
Advances in the Characterization of Usnea barbata (L.) Weber ex F.H. Wigg from Călimani Mountains, Romania

 ,
, 
 ,
,  ,
,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lichen Sample Preparation
2.2. Lichen Morphology
2.2.1. Fluorescent Microscopy
2.2.2. Scanning Electron Microscopy
2.3. Color Evaluation
2.4. Elemental Analysis
2.5. FT-IR Analysis
2.6. Total Phenolic Content
2.6.1. Dried Lichen Extracts in Ethanol and Acetone
2.6.2. Folin–Ciocâlteu Method
2.7. Free Radical Scavenging Activity
2.8. Statistical Analysis
3. Results
3.1. Lichen Samples
3.2. Lichen Morphology
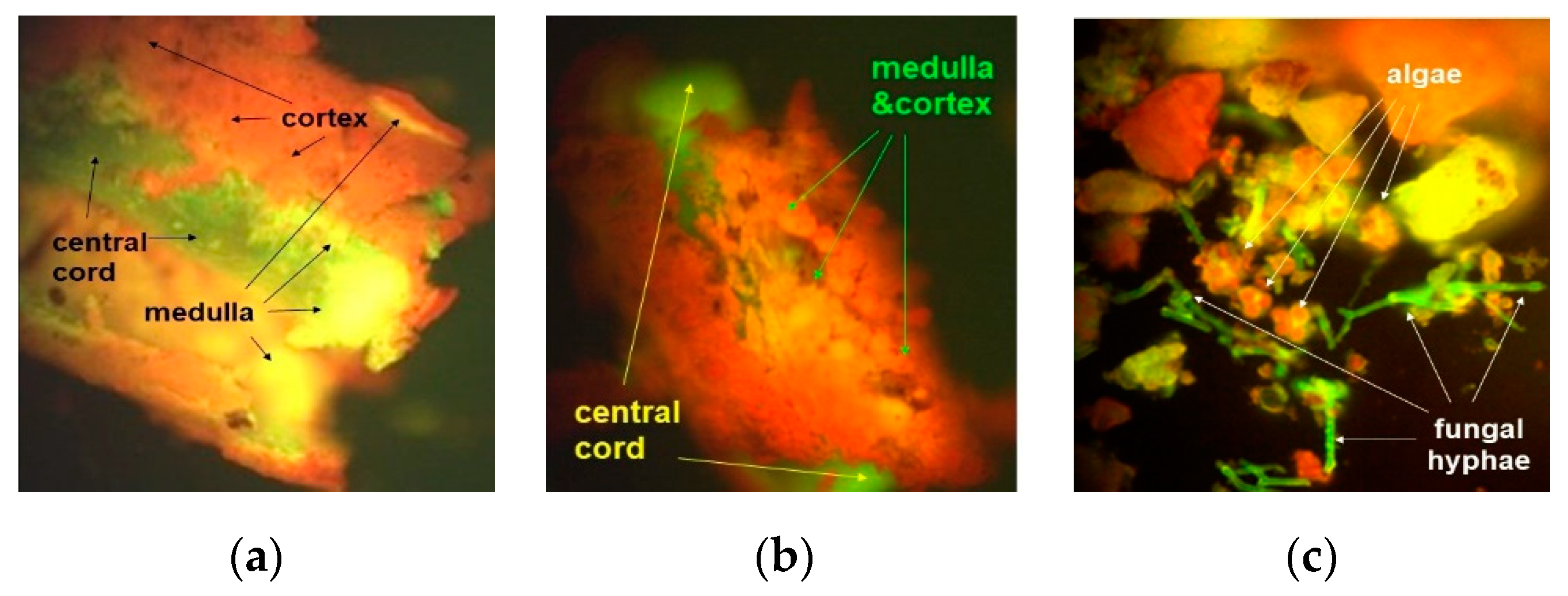
3.2.1. Fluorescence Microscopy
3.2.2. Scanning Electron Microscopy
3.3. Color Evaluation
3.4. Elemental Analysis
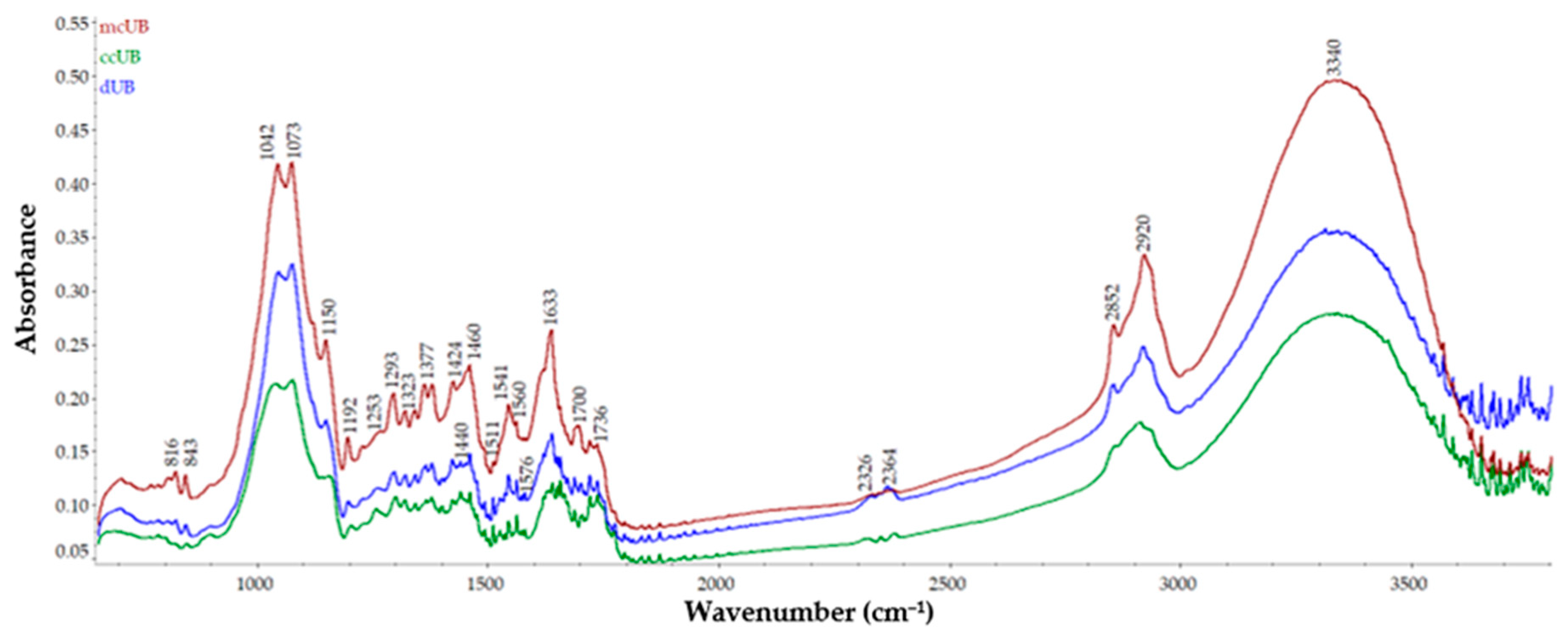
3.5. FT-IR Analysis
3.6. Total Phenolic Content
3.7. Free Radical Scavenging Activity
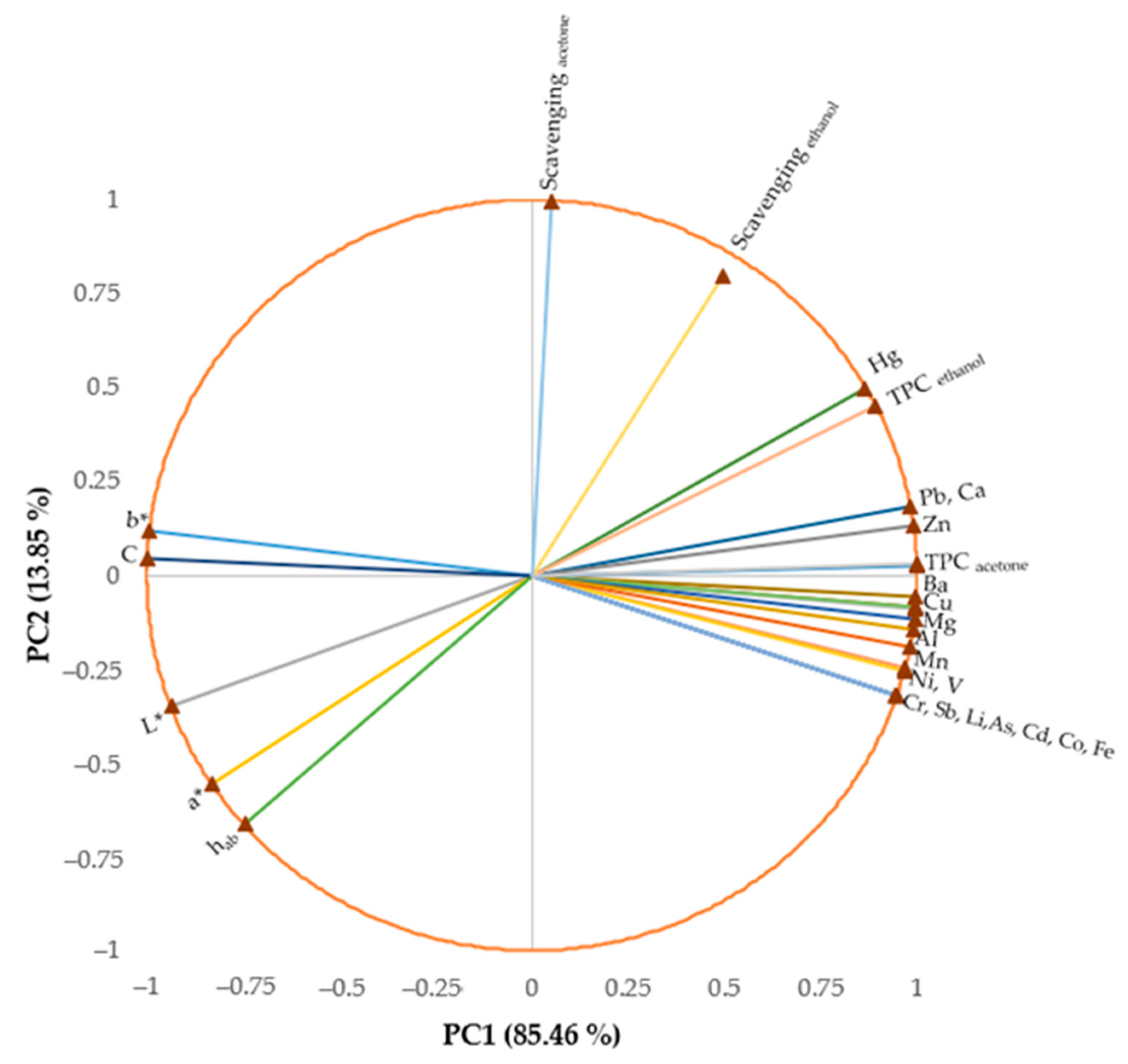
3.8. Relationships between Variables
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grube, M.; Wedin, M. Lichenized Fungi and the Evolution of Symbiotic Organization. Microbiol. Spectr. 2016, 4, funk-0011-2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido-Benavent, I.; Pérez-Ortega, S. Past, present, and future research in bipolar lichen-forming fungi and their photobionts. Am. J. Bot. 2017, 104, 1660–1674. [Google Scholar] [CrossRef] [Green Version]
- Shukla, V.; Joshi, G.P.; Rawat, M.S.M. Lichens as a potential natural source of bioactive compounds: A review. Phytochem. Rev. 2010, 9, 303–314. [Google Scholar] [CrossRef]
- Zambare, V.P.; Christopher, L.P. Biopharmaceutical potential of lichens. Pharm. Biol. 2012, 50, 778–798. [Google Scholar] [CrossRef]
- Prateeksha; Paliya, B.S.; Bajpai, R.; Jadaun, V.; Kumar, J.; Kumar, S.; Upreti, D.K.; Singh, B.N.R.; Nayaka, S.; Joshi, Y.; et al. The genus Usnea: A potent phytomedicine with multifarious ethnobotany, phytochemistry and pharmacology. RSC Adv. 2016, 6, 21672–21696. [Google Scholar] [CrossRef]
