] [PubMed]
- Rastija, V.; Vrandečić, K.; Ćosić, J.; Šarić, G.K.; Majić, I.; Agić, D.; Šubarić, D.; Karnaš, M.; Bešlo, D.; Komar, M.; et al. Effects of coumarinyl Schiff bases against phytopathogenic fungi, the soil-beneficial bacteria and entomopathogenic nematodes: Deeper insight into the mechanism of action. Molecules 2022, 27, 2196. [Google Scholar] [CrossRef]
- Chen, J.; Yu, Y.; Li, S.; Ding, W. Resveratrol and coumarin: Novel agricultural antibacterial agent against Ralstonia solanacearum in vitro and in vivo. Molecules 2016, 21, 1501. [Google Scholar] [CrossRef]
- Rehman, S.; Ikram, M.; Baker, R.J.; Zubair, M.; Azad, E.; Min, S.; Riaz, K.; Mok, K.H.; Rehman, S.-U. Synthesis, characterization, in vitro antimicrobial, and U2OS tumoricidal activities of different coumarin derivatives. Chem. Cent. J. 2013, 7, 68. Available online: http://hdl.handle.net/2262/69349 (accessed on 15 February 2023). [CrossRef]
- Dekić, B.R.; Radulović, N.S.; Dekić, V.S.; Vukićević, R.D.; Palić, R.M. Synthesis and antimicrobial activity of new 4-heteroarylamino coumarin derivatives containing nitrogen and sulfur as heteroatoms. Molecules 2010, 15, 2246–2256. [Google Scholar] [CrossRef]
- Cui, H.; Jin, H.; Liu, Q.; Yan, Z.; Ding, L.; Qin, B. Nematicidal metabolites from roots of Stellera chamaejasme against Bursaphelenchus xylophilus and Bursaphelenchus mucronatus. Pest Manag. Sci. 2014, 70, 827–835. [Google Scholar] [CrossRef]
- Pan, L.; Li, X.-Z.; Sun, S.-A.; Guo, H.-R.; Qin, B. Design and synthesis of novel coumarin analogs and their nematicidal activity against five phytonematodes. Chin. Chem. Lett. 2016, 27, 375–379. [Google Scholar] [CrossRef]
- Das, S.K. Screening of bioactive compounds for development of new pesticides: A Mini Review. Univers. J. Agric. Res. 2016, 4, 15–20. [Google Scholar] [CrossRef]
- Cao, X.; Xu, S.; Li, X.; Shen, X.; Zhang, Q.; Li, J.; Chen, C. N-Nitrourea derivatives as novel potential fungicides against Rhizoctonia solani: Synthesis, antifungal activities, and 3D-QSAR. Chem. Biol. Drug Des. 2012, 80, 81–89. [Google Scholar] [CrossRef]
- Du, H.; Wang, J.; Hu, Z.; Yao, X.; Zhang, X. Prediction of fungicidal activities of rice blast disease based on least-squares support vector machines and project pursuit pegression. J. Agric. Food Chem. 2008, 56, 10785–10792. [Google Scholar] [CrossRef]
- Lakshman, B.; Gupta, R.L.; Prasad, D. Quantitative structure activity relationships for the nematicidal activity of 4-amino-5-substituted aryl-3-mercapto-(4H)-1,2,4-triazoles. Indian J. Chem. 2010, 49B, 1657–1661. [Google Scholar]
- Kar, S.; Roy, K.; Leszczynski, J. On Applications of QSARs in Food and Agricultural Sciences: History and Critical Review of Recent Developments. In Advances in QSAR Modeling, Challenges and Advances in Computational Chemistry and Physics; Roy, K., Ed.; Springer International Publishing AG: Cham, Switzerland, 2017; Volume 24, pp. 203–300. [Google Scholar] [CrossRef]
- Devillers, J.; Pham-Delègue, M.H.; Decourtye, A.; Budzinski, H.; Cluzeau, S.; Maurin, G. Structure-toxicity modeling of pesticides to honey bees. SAR QSAR Environ. Res. 2002, 13, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Roy, K.; Kar, S.; Das, R.N. Statistical Methods in QSAR/QSPR. In A Primer on QSAR/QSPR Modeling: Fundamental Concepts (SpringerBriefs in Molecular Science); Roy, K., Kar, S., Das, R.N., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 37–59. [Google Scholar] [CrossRef]
- Gramatica, P. Principles of QSAR Modeling: Comments and suggestions from personal experience. Int. J. Quant. Struct. Prop. Relatsh. 2020, 5, 61–97. [Google Scholar] [CrossRef]
- Liu, P.; Long, W. Current mathematical methods used in QSAR/QSPR studies. Int. J. Mol. Sci. 2009, 10, 1978–1998. [Google Scholar] [CrossRef]
- Verma, J.; Khedkar, V.M.; Coutinho, E.C. 3D-QSAR in drug design-a review. Curr. Top. Med. Chem. 2010, 10, 95–115. [Google Scholar] [CrossRef]
- Cramer, R.D. Topomer CoMFA: A design methodology for rapid lead optimization. J. Med. Chem. 2003, 46, 374–388. [Google Scholar] [CrossRef]
- Klebe, G.; Abraham, U.; Mietzner, T. Molecular similarity indices in a comparative analysis (CoMSIA) of drug molecules to correlate and predict their biological activity. J. Med. Chem. 1994, 37, 4130–4146. [Google Scholar] [CrossRef]
- Villaverde, J.J.; Sevilla-Morán, B.; López-Goti, C.; Alonso-Prados, J.L.; Sandín-España, P. QSAR/QSPR models based on quantum chemistry for risk assessment of pesticides according to current European legislation. SAR QSAR Environ. Res. 2020, 31, 49–72. [Google Scholar] [CrossRef] [PubMed]
- OECD. Guidance Document on the Validation of (Quantitative) Structure-Activity Relationship [(Q)SAR] Models: OECD Series on Testing and Assessment; OECD Publishing: Paris, France, 2014; Volume 69. [Google Scholar] [CrossRef]
- ECHA-11-R-004.2-EN. The Use of Alternatives to Testing on Animals for the REACH Regulation 2011; European Chemicals Agency: Helsinki, Finland, 2011; Available online: https://echa.europa.eu/documents/10162/13639/alternatives_test_animals_2011_en.pdf/9b0f7e93-4d61-401d-ba2c-80b3b9faaf66 (accessed on 15 January 2023).