- Yang, M.X.; Devkota, S.; Wang, L.S.; Scheidegger, C. Ethnolichenology—the use of lichens in the himalayas and southwestern parts of china. Diversity 2021, 13, 330. [Google Scholar] [CrossRef]
- Huang, Y.P.; Xiang, J.T.; Wang, C.H.; Ren, D.; Johnson, D.; Xu, T. Lichen as a Biomonitor for Vehicular Emission of Metals: A Risk Assessment of Lichen Consumption by the Sichuan Snub-Nosed Monkey (Rhinopithecus roxellana). Ecotoxicol. Environ. Saf. 2019, 180, 679–685. [Google Scholar] [CrossRef]
- Behera, B.C.; Verma, N.; Sonone, A.; Makhija, U. Optimization of culture conditions for lichen Usnea ghattensis G. Awasthi to increase biomass and antioxidant metabolite production. Food Technol. Biotechnol. 2009, 47, 7–12. [Google Scholar]
- Randlane, T.; Tõrra, T.; Saag, A.; Saag, L. Key to European Usnea species. Bibl. Lichenol. 2009, 100, 419–462. [Google Scholar]
- Mesta, A.R.; Rajeswari, N.; Kanivebagilu, V.S. Distribution of bioactive compounds in usneoid lichens from Western Ghats. Plant Arch. 2019, 19, 2163–2168. [Google Scholar]
- Shcherbokova, A.; Koptina, A.; Samiento-Diaz, M.; Ulrich-Merzenich, G. Pharmacological activities of lichens from Russia. Planta Med. 2015, 81, PM_134. [Google Scholar] [CrossRef]
- Ohmura, Y. A synopsis of the lichen genus Usnea (Parmeliaceae, Ascomycota) in Taiwan. Mem. Natl. Museum Nat. Sci. Tokyo 2012, 48, 91–137. [Google Scholar]
- Cansaran, D.; Kahya, D.; Yurdakulol, E.; Atakol, O. Identification and quantitation of usnic acid from the lichen Usnea species of Anatolia and antimicrobial activity. Z. Naturforsch. Sect. C J. Biosci. 2006, 61, 773–776. [Google Scholar] [CrossRef] [PubMed]
- Rafat, A. Ecological and Biological Studies of New Zealand Lichens in the Genus Usnea. Ph.D. Thesis, Lincoln University, Chester County, PA, USA, 2014; pp. 18–153. [Google Scholar]