- Jitonnom, J. Computer-aided pesticide design: A short review. In Short Views on Insect Biochemistry and Molecular Biology; Section VIII: Insect Bioinformatics; Chandrasekar, R., Tyagi, B.K., Gui, Z., Reeck, G.R., Eds.; International Book Mission Academic Publisher: Tamilnadu, India, 2014; Volume 2, pp. 685–707. [Google Scholar]
- Meng, X.Y.; Zhang, H.-X.; Mezei, M.; Cui, M. Molecular Docking: A powerful approach for structure-based drug discovery. Curr. Comput. Aided Drug Des. 2011, 7, 146–157. [Google Scholar] [CrossRef]
- Hospital, A.; Goñi, J.R.; Orozco, M.; Gelpi, J.L. Molecular dynamics simulations: Advances and applications. Adv. Appl. Bioinforma. Chem. 2015, 8, 37–47. [Google Scholar] [CrossRef]
- Stringlis, I.A.; de Jonge, R.; Pieterse, C.M.J. The age of coumarins in plant–microbe interactions. Plant Cell Physiol. 2019, 60, 1405–1419. [Google Scholar] [CrossRef] [PubMed]
- Srinivasa, C.; Mellappa, G.; Patil, S.M.; Ramu, R.; Shreevatsa, B.; Dharmashekar, C.; Kollur, S.P.; Syed, A.; Shivamallu, C. Plants and endophytes—A partnership for the coumarin production through the microbial systems. Mycology 2022, 13, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Perkowska, I.; Potrykus, M.; Siwinska, J.; Siudem, D.; Lojkowska, E.; Ihnatowicz, A. Interplay between coumarin accumulation, iron deficiency and plant resistance to Dickeya spp. Int. J. Mol. Sci. 2021, 22, 6449. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, J.; Li, Y.; He, Y.; Yang, Y.; Liu, D.; Wu, C. Effects of coumarin on rhizosphere microbiome and metabolome of Lolium multiflorum. Plants 2023, 12, 1096. [Google Scholar] [CrossRef] [PubMed]
- Harbort, C.J.; Hashimoto, M.H.; Inoue, H.; Niu, Y.; Guan, R.; Rombolá, A.D.; Kopriva, S.; Voges, M.J.; Sattely, E.S.; Garrido-Oter, R.; et al. Root-secreted coumarins and the microbiota interact to improve iron nutrition in Arabidopsis. Cell Host Microbe 2020, 28, 825–837. [Google Scholar] [CrossRef]
- Bye, A.; King, H.K. The biosynthesis of 4-hydroxycoumarin and dicoumarol by Aspergillus fumigatus Fresenius. Biochem. J. 1970, 117, 237–245. [Google Scholar] [CrossRef]
- Aguirre-Pranzoni, C.; Orden, A.A.; Bisogno, F.R.; Ardanaz, C.E.; Tonn, C.E.; Kurina-Sanz, M. Coumarin metabolic routes in Aspergillus spp. Fungal Biol. 2011, 115, 245–252. [Google Scholar] [CrossRef]
- Hoult, J.; Paya, M. Pharmacological and biochemical actions of simple coumarins: Natural products with therapeutic potential. Gen. Pharmac. 1996, 27, 713–722. [Google Scholar] [CrossRef]
- Matos, M.J.; Santana, L.; Uriarte, E.; Abreu, O.A.; Molina, E.; Yordi, E.G. Coumarins—An important class of phytochemicals. In Phytochemicals: Isolation, Characterisation and Role in Human Health; Rao, A.V., Rao, L.G., Eds.; InTech: London, UK, 2015. [Google Scholar] [CrossRef]
- Shimizu, B.-I. 2-Oxoglutarate-dependent dioxygenases in the biosynthesis of simple coumarins. Front. Plant Sci. 2014, 5, 549. [Google Scholar] [CrossRef]
- Wu, F.; Duan, Z.; Xu, P.; Yan, Q.; Meng, M.; Cao, M.; Jones, C.S.; Zong, X.; Zhou, P.; Wang, Y.; et al. Genome and systems biology of Melilotus albus provides insights into coumarins biosynthesis. Plant Biotechnol. J. 2022, 20, 592–609. [Google Scholar] [CrossRef] [PubMed]
- Vismans, G.; van Bentum, S.; Spooren, J.; Song, Y.; Goossens, P.; Valls, J.; Snoek, B.L.; Thiombiano, B.; Schilder, M.; Dong, L.; et al. Coumarin biosynthesis genes are required after foliar pathogen infection for the creation of a microbial soil-borne legacy that primes plants for SA-dependent defenses. Sci. Rep. 2022, 12, 22473. [Google Scholar] [CrossRef]
- Stringlis, I.A.; Yu, K.; Feussner, K.; de Jonge, R.; Van Bentum, S.; Van Verk, M.C.; Berendsen, R.L.; Bakker, P.A.; Feussner, I.; Pieterse, C.M. MYB72-dependent coumarin exudation shapes root microbiome assembly to promote plant health. Proc. Natl. Acad. Sci. USA 2018, 115, E5213–E5222. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Li, P.; Tang, Y.; Fawcett, J.P.; Gu, J. Quantitation of Armillarisin A in human plasma by liquid chromatography–electrospray tandem mass spectrometry. J. Pharm. Biomed. Anal. 2007, 43, 1860–1863. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.H.; Jaki, B.U.; Klein, L.L.; Lankin, D.C.; McAlpine, J.B.; Napolitano, J.G.; Fryling, N.A.; Franzblau, S.G.; Cho, S.H.; Stamets, P.E.; et al. Cholinated coumarins from the polypore mushroom Fomitopsis officialis and their activity against Mycobacterium tuberculosis. J. Nat. Prod. 2013, 76, 1916–1922. [Google Scholar] [CrossRef]
- Robe, K.; Izquierdo, E.; Vignols, F.; Rouached, H.; Dubos, C. The coumarins: Secondary metabolites playing a primary role in plant nutrition and health. Trends Plant Sci. 2021, 26, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Kai, K.; Shimizu, B.; Mizutani, M.; Watanabe, K.; Sakata, K. Accumulation of coumarins in Arabidopsis thaliana. Phytochemistry 2006, 67, 379–386. [Google Scholar] [CrossRef]
- Gnonlonfin, G.J.B.; Sanni, A.; Brimer, L. Review Scopoletin—A coumarin phytoalexin with medicinal properties. Crit. Rev. Plant Sci. 2012, 31, 47–56. [Google Scholar] [CrossRef]
- Beyer, S.F.; Beesley, A.; Rohmann, P.F.W.; Schultheiss, H.; Conrath, U.; Langenbach, J.G. The Arabidopsis non-host defence-associated coumarin scopoletin protects soybean from Asian soybean rust. Plant J. 2019, 99, 397–413. [Google Scholar] [CrossRef]
- Chong, J.; Baltz, R.; Schmitt, C.; Beffa, R.; Fritig, B.; Saindrenan, P. Downregulation of a pathogen-responsive tobacco UDPGlc: Phenylpropanoid glucosyltransferase reduces scopoletin glucoside accumulation, enhances oxidative stress, and weakens virus resistance. Plant Cell 2002, 14, 1093–1107. [Google Scholar] [CrossRef]
- Deng, Y.; Nicholson, R.A. Antifungal properties of surangin B, a coumarin from Mammea longifolia. Planta Med. 2005, 71, 364–365. [Google Scholar] [CrossRef] [PubMed]
- Mazimba, O. Umbelliferone: Sources, chemistry and bioactivities review. Bull. Fac. Pharm. Cairo Univ. 2017, 55, 223–232. [Google Scholar] [CrossRef]
- Yang, L.; Wu, L.; Yao, W.; Zhao, S.; Wang, J.; Li, S.; Ding, W. Hydroxycoumarins: New, effective plant-derived compounds reduce Ralstonia pseudosolanacearum populations and control tobacco bacterial wilt. Microbiol. Res. 2018, 215, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Ding, W.; Xu, Y.; Wu, D.; Li, S.; Chen, J.; Guo, B. New insights into the antibacterial activity of hydroxycoumarins against Ralstonia solanacearum. Molecules 2016, 21, 468. [Google Scholar] [CrossRef]
- Yang, L.; Li, S.; Qin, X.; Jiang, G.; Chen, J.; Li, B.; Yao, X.; Liang, P.; Zhang, Y.; Ding, W. Exposure to umbelliferone reduces Ralstonia solanacearum biofilm formation, transcription of type III secretion system regulators and effectors and virulence on tobacco. Front. Microbiol. 2017, 8, 1234. [Google Scholar] [CrossRef]
- Pastirova, A.; Repcak, M.; Eliasova, A. Salicylic acid induces changes of coumarin metabolites in Matricaria chamomilla L. Plant Sci. 2004, 167, 819–824. [Google Scholar] [CrossRef]
- Han, S.; Yang, L.; Wang, Y.; Ran, Y.; Li, S.; Ding, W. Preliminary studies on the antibacterial mechanism of a new plant-derived compound, 7-methoxycoumarin, against Ralstonia solanacearum. Front. Microbiol. 2021, 12, 697911. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Y.; He, X.; Xiao, Q.; Han, S.; Jia, Z.; Li, S.; Ding, W. Discovery of a novel plant-derived agent against Ralstonia solanacearum by targeting the bacterial division protein FtsZ. Pestic. Biochem. Physiol. 2021, 177, 104892. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Xue, M.; Gu, G.; Wang, W.; Li, D.; Lai, D.; Zhou, L. Synthesis and antibacterial activity of novel chalcone derivatives bearing a coumarin moiety. Lophiostomin A–D: New 3,4-dihydroisocoumarin derivatives from the endophytic fungus Lophiostoma sp. Sigrf10. RSC Adv. 2020, 10, 6985. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.M.; Roseland, C.R. Induction of the coumarins scopoletin and ayapin in sunflower by insect–feeding stress and effects of coumarins on the feeding of sunflower beetle (Coleoptera: Chrysomelidae). Environ. Entomol. 1991, 20, 1166–1172. [Google Scholar] [CrossRef]
- Pavela, R.; Maggi, F.; Benelli, G. Coumarin (2H-1-benzopyran-2-one): A novel and eco-friendly aphicide. Nat. Prod. Res. 2019, 35, 1566–1571. [Google Scholar] [CrossRef]
- Jebir, R.M.; Mustafa, Y.F. Novel coumarins isolated from the seeds of Citrullus lanatus as potential antimicrobial agents. Eurasian Chem. Commun. 2022, 4, 692–708. [Google Scholar] [CrossRef]
- Bruni, R.; Barreca, D.; Protti, M.; Brighenti, V.; Righetti, L.; Anceschi, L.; Mercolini, L.; Benvenuti, S.; Gattuso, G.; Pellati, F. Botanical sources, chemistry, analysis, and biological activity of furanocoumarins of pharmaceutical interest. Molecules 2019, 24, 2163. [Google Scholar] [CrossRef] [PubMed]
- Hadaček, F.; Müller, C.; Werner, A.; Greger, H.; Proksch, P. Analysis, isolation and insecticidal activity of linear furanocoumarins and other coumarin derivatives from Peucedanum (Apiaceae: Apioideae). J. Chem. Ecol. 1994, 20, 2035–2054. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-B.; Li, G.-H.; Li, L.; Zheng, L.-J.; Huang, R.; Zhang, K.-Q. Nematicidal coumarins from Heracleum candicans Wall. Nat. Prod. Res. 2008, 22, 666–671. [Google Scholar] [CrossRef]
- Guo, Q.; Du, G.; He, H.; Xu, H.; Guo, D.; Li, R. Two nematicidal furocoumarins from Ficus carica L. leaves and their physiological effects on pine wood nematode (Bursaphelenchus xylophilus). Nat. Prod. Res. 2016, 30, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Lai, D.; Liu, Q.Z.; Zhou, L.; Liu, Z.L. Identification of nematicidal constituents of Notopterygium incisum rhizomes against Bursaphelenchus xylophilus and Meloidogyne incognita. Molecules 2016, 21, 1276. [Google Scholar] [CrossRef]
- Caboni, P.; Saba, M.; Oplos, C.; Aissani, N.; Maxia, A.; Menkissoglu-Spiroudi, U.; Casua, L.; Ntallia, N. Nematicidal activity of furanocoumarins from parsley against Meloidogyne spp. Pest Manag. Sci. 2015, 71, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.-Q.; Du, G.-C.; Li, Y.-X.; Liang, C.-Y.; Wang, C.; Zhang, Y.-N.; Li, R.-G. Nematotoxic coumarins from Angelica pubescens Maxim. f. biserrata Shan et Yuan roots and their physiological effects on Bursaphelenchus xylophilus. J. Nematol. 2018, 50, 1–10. [Google Scholar] [CrossRef]