- Đorðević, S.; Ivanović, J.; Kukić-Marković, J.; Petrović, S.; Žižović, I.; Tadić, V.; Marković, G. HPLC determination of usnic acid content in different extracts of Usnea barbata. Planta Med. 2010, 76, LS1. [Google Scholar] [CrossRef]
- Redzic, S.; Barudanovic, S.; Pilipovic, S. Wild Mushrooms and Lichens used as Human Food for Survival in War Conditions; Podrinje. Hum. Ecol. Rev. 2010, 17, 175–181. [Google Scholar]
- Gómez-Serranillos, M.P.; Fernández-Moriano, C.; González-Burgos, E.; Divakar, P.K.; Crespo, A. Parmeliaceae family: Phytochemistry, pharmacological potential and phylogenetic features. RSC Adv. 2014, 4, 59017–59047. [Google Scholar] [CrossRef]
- Bazarnova, Y.; Politaeva, N.; Lyskova, N. Research for the lichen Usnea barbata metabolites. Z. Naturforsch. Sect. C J. Biosci. 2018, 73, 291–296. [Google Scholar] [CrossRef]
- Salgado, F.; Albornoz, L.; Cortéz, C.; Stashenko, E.; Urrea-Vallejo, K.; Nagles, E.; Galicia-Virviescas, C.; Cornejo, A.; Ardiles, A.; Simirgiotis, M.; et al. Secondary metabolite profiling of species of the genus Usnea by UHPLC-ESI-OT-MS-MS. Molecules 2018, 23, 54. [Google Scholar] [CrossRef] [Green Version]
- Ranković, B. Lichen Secondary Metabolites: Bioactive Properties and Pharmaceutical Potential; Springer: Cham, Switzerland, 2015; pp. 1–29. [Google Scholar]
- Bubach, D.; Dufou, L.; Catán, S.P. Evaluation of dispersal volcanic products of recent events in lichens in environmental gradient, Nahuel Huapi National Park, Argentina. Environ. Monit. Assess. 2014, 186, 4997–5007. [Google Scholar] [CrossRef]
- Sanders, W.B.; Masumoto, H. Lichen algae: The photosynthetic partners in lichen symbioses. Lichenologist 2021, 53, 347–393. [Google Scholar] [CrossRef]
- Lyskova, N.S.; Bazarnova, I.G.; Kruchina-Bogdanov, I.V. Study of the composition and properties of secondary metabolites of the Lichen Usnea barbata. Khimiya Rastit. Syr’ya 2018, 1, 121–127. [Google Scholar]
- Bačkor, M.; Fahselt, D. Lichen photobionts and metal toxicity. Symbiosis 2008, 46, 1–10. [Google Scholar]
- Hájek, J.; Hojdová, A.; Trnková, K.; Váczi, P.; Bednaříková, M.; Barták, M. Responses of thallus anatomy and chlorophyll fluorescence-based photosynthetic characteristics of two antarctic species of genus Usnea to low temperature. Photosynthetica 2021, 59, 95–105. [Google Scholar] [CrossRef]
- Coufalík, P.; Uher, A.; Zvěřina, O.; Komárek, J. Determination of cadmium in lichens by solid sampling graphite furnace atomic absorption spectrometry (SS-GF-AAS). Environ. Monit. Assess. 2020, 192, 222. [Google Scholar] [CrossRef]
- Carreras, H.A.; Wannaz, E.D.; Perez, C.A.; Pignata, M.L. The role of urban air pollutants on the performance of heavy metal accumulation in Usnea amblyoclada. Environ. Res. 2005, 97, 50–57. [Google Scholar] [CrossRef]
- Popovici, V.; Bucur, L.; Calcan, S.I.; Cucolea, E.I.; Costache, T.; Rambu, D.; Schröder, V.; Gîrd, C.E.; Gherghel, D.; Vochita, G.; et al. Elemental Analysis and In Vitro Evaluation of Antibacterial and Antifungal Activities of Usnea barbata (L.) Weber ex F.H. Wigg from Călimani Mountains, Romania. Plants 2022, 11, 32. [Google Scholar] [CrossRef]
- Popovici, V.; Bucur, L.; Gîrd, C.E.; Rambu, D.; Calcan, S.I.; Cucolea, E.I.; Costache, T.; Ungureanu-Iuga, M.; Oroian, M.; Mironeasa, S.; et al. Antioxidant, Cytotoxic, and Rheological Properties of Canola Oil Extract of Usnea barbata ( L.) Weber ex F. H. Wigg from Călimani Mountains, Romania. Plants 2022, 11, 854. [Google Scholar] [CrossRef]
- Pop, A.; Fizeșan, I.; Vlase, L.; Rusu, M.E.; Cherfan, J.; Babota, M.; Gheldiu, A.M.; Tomuta, I.; Popa, D.S. Enhanced recovery of phenolic and tocopherolic compounds from walnut (Juglans regia L.) male flowers based on process optimization of ultrasonic assisted-extraction: Phytochemical profile and biological activities. Antioxidants 2021, 10, 607. [Google Scholar] [CrossRef]
- Farmacopeea Rom. Ed. X, 1993. Available online: https://ro.scribd.com/doc/215542717/Farmacopeea-Romana-X (accessed on 2 February 2022).