- Dugrand-Judek, A.; Olry, A.; Hehn, A.; Costantino, G.; Ollitrault, P.; Froelicher, Y.; Bourgaud, F. The distribution of coumarins and furanocoumarins in Citrus species closely matches citrus phylogeny and reflects the organization of biosynthetic pathways. PLoS ONE 2015, 10, e0142757. [Google Scholar] [CrossRef]
- Ramírez-Pelayo, C.; Martínez-Quiñones, J.; Gil, J.; Durango, D. Coumarins from the peel of citrus grown in Colombia: Composition, elicitation and antifungal activity. Heliyon 2019, 5, e01937. [Google Scholar] [CrossRef]
- Appendino, G.; Bianchi, F.; Bader, A.; Campagnuolo, C.; Fattorusso, E.; Taglialatela-Scafati, O.; Blanco-Molina, M.; Macho, A.; Fiebich, B.L.; Bremner, P.; et al. Coumarins from Opopanax chironium. New dihydrofuranocoumarins and differential induction of apoptosis by imperatorin and heraclenin. J. Nat. Prod. 2004, 67, 532–536. [Google Scholar] [CrossRef]
- Ghasemi, S.; Habibi, Z. A new dihydrofuranocoumarin from Opopanax hispidus (Friv.) Griseb. Nat. Prod. Res. 2014, 28, 1808–1812. [Google Scholar] [CrossRef]
- Alami, I.; Clerivet, A.; Naji, M.; Munster, M.V.; Macheix, J.J. Elicitation of Platanus_acerifolia cell-suspension cultures induces the synthesis of xanthoarnol, a dihydrofuranocoumarin phytoalexin. Phytochemistry 1999, 51, 733–736. [Google Scholar] [CrossRef]
- Taechowisan, T.; Lu, C.; Shen, Y.; Lumyong, S. Secondary metabolites from endophytic Streptomyces aureofaciens CMUAc130 and their antifungal activity. Microbiology 2005, 151, 1691–1695. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Wang, D.X.; Yang, Y.; Liu, P.; Chen, R.Y.; Yu, D.Q. A novel 3-arylcoumarin and three new 2-arylbenzofurans from Mucuna birdwoodiana. Chem. Pharm. Bull. 2010, 58, 254–256. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Wang, P.; Yuan, W.; Li, S. Flavonoids and 3-arylcoumarin from Pterocarpus soyauxii. Planta Med. 2013, 79, 487–491. [Google Scholar] [CrossRef]
- Matos, M.J.; Uriarte, E.; Santana, L. 3-Phenylcoumarins as a privileged scaffold in medicinal chemistry: The Landmarks of the past decade. Molecules 2021, 26, 6755. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.T.; To, D.C.; Tran, T.T.; Tran, M.H.; Nguyen, P.H. PTP1B and-glucosidase inhibitors from Selaginella rolandi-principis and their glucose uptake stimulation. J. Nat. Med. 2021, 75, 186–193. [Google Scholar] [CrossRef]
- Khandy, M.T.; Sofronova, A.K.; Gorpenchenko, T.Y.; Chirikova, N.K. Plant pyranocoumarins: Description, biosynthesis, application. Plants 2022, 11, 3135. [Google Scholar] [CrossRef]
- Anaya, A.L.; Macías-Rubalcava, M.; Cruz-Ortega, R.; García-Santana, C.; Sánchez-Monterrubio, P.N.; Hernández-Bautista, B.E.; Mata, R. Allelochemicals from Stauranthus perforatus, a Rutaceous tree of the Yucatan Peninsula, Mexico. Phytochemistry 2005, 66, 487–494. [Google Scholar] [CrossRef]
- Rosselli, S.; Maggio, A.M.; Faraone, N.; Spadaro, V.; Morris-Natschke, S.L.; Bastow, K.F.; Lee, K.-H.; Bruno, M. The cytotoxic properties of natural coumarins isolated from roots of Ferulago campestris (Apiaceae) and of synthetic ester derivatives of aegelinol. Nat. Prod. Commun. 2009, 4, 1701–1706. [Google Scholar] [CrossRef]
- Mukandiwa, L.; Ahmed, A.; Eloff, J.N.; Naidoo, V. Isolation of seselin from Clausena anisata (Rutaceae) leaves and its effects on the feeding and development of Lucilia cuprina larvae may explain its use in ethnoveterinary medicine. J. Ethnopharmacol. 2013, 2, 886–891. [Google Scholar] [CrossRef] [PubMed]
- Dondon, R.; Bourgeois, P.; Fery-Forgues, S. A new bicoumarin from the leaves and stems of Triphasia trifolia. Fitoterapia 2006, 77, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Sofian, F.F.; Subarnas, A.; Koseki, T.; Shiono, Y. Structure elucidation of a new bicoumarin derivative from the leaves of Dysoxylum parasiticum (Osbeck) Kosterm. Magn. Reson. Chem. 2022, 60, 857–863. [Google Scholar] [CrossRef]
- Xiao, Y.-Q.; Liu, X.-H.; Taniguchi, M.; Baba, K. Bicoumarins from Pleurosperum rivulorum. Phytochemistry 1997, 45, 1275–1277. [Google Scholar] [CrossRef]
- Timson, D.J. Dicoumarol: A drug which hits at least two very different targets in vitamin K metabolism. Curr. Drug Targets 2017, 18, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Teng, P.; Zhang, Y.-L.; Xu, Z.-J.; Zhang, M.-Z.; Zhang, W.-H. Design, synthesis and antifungal activity evaluation of coumarin-3-carboxamide derivatives. Fitoterapia 2018, 127, 387–395. [Google Scholar] [CrossRef]
- Geweely, N.S. Novel inhibition of some pathogenic fungal and bacterial species by new synthetic phytochemical coumarin derivatives. Ann. Microbiol. 2009, 59, 359–368. [Google Scholar] [CrossRef]
- Montagner, C.; de Souza, S.M.; Groposo, C.; Monache, F.D.; Smânia, E.F.A.; Smânia Jr., A. Antifungal activity of coumarins. Z. Nat. C 2008, 63, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Kovač, T.; Kovač, M.; Strelec, I.; Nevistić, A.; Molnar, M. Antifungal and antiaflatoxigenic activities of coumarinyl thiosemicarbazides against Aspergillus flavus NRRL 3251. Arh. Hig. Rada Toksikol. 2017, 68, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Lonačarić, M.; Sušjenka, M.; Molnar, M. An extensive study of coumarin synthesis via Knoevenagel condensation in choline chloride based deep eutectic solvents. Curr. Org. Synth. 2020, 17, 98–108. [Google Scholar] [CrossRef]
- Medimagh-Saidana, S.; Romdhane, A.; Daami-Remadi, M.; Jabnoun-Khiareddine, H.; Touboul, D.; Jannet, H.B.; Hamza, M.A. Synthesis and antimicrobial activity of novel coumarin derivatives from 4-methylumbelliferone. Med. Chem. Res. 2015, 24, 3247–3257. [Google Scholar] [CrossRef]
- Uroos, M.; Javaid, A.; Bashir, A.; Tariq, J.; Khan, I.H.; Naz, S.; Fatima, S.; Sultan, M. Green synthesis of coumarin derivatives using Brønsted acidic pyridinium based ionic liquid [MBSPy][HSO4] to control an opportunistic human and a devastating plant pathogenic fungus Macrophomina phaseolina. RSC Adv. 2022, 12, 23963. [Google Scholar] [CrossRef] [PubMed]
- Desheesh, M.A.; El-Zemity, S.R.; Kadous, E.A.; Fahmy, M.M.; Tawfeek, E.E. Antimicrobial activities of synthesized 3-acetyl coumarin and benzo-4-methyl coumarin. Alex. Sci. Exch. J. 2017, 38, 515–520. [Google Scholar] [CrossRef]
- Wang, Y.H.; Jiang, S.C.; Chen, Y.T.; Xia, R.-J.; Tang, X.; He, M.; Xue, W. Synthesis and antibacterial activity of novel chalcone derivatives bearing a coumarin moiety. Chem. Pap. 2019, 73, 2493–2500. [Google Scholar] [CrossRef]
- Feng, Y.-M.; Qi, P.-Y.; Xiao, W.-L.; Zhang, T.-H.; Zhou, X.; Liu, L.-W.; Yang, S. Fabrication of isopropanolamine-decorated coumarin derivatives as novel quorum sensing inhibitors to suppress plant bacterial disease. J. Agric. Food Chem. 2022, 70, 6037–6049. [Google Scholar] [CrossRef]
- Desmedt, W.; Mangelinckx, S.; Kyndt, T.; Vanholme, B. A phytochemical perspective on plant defense against nematodes. Front. Plant Sci. 2020, 11, 602079. [Google Scholar] [CrossRef]
- Directive 91/414/EEC of the European Communities Concerning the Placing of Plant Protection Products on the Market. 15 July 1991. Available online: https://www.ecolex.org/details/legislation/council-directive-91414eec-concerning-the-placing-of-plant-protection-products-on (accessed on 15 January 2023).
- Desaeger, J.; Wram, C.; Zasada, I. New reduced-risk agricultural nematicides—Rationale and review. J. Nematol. 2020, 52, e2020–e2091. [Google Scholar] [CrossRef]
- Takaishi, K.; Izumi, M.; Baba, N.; Kawazu, K.; Nakajima, S. Synthesis and biological evaluation of alkoxycoumarins as novel nematicidal constituents. Bioorg. Med. Chem. Lett. 2008, 18, 5614–5617. [Google Scholar] [CrossRef]
- Dawara, L.; Singh, R.V. Synthesis, spectroscopic characterization, antimicrobial, pesticidal and nematicidal activity of some nitrogen-oxygen and nitrogen-sulfur donor coumarins based ligands and their organotin(IV) complexes. Appl. Organometal. Chem. 2011, 25, 643–652. [Google Scholar] [CrossRef]
- Kapoor, P.; Singh, R.V.; Fahmi, N. Coordination chemistry of rare earth metal complexes with coumarin-based imines: Ecofriendly synthesis, characterization, antimicrobial, DNA cleavage, pesticidal, and nematicidal activity evaluations. J. Coord. Chem. 2012, 65, 262–277. [Google Scholar] [CrossRef]
- Luo, J.; Lai, T.; Guo, T.; Chen, F.; Zhang, L.; Ding, W.; Zhang, Y. Synthesis and acaricidal activities of scopoletin phenolic ether derivatives: QSAR, molecular dockings and in silico ADME predictions. Molecules 2018, 23, 995. [Google Scholar] [CrossRef] [PubMed]
- Mauri, A.; Consonni, V.; Pavan, M.; Todeschini, R. DRAGON software: An easy approach to molecular descriptor calculations. MATCH Commun. Math. Comput. Chem. 2006, 56, 237–248. [Google Scholar]
- Bingchuan, Z.; Jinxiang, L.; Ting, L.; Dan, W.; Wei, D.; Yongqiang, Z. Study on acaricidal bioactivity and quantitative structure activity relationship of coumarin compounds against Tetranychus cinnabarinus Bois. (Acari: Tetranychidae). Chin. J. Pestic. Sci. 2016, 18, 37–48. [Google Scholar] [CrossRef]
- Hou, Q.-L.; Luo, J.-X.; Zhang, B.-C.; Jiang, G.-F.; Ding, W.; Zhang, Y.-Q. 3D-QSAR and molecular docking studies on the TcPMCA1-mediated detoxification of scopoletin and coumarin derivatives. Int. J. Mol. Sci. 2017, 18, 1380. [Google Scholar] [CrossRef] [PubMed]
- Tyndall, J.D.A.; Sabherwal, M.; Sagatova, A.A.; Keniya, M.V.; Negroni, J.; Wilson, R.K.; Woods, M.A.; Tietjen, K.; Monk, B.C. Structural and functional elucidation of yeast lanosterol 14α-demethylase in complex with agrochemical antifungals. PLoS ONE 2016, 11, e0167485. [Google Scholar] [CrossRef]
- Lockhart, D.E.A.; Schuettelkopf, A.; Blair, D.E.; van Aalten, D.M.F. Screening-based discovery of Aspergillus fumigatus plant-type chitinase inhibitors. FEBS Lett. 2014, 588, 3282–3290. [Google Scholar] [CrossRef]