- Popovici, V.; Bucur, L.; Popescu, A.; Schröder, V.; Costache, T.; Rambu, D.; Cucolea, I.E.; Gîrd, C.E.; Caraiane, A.; Gherghel, D.; et al. Antioxidant and cytotoxic activities of Usnea barbata (L.) F.H. Wigg. dry extracts in different solvents. Plants 2021, 10, 909. [Google Scholar] [CrossRef]
- Pegg, T.J.; Gladish, D.K.; Baker, R.L. Algae to angiosperms: Autofluorescence for rapid visualization of plant anatomy among diverse taxa. Appl. Plant Sci. 2021, 9, 1–8. [Google Scholar] [CrossRef]
- Pathan, A.K.; Bond, J.; Gaskin, R.E. Sample preparation for SEM of plant surfaces. Mater. Today 2010, 12, 32–43. [Google Scholar] [CrossRef]
- Yuan, J.; Wang, X.; Zhou, H.; Li, Y.; Zhang, J.; Yu, S.; Wang, M.; Hao, M.; Zhao, Q.; Liu, L.; et al. Comparison of Sample Preparation Techniques for Inspection of Leaf Epidermises Using Light Microscopy and Scanning Electronic Microscopy. Front. Plant Sci. 2020, 11, 133. [Google Scholar] [CrossRef]
- Thiberge, S.; Nechushtan, A.; Sprinzak, D.; Gileadi, O.; Behar, V.; Zik, O.; Chowers, Y.; Michaeli, S.; Schlessinger, J.; Moses, E. Scanning electron microscopy of cells and tissues under fully hydrated conditions. Proc. Natl. Acad. Sci. USA 2004, 101, 3346–3351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanovic, J.; Meyer, F.; Misic, D.; Asanin, J.; Jaeger, P.; Zizovic, I.; Eggers, R. Influence of different pre-treatment methods on isolation of extracts with strong antibacterial activity from lichen Usnea barbata using carbon dioxide as a solvent. J. Supercrit. Fluids 2013, 76, 1–9. [Google Scholar] [CrossRef]
- Ingoífsdotir, K. Molecules of Interest Usnic acid. Phytochemistry 2002, 61, 729–736. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Solhaug, K.A. Differences in the Susceptibility to Light Stress Between Epiphytic Lichens of Ancient and Young Boreal Forest Stands. Funct. Ecol. 1996, 10, 344. [Google Scholar] [CrossRef]
- Varol, M.; Tay, T.; Candan, M.; Türk, A.; Koparal, A.T. Evaluation of the sunscreen lichen substances usnic acid and atranorin. Biocell 2015, 39, 25–31. [Google Scholar]
- Zulaini, A.A.M.; Muhammad, N.; Asman, S.; Hashim, N.H.; Jusoh, S.; Abas, A.; Yusof, H.; Din, L. Evaluation of transplanted lichens, Parmotrema tinctorum and Usnea diffracta as bioindicator on heavy metals accumulation in southern Peninsular Malaysia. J. Sustain. Sci. Manag. 2019, 14, 1–13. [Google Scholar]
- Bačkorová, M.; Jendželovský, R.; Kello, M.; Bačkor, M.; Mikeš, J.; Fedoročko, P. Lichen secondary metabolites are responsible for induction of apoptosis in HT-29 and A2780 human cancer cell lines. Toxicol. Vitr. 2012, 26, 462–468. [Google Scholar] [CrossRef]
- Gadea, A.; Fanuel, M.; Le Lamer, A.C.; Boustie, J.; Rogniaux, H.; Charrier, M.; Devehat, F.L. Le Mass spectrometry imaging of specialized metabolites for predicting lichen fitness and snail foraging. Plants 2020, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Talamond, P.; Verdeil, J.L.; Conéjéro, G. Secondary metabolite localization by autofluorescence in living plant cells. Molecules 2015, 20, 5024–5037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Pogam, P.; Legouin, B.; Le Lamer, A.C.; Boustie, J.; Rondeau, D. Analysis of the cyanolichen Lichina pygmaea metabolites using in situ DART-MS: From detection to thermochemistry of mycosporine serinol. J. Mass Spectrom. 2015, 50, 445–462. [Google Scholar] [CrossRef]
- Le Pogam, P.; Legouin, B.; Geairon, A.; Rogniaux, H.; Lohézic-Le Dévéhat, F.; Obermayer, W.; Boustie, J.; Le Lamer, A.C. Spatial mapping of lichen specialized metabolites using LDI-MSI: Chemical ecology issues for Ophioparma ventosa. Sci. Rep. 2016, 6, 37807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komaty, S.; Letertre, M.; Dang, H.D.; Jungnickel, H.; Laux, P.; Luch, A.; Carrié, D.; Merdrignac-Conanec, O.; Bazureau, J.P.; Gauffre, F.; et al. Sample preparation for an optimized extraction of localized metabolites in lichens: Application to Pseudevernia furfuracea. Talanta 2016, 150, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Culicov, O.A.; Yurukova, L. Comparison of element accumulation of different moss and lichen-bags, exposed in the city of Sofia (Bulgaria). J. Atmos. Chem. 2006, 55, 1–12. [Google Scholar] [CrossRef]
- Jayasekera, R.; Rossbach, M. Background levels of heavy metals in plants of different taxonomic groups from a montane rain forest in Sri Lanka. Environ. Geochem. Health 1996, 18, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.E.; Finoia, M.G.; Bocca, B.; Mele, G.; Alimonti, A.; Pino, A. Atmospheric background trace elements deposition in Tierra del Fuego region (Patagonia, Argentina), using transplanted Usnea barbata lichens. Environ. Monit. Assess. 2012, 184, 527–538. [Google Scholar] [CrossRef]
- Zambrano, A.; Nash, T.H. Lichen responses to short-term transplantation in Desierto de los Leones, Mexico City. Environ. Pollut. 2000, 107, 407–412. [Google Scholar] [CrossRef]
- Conti, M.E.; Jasan, R.; Finoia, M.G.; Iavicoli, I.; Plá, R. Trace elements deposition in the Tierra del Fuego region (south Patagonia) by using lichen transplants after the Puyehue-Cordón Caulle (north Patagonia) volcanic eruption in 2011. Environ. Sci. Pollut. Res. 2016, 23, 6574–6583. [Google Scholar] [CrossRef] [Green Version]
- Bergamaschi, L.; Rizzio, E.; Giaveri, G.; Loppi, S.; Gallorini, M. Comparison between the accumulation capacity of four lichen species transplanted to a urban site. Environ. Pollut. 2007, 148, 468–476. [Google Scholar] [CrossRef]
- Meli, M.A.; Desideri, D.; Cantaluppi, C.; Ceccotto, F.; Feduzi, L.; Roselli, C. Elemental and radiological characterization of commercial Cetraria islandica (L.) Acharius pharmaceutical and food supplementation products. Sci. Total Environ. 2018, 613–614, 1566–1572. [Google Scholar] [CrossRef] [PubMed]
- Rola, K. Insight into the pattern of heavy-metal accumulation in lichen thalli. J. Trace Elem. Med. Biol. 2020, 61, 126512. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, M.; Xu, B. A comprehensive review on secondary metabolites and health-promoting effects of edible lichen. J. Funct. Foods 2021, 80, 104283. [Google Scholar] [CrossRef]
- Kwong, S.P.; Wang, C. Review: Usnic acid-induced hepatotoxicity and cell death. Environ. Toxicol. Pharmacol. 2020, 80, 103493. [Google Scholar] [CrossRef] [PubMed]
- Marinescu, E.; Elisei, A.M.; Aprotosoaie, A.C.; Cioancă, O.; Trifan, A.; Miron, A.; Robu, S.; Ifrim, C.; Hăncianu, M. Assessment of heavy metals content in some medicinal plants and spices commonly used in Romania. Farmacia 2020, 68, 1099–1105. [Google Scholar] [CrossRef]
- Luo, L.; Wang, B.; Jiang, J.; Fitzgerald, M.; Huang, Q.; Yu, Z.; Li, H.; Zhang, J.; Wei, J.; Yang, C.; et al. Heavy Metal Contaminations in Herbal Medicines: Determination, Comprehensive Risk Assessments, and Solutions. Front. Pharmacol. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- FAO/WHO, General Standard for Contaminants and Toxins in Food and Feed. Codex Alimentarius Commission, Adopted in 1995. Revised in 1997, 2006, 2008, 2009 Amended in 2010, 2012, 2013, 2014, 2015. Available online: http://www.fao.org/fao-who-codexalimentarius/shproxy/en/?lnk=1&url=https%253A%252F%252Fworkspace.fao.org%252Fsites%252Fcodex%252FStandards%252FCXS%2B193-1995%252FCXS_193e.pdf (accessed on 2 April 2022).