- Wu, J.; Tao, Y.; Zhang, M.; Howard, M.H.; Gutteridge, S.; Ding, J. Crystal structures of Saccharomyces cerevisiae N-myristoyltransferase with bound myristoyl-CoA and inhibitors reveal the functional roles of the N-terminal region. J. Biol. Chem. 2007, 282, 22185–22194. [Google Scholar] [CrossRef]
- Sulzenbacher, G.; Schülein, M.; Davies, G.J. Structure of the endoglucanase I from Fusarium oxysporum: Native, cellobiose, and 3,4-epoxybutyl β-D-cellobioside-inhibited forms, at 2.3 Å resolution. Biochemistry 1997, 36, 5902–5911. [Google Scholar] [CrossRef]
- Olivieri, F.; Zanetti, E.; Oliva, C.R.; Covarrubias, A.A.; Casalongué, C.A. Characterization of an extracellular serine protease of Fusarium eumartii and its action on pathogenesis related proteins. Eur. J. Plant Pathol. 2002, 108, 63–72. [Google Scholar] [CrossRef]
- Santen, Y.; Benen, J.A.E.; Schröter, K.-H.; Kalk, K.H.; Armand, S.; Visser, J.; Dijkstra, B.W. 1.68-Å Crystal structure of endopolygalacturonase II from Aspergillus niger and identification of active site residues by site-directed mutagenesis. J. Biol. Chem. 1999, 274, 30474–30480. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Wei, W.; Fu, Y.; Cheng, J.; Xie, J.; Li, G.; Yi, X.; Kang, Z.; Dickman, M.B.; Jiamg, D. A secretory protein of necrotrophic fungus Sclerotinia sclerotiorum that suppresses host resistance. PLoS ONE 2013, 8, e53901. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, J.; Shen, H.; Cheng, J.; Li, Z.; Xu, X. Synthesis, nematicidal activity and docking study of novel chromone derivatives containing substituted pyrazole. Chin. Chem Lett. 2018, 29, 911–914. [Google Scholar] [CrossRef]
- Baruah, P.; Basumatary, G.S.; Yesylevskyy, O.; Aguan, K.; Bez, G.; Mitra, S. Novel coumarin derivatives as potent acetylcholinesterase inhibitors: Insight into efficacy, mode and site of inhibition. J. Biomol. Struct. Dyn. 2019, 37, 1750–1765. [Google Scholar] [CrossRef]
- Collin, F.; Karkare, S.; Maxwell, A. Exploiting bacterial DNA gyrase as a drug target: Current state and perspectives. Appl. Microbiol. Biotechnol. 2011, 92, 479–497. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, H.; Najafi, A.; Eslami, H.; Negahdari, B.; Moghaddam, M.M. Identification of novel bacterial DNA gyrase inhibitors: An in silico study. Res. Pharm. Sci. 2016, 11, 250–258. [Google Scholar]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Hao, G.; Dong, Q.; Yang, G.A. Comparative study on the constitutive properties of marketed pesticides. Mol. Inform. 2011, 30, 614–622. [Google Scholar] [CrossRef]
- Clarke, E.D.; Delaney, J.S. Physical and molecular properties of agrochemicals: An analysis of screen inputs, hits, leads, and products. Chimia 2003, 57, 731–734. [Google Scholar] [CrossRef]
- Avram, S.; Funar-Timofei, S.; Borota, A.; Chennamaneni, S.R.; Manchala, A.K.; Muresan, S. Quantitative estimation of pesticide-likeness for agrochemical discovery. J. Cheminform. 2014, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.; Wu, Z.; Yi, J.; Fu, L.; Yang, Z.; Hsieh, C.; Yin, M.; Zeng, X.; Wu, C.; Lu, A.; et al. ADMETlab 2.0: An integrated online platform for accurate and comprehensive predictions of ADMET properties. Nucl. Acids Res. 2021, 49, W5–W14. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Helepciuc, F.-E.; Todor, A. Evaluating the effectiveness of the EU’s approach to the sustainable use of pesticides. PLoS ONE 2021, 16, 1–18. [Google Scholar] [CrossRef]
- Umetsu, N.; Shirai, Y. Development of novel pesticides in the 21st century. J. Pestic. Sci. 2020, 45, 54–74. [Google Scholar] [CrossRef] [PubMed]
- Majić, I.; Sarajlić, A.; Lakatos, T.; Tóth, T.; Raspudić, E.; Kanižai Šarić, G.; Laznik, Ž. Compatibility of bio-nematicide and plant stimulant of microbial origin with Heterorhabditis bacteriophora. IOBC/WPRS Bull. 2017, 129, 125–129. [Google Scholar]
- Bai, X.; Adams, B.J.; Ciche, T.A.; Clifton, S.; Gaugler, R.; Kim, K.S.; Grewal, P.S. A lover and a fighter: The genome sequence of an entomopathogenic nematode Heterorhabditis bacteriophora. PLoS ONE 2013, 8, e69618. [Google Scholar] [CrossRef]
- Özdemir, E.; İnak, E.; Evlice, E.; Laznik, Z. Compatibility of entomopathogenic nematodes with pesticides registered in vegetable crops under laboratory conditions. J. Plant Dis. Prot. 2020, 127, 529–535. [Google Scholar] [CrossRef]
- Ntalli, N.G.; Caboni, P. Botanical nematicides: A review. J. Agric. Food Chem. 2012, 60, 9929–9940. [Google Scholar] [CrossRef]
- Abenavoli, M.R.; Cacco, G.; Sorgonà, A.; Marabottini, R.; Paolacci, A.R.; Ciaffi, M.; Badiani, M. The inhibitory effects of coumarin on the germination of durum wheat (Triticum turgidum ssp. durum, cv. Simeto) seeds. J. Chem. Ecol. 2006, 32, 489–506. [Google Scholar] [CrossRef]
- Chen, B.X.; Peng, Y.-X.; Gao, J.-D.; Zhang, Q.; Liu, Q.-J.; Fu, H.; Liu, J. Coumarin-induced delay of rice seed germination is mediated by suppression of abscisic acid catabolism and reactive oxygen species production. Front. Plant Sci. 2019, 10, 828. [Google Scholar] [CrossRef] [PubMed]