- Union, E. Health and Food Safety; (EC) No 118/2006-Maximum Levels for Contaminants in Food Incl Mycotoxins. Off. J. Eur. Union 2006, 5–24. Available online: https://eur-lex.europa.eu/eli/reg/2021/1323/oj (accessed on 2 April 2022).
- FAO/WHO Guidelines for Assessing Quality of Herbal Medicines with Reference to Contaminants and Residues. WHO Libr. Cat. 2007, 118. Available online: https://apps.who.int/iris/bitstream/handle/10665/43510/9789241594448_eng.pdf?sequence=1&isAllowed=y (accessed on 2 April 2022).
- European Pharmacopoeia (Ph. Eur.) 10th Edition|EDQM European Directorate for the Quality of Medicines. Available online: https://www.edqm.eu/en/european-pharmacopoeia-ph-eur-10th-edition (accessed on 17 November 2020).
- Simkova, K.; Polesny, Z. Ethnobotanical review of wild edible plants used in the Czech Republic. J. Appl. Bot. Food Qual. 2015, 88, 49–67. [Google Scholar]
- Dobrescu, D.; Tanasescu, M.; Mezdrea, A.; Ivan, C.; Ordosch, E.; Neagoe, F.; Rizeanu, A.; Trifu, L.; Enescu, V. Contributions to the complex study of some lichens-Usnea genus. Pharmacological studies on Usnea barbata and Usnea hirta species. Rom. J. Physiol. 1993, 30, 101–107. [Google Scholar]
- Yang, Z.; Hu, Y.; Yue, P.; Luo, H.; Li, Q.; Li, H.; Zhang, Z.; Peng, F. Physicochemical Properties and Skin Protection Activities of Polysaccharides from Usnea longissima by Graded Ethanol Precipitation. ACS Omega 2021, 6, 25010–25018. [Google Scholar] [CrossRef]
- Thummajitsakul, S.; Samaikam, S.; Tacha, S.; Silprasit, K. Study on FTIR spectroscopy, total phenolic content, antioxidant activity and anti-amylase activity of extracts and different tea forms of Garcinia schomburgkiana leaves. LWT 2020, 134, 110005. [Google Scholar] [CrossRef]
- Zaini, E.; Nisak, R.K.; Utami, R.D.; Fitriani, L.; Ismed, F. Effect of milling on physicochemical properties of usnic acid isolated from Usnea sp. Orient. J. Chem. 2017, 33, 3031–3036. [Google Scholar] [CrossRef]
- Popovici, V.; Bucur, L.; Vochita, G.; Gherghel, D.; Mihai, C.T.; Rambu, D.; Calcan, S.I.; Costache, T.; Cucolea, I.E.; Matei, E.; et al. In vitro anticancer activity and oxidative stress biomarkers status determined by Usnea barbata (L.) F.H. Wigg. dry extracts. Antioxidants 2021, 10, 1141. [Google Scholar] [CrossRef] [PubMed]
- Kondratiuk, A.S.; Savchuk, O.M.; Hur, J.S. Optimization of protein extraction for lichen thalli. Mycobiology 2015, 43, 157–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Tang, H.; Zhang, Z. Ultramicromorphological observation of Usnea longissima Ach. Afr. J. Biotechnol. 2012, 11, 7102–7108. [Google Scholar]
- Rahman, M.M.; Islam, M.B.; Biswas, M.; Khurshid Alam, A.H.M. In vitro antioxidant and free radical scavenging activity of different parts of Tabebuia pallida growing in Bangladesh. BMC Res. Notes 2015, 8, 621. [Google Scholar] [CrossRef] [Green Version]
- Tirzitis, G.; Bartosz, G. Determination of antiradical and antioxidant activity: Basic principles and new insights. Acta Biochim. Pol. 2010, 57, 139–142. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | dUB | mcUB | ccUB | F-Value |
|---|---|---|---|---|