- Runciman, D.J.; Lee, A.M.; Reed, K.F.; Walsh, J.R. Dicoumarol toxicity in cattle associated with ingestion of silage containing sweet vernal grass (Anthoxanthum odoratum). Aust. Vet. J. 2002, 80, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, H.; Lacasta, D.; Ramos, J.J.; Quintas, H.; Ruiz de Arcaute, M.; Ramo, M.Á.; Villanueva-Saz, S.; Ferrer, L.M. Anaemia in ruminants caused by plant consumption. Animals 2022, 12, 2373. [Google Scholar] [CrossRef] [PubMed]
- Lake, B.G. Coumarin metabolism, toxicity and carcinogenicity: Relevance for human risk assessment. Food Chem. Toxicol. 1999, 37, 423–453. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H.; Jiang, J.M.; Zheng, D.; Chen, Y.Y.; Wan, S.J.; Tan, H.-S.; Tang, L.-M.; Xu, H.-X. Hepatotoxicity induced by psoralen and isopsoralen from Fructus psoraleae: Wistar rats are more vulnerable than ICR mice. Food Chem. Toxicol. 2019, 125, 133–140. [Google Scholar] [CrossRef]
- Shokoohinia, Y.; Bazargan, S.; Miraghaee, S.; Javadirad, E.; Farahani, F.; Hosseinzadeh, L. Safety assessment of osthole isolated from Prangos ferulacea: Acute and subchronic toxicities and modulation of cytochrome P450. Jundishapur J. Nat. Pharm. Prod. 2017, 12, e63764. [Google Scholar] [CrossRef]
- Abraham, K.; Wöhrlin, F.; Lindtner, O.; Heinemeyer, G.; Lampen, A. Toxicology and risk assessment of coumarin: Focus on human data. Mol. Nutr. Food Res. 2010, 54, 228–239. [Google Scholar] [CrossRef]
- Loprinzi, C.L.; Kugler, J.W.; Sloan, J.A.; Rooke, T.W.; Quella, S.K.; Novotny, P.; Mowat, R.B.; Michalak, J.C.; Stella, P.J.; Levitt, R.; et al. Lack of effect of coumarin in women with lymphedema after treatment for breast cancer. N. Engl. J. Med. 1999, 340, 346–350. [Google Scholar] [CrossRef]
- Wagstaff, D.J. Dietary Exposure to Furocoumarins. Regul. Toxicol. Pharmacol. 1991, 14, 261–272. [Google Scholar] [CrossRef]
- Heghes, S.C.; Vostinaru, O.; Mogosan, C.; Miere, D.; Iuga, C.A.; Filip, L. Safety profile of nutraceuticals rich in coumarins: An update. Front. Pharmacol. 2022, 13, 803338. [Google Scholar] [CrossRef]
- Moreira-Filho, J.T.; Braga, R.C.; Lemos, J.M.; Alves, V.M.; Borba, J.V.V.B.; Costa, W.S.; Kleinstreuer, N.; Muratov, E.N.; Andrade, C.H.; Neves, B.J. BeeToxAI: An artificial intelligence-based web app to assess acute toxicity of chemicals to honey bees. Artif. Intell. Life Sci. 2021, 1, 100013. [Google Scholar] [CrossRef]
- Como, F.; Carnesecchi, E.; Volani, S.; Dorne, J.L.; Richardson, J.; Bassan, A.; Pavan, M.; Benfenati, E. Predicting acute contact toxicity of pesticides in honeybees (Apis mellifera) through a k-nearest neighbor model. Chemosphere 2017, 166, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.M. User’s Guide for T.E.S.T. (Version 5.1) (Toxicity Estimation Software Tool): A Program to Estimate Toxicity from Molecular Structure; U.S. Environmental Protection Agency: Washington, DC, USA, 2020. Available online: https://www.epa.gov/chemical-research/toxicity-estimation-software-tool-test (accessed on 15 January 2023).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum/ Famillies | Species | Group of Coumarins | Specific Compounds | Known Biological Activities | Ref. |
|---|---|---|---|---|---|
| Bacteria | |||||
| Streptomyces | Streptomyces roseochromogenes var. oscitans | 3-amino-4,7-dihydroxycoumarins | clorobiocin, novobiocin, coumermycin | antibacterial | [15] |
| Porifera | |||||
| Axinellidae | Axinella cf. corrugate | simple | esculetin-4-carboxylic acid esters | anti-SARS-CoV | [22] |
| Fungi | |||||
| Pleosporaceae | Alternaria alternata | simple | isofraxidin | antibacterial | [61] |
| Trichocomaceae | Aspergillus fumigatus Fresenius | simple, bicoumarins | 4-hydroxycoumarin, dicoumarol | biosynthesis of coumarin | [65,66] |
| Physalacriaceae | Armillariella tabescens | simple | armillarisin A | choleretic | [73] |
| Fomitopsidaceae | Fomitopsis officinalis | simple | 6-chloro-2-oxo-4-phenyl-coumarins | anti-TBC | [74] |
| Lophiostomataceae | Lophiostoma sp. Sigrf10 | 3,4-dihydroisocoumarin | lophiostomin derivatives | antifungal, antibacterial | [88] |
| Plants | |||||
| Citrus | C. maxima, C. medica, C. reticulata, C. micrantha | simple, furanocoumarin | unknown | [99] | |
| Citrus sinensis, C. reticulata, C. aurantifolia | simple, furanocoumarin | limettin, isopimpinellin, psoralen, bergamottin | antifungal | [100] | |
| Cucurbitaceae | Citrullus lanatus | simple | derivates of 5,7-dimethoxycoumarin | antimicrobial | [91] |
| Apiaceae | Ferulago campestris | pyranocoumarin | aegelinol, grandivittin, | cytotoxicity | [111] |
| (or Umbelliferae) | furanocoumarin | bergapten, felamidin, isoimperatorin | antimicrobial, antioxidant | [16] | |
| Notopterygium incisum | dihidrofuranocoumarin | columbianetin | nematicidal | [96] | |
| linear furanocoumarin | isoimperatorin | nematicidal | |||
| Petroselinum crispum | furanocoumarins | xanthotoxin, psoralen, bergapten | nematicidal | [97] | |
| Angelica pubescens Maxim. f. biserrata Shan et Yuan | simple, dihidrofuranocoumarin, | osthole, columbianadin | nematicidal | [98] | |
| furanocoumarin | bergapten, xanthotoxin | nematicidal | [115] | ||
| Pleurospermum rivulorum | bicoumarin | rivulobirins | unknown | ||
| Opopanax hispidus(Friv.) Griseb. | dihydrofuranocoumarin | 3′-isobutyryl-3′-hydroxymarmesin | unknown | [102] | |
| simple, furanocoumarin | officinalin, oreoselon, peucedanin, | unknown | |||
| Peucedanum sp. | simple, furanocoumarin | ostruthin, osthol; isoimperatorin | insecticidal | [93] | |
| dihydropyranocoumarin | xanthalin, peuarenarin | insecticidal | |||
| dihydrofuranocoumarin | athamantin, columbianadin | insecticidal | |||
| Semenovia transiliensis | furanocoumarin | Imperatorin, xanthotoxin | herbicidal | [31] | |
| Heracleum candicans Wall. | uranocoumarin | 8-geranyloxy psolaren, imperatorin, heraclenin | nematicidal | [94] | |
| Fabaceae | Melilotus officinalis | simple | dihydrocoumarin | cytotoxicity | [24] |
| bicoumarin | dicoumarol | anticoagulant | [116] | ||
| Mucuna birdwoodiana | phenylcoumarin | mucodianin A | unknown | [107] | |
| Sphenostylis marginata | phenylcoumarin | sphenostylisin A | anticancer | [68] | |
| Pterocarpus soyauxii | phenylcoumarin | pterosonins | anticancer | [106] | |
| Millettia thonningii | pyranocoumarin, furanocoumarin | robustic acid, thonningine-C | antifungal | [20] | |
| Solanaceae | Nicotiana tabacum | simple | scopolin, scopoletin | antiviral | [79] |
| Lamiaceae | Baikal skullcap | simple | 7.8-dihydroxy-4-methylcumarin | antibacterial | [17] |
| Brassicales | Arabidopsis thaliana | simple | scopoletin | antifungal | [78] |
| Moraceae | Ficus carica | furocoumarin | bergapten, psoralen | nematicidal | [95] |
| Meliaceae | Dysoxylum parasiticum (Osbeck) Kosterm | bicoumarin | bidysoxyletine | unknown | [114] |
| Rutaceae | Triphasia trifolia | simple, furocoumarin | umbelliferone, isopimpinellin, | unknown | [113] |
| Xanthoxylum arnottianum | dihydrofuranocoumarin | xanthoarnol | antifungal | [103] | |
| Staurantus perforatus | pyranocoumarin | xanthyletin | phytotoxic | [110] | |
| Ruta angustifolia | furocoumarin, dihydrofuranocoumarin | chalepensin, chalepin | anticancer, antiviral | [28] | |
| Clausena anisata | pyranocoumarin | seselin | antifeedant | [112] | |
| Thymelaeaceae | Wikstroemia indica (L.) | bicoumarin | daphnoretin | antiviral, antitumor | [29] |
| Calophyllaceae | Mammea longifolia | simple | surangib B | antifungal | [80] |
| Coumarin Derivatives | Biological Activity | Phytopathogenic Organism | Reference |
|---|---|---|---|
| coumarin-3-carboxamides/hydrazides | antifungal | Botrytis cinerea, Colletotrichum capsica, Rhizoctorzia solani, Cucumber anthrax, and Alternaria leaf spot | [117] |
| sodium 3-hydroxycoumarin | antifungal | Moniliophthora perniciosa | [33] |
| copper (II) complexe with coumarins, [L2Cu(OAc)] | antifungal antibacterial | Alternaria alternata Bacillus subtilis | [118] |
| 8-substituted coumarins | antifungal | Botrytis cinerea, Colletotrichum gloeosporioides, Fusarium oxysporum, Valsa mali | [34] |
| 7-substituted-coumarinyl thiosemicarbazides | antifungal | Aspergillus flavus Link | [120] |
| -CN and 6-OH simple coumarins | antifungal | Fusarium oxysporum f. sp. lycopersici, Fusarium culmorum, Macrophomina phaseolina, and Sclerotinia sclerotiourum | [35] |
| coumarinyl Schiff bases | antifungal | Macrophomina phaseolina | [36] |
| 4-methylumbelliferone, coumarin esters | antifungal | Fusarium oxysporum f. sp. lycopersici | [122] |
| 4-(chloromethyl)-7-hydroxycoumarin; 4-(chloromethyl)-7,8-dihydroxycoumarin | antifungal | Macrophomina phaseolina | [123] |
| benzo-4-methyl coumarin | antibacterial | Erwinia amylovora, Ralstonia solanacearum | [124] |
| isopropanolamine coumarin derivative | antibacterial | Xanthomonas oryzae pv. oryzae (Xoo) | [126] |
| 5-ethoxycoumarin | nematicidal | Bursaphelenchus xylophilus | [130] |
| tin(IV) complexes with 3-formyl-4-chlorocoumarin hydrazinecarbothioamide and 3-formyl-4-chlorocoumarin thiosemicarbazone | nematicidal | Meloidogyne incognita, Ditylenchus destructor, Bursaphelenchus mucronatus, B. xylophilus | [41] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rastija, V.; Vrandečić, K.; Ćosić, J.; Kanižai Šarić, G.; Majić, I.; Karnaš, M. Prospects of Computer-Aided Molecular Design of Coumarins as Ecotoxicologically Safe Plant Protection Agents. Appl. Sci. 2023, 13, 6535. https://doi.org/10.3390/app13116535
Rastija V, Vrandečić K, Ćosić J, Kanižai Šarić G, Majić I, Karnaš M. Prospects of Computer-Aided Molecular Design of Coumarins as Ecotoxicologically Safe Plant Protection Agents. Applied Sciences. 2023; 13(11):6535. https://doi.org/10.3390/app13116535
Chicago/Turabian StyleRastija, Vesna, Karolina Vrandečić, Jasenka Ćosić, Gabriella Kanižai Šarić, Ivana Majić, and Maja Karnaš. 2023. "Prospects of Computer-Aided Molecular Design of Coumarins as Ecotoxicologically Safe Plant Protection Agents" Applied Sciences 13, no. 11: 6535. https://doi.org/10.3390/app13116535
APA StyleRastija, V., Vrandečić, K., Ćosić, J., Kanižai Šarić, G., Majić, I., & Karnaš, M. (2023). Prospects of Computer-Aided Molecular Design of Coumarins as Ecotoxicologically Safe Plant Protection Agents. Applied Sciences, 13(11), 6535. https://doi.org/10.3390/app13116535