| Color Properties | ||||
| L* (adim.) | 63.02 ± 0.11 b | 59.08 ± 0.15 c | 70.29 ± 0.13 a | 5703.14 *** |
| a* (adim.) | −1.29 ± 0.02 b | −1.83 ± 0.03 c | 2.12 ± 0.17 a | 1414.01 *** |
| b* (adim.) | 13.60 ± 0.02 b | 12.52 ± 0.04 c | 13.92 ± 0.05 a | 1086.01 *** |
| hab (°) | 178.52 ± 0.00 c | 178.57 ± 0.00 b | 181.42 ± 0.01 a | 165,195.71 *** |
| C* (adim.) | 13.66 ± 0.02 b | 12.65 ± 0.05 c | 14.08 ± 0.04 a | 1125.62 *** |
| Element Content (μg/g) | Sample | F-Value | ||
|---|---|---|---|---|
| dUB | mcUB | ccUB | ||
| Al | 87.879 ± 1.152 b | 285.828 ± 2.748 a | 21.111 ± 0.608 c | 18,444.13 *** |
| As | ND | 0.219 ± 0.005 | ND | - |
| Ba | 3.782 ± 0.052 b | 12.113 ± 0.604 a | 0.596 ± 0.004 c | 866.03 *** |
| Ca | 979.766 ± 12.285 b | 1549.600 ± 18.406 a | 434.269 ± 22.222 c | 2846.40 *** |
| Cd | ND | 0.164 ± 0.001 | ND | - |
| Co | ND | 0.225 ± 0.001 | ND | - |
| Cr | 1.002 ± 0.008 b | 2.494 ± 0.010 a | 0.864 ± 0.007 c | 34,521.18 *** |
| Cu | 1.523 ± 0.013 b | 2.246 ± 0.005 a | 1.286 ± 0.014 c | 5771.15 *** |
| Fe | 52.561 ± 2.582 b | 282.468 ± 1.149 a | 33.107 ± 0.322 c | 21,398.25 *** |
| Li | ND | 0.185 ± 0.007 | ND | - |
| Mg | 172.721 ± 0.647 b | 524.239 ± 2.419 a | 53.482 ± 0.597 c | 81,353.06 *** |
| Mn | 101.425 ± 1.423 b | 354.041 ± 1.083 a | 29.098 ± 0.347 c | 78,943.06 *** |
| Ni | 0.449 ± 0.011 b | 1.033 ± 0.005 a | 0.355 ± 0.007 c | 5658.89 *** |
| Pb | 1.296 ± 0.007 b | 2.177 ± 0.014 a | 0.584 ± 0.004 c | 21,958.36 *** |
| V | 0.241 ± 0.004 b | 1.237 ± 0.005 a | ND | 92,152.36 *** |
| Zn | 20.536 ± 0.125 b | 33.223 ± 0.164 a | 13.588 ± 0.097 c | 17,179.89 *** |
| Hg | 0.671 ± 0.020 b | 0.708 ± 0.005 a | 0.539 ± 0.017 c | 99.48 *** |
| Sb | ND | 0.152 ± 0.003 | ND | - |
| Parameter | dUB | mcUB | ccUB | F-Value |
|---|---|---|---|---|
| TPC ethanol | 23.481 ± 0.039 b,x | 25.212 ± 0.084 a,y | 18.383 ± 0.004 c,x | 13200.49 *** |
| TPC acetone | 22.675 ± 0.108 b,y | 36.243 ± 0.093 a,x | 15.170 ± 0.129 c,y | 27784.01 *** |
| t-value | 12.16 *** | 152.42 *** | 43.12 ** | |
| AA acetone (% DPPH scavenging) | 16.878 ± 0.204 a,x | 12.747 ± 0.221 b,y | 11.336 ± 0.174 c,y | 10.96 ** |
| AA ethanol (% DPPH scavenging) | 15.985 ± 0.197 a,y | 15.735 ± 0.185 a,x | 15.080 ± 0.326 b,x | 618.35 ** |
| t-value | 5.45 ** | 17.96 *** | 17.55 *** |
| Legislative Authority | WHO/EU [60,61] | WHO [62] | Eu.Ph. [63] | Ch.Ph. [59] | Heavy Metals Content in U. barbata (μg/g) | ||
|---|---|---|---|---|---|---|---|
| Heavy Metal | Permissible Limits (μg/g) | dUB | mcUB | ccUB | |||
| Edible Plants | Medicinal Plants | ||||||
| As | 0.5 | 10 | 2 | ND | 0.219 | ND | |
| Cd | 0.02 | 0.2 | 1 | 0.3 | ND | 0.164 | ND |
| Cr | 1.3 | 2 | ND | 0.225 | ND | ||
| Co | 0.01 | 1.002 | 2.494 | 0.864 | |||
| Cu | 10 | 20 | 20 | 1.523 | 2.246 | 1.286 | |
| Hg | 0.03 | 1 | 0.1 | 0.2 | 0.671 | 0.708 | 0.539 |
| Ni | 0.1 | 0.449 | 1.033 | 0.355 | |||
| Pb | 2 | 10 | 5 | 1.296 | 2.177 | 0.584 | |
| V | 0.03 | 0.241 | 1.237 | ND | |||
| Zn | 50 | 50 | 20.536 | 33.223 | 13.588 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popovici, V.; Bucur, L.; Gîrd, C.E.; Calcan, S.I.; Cucolea, E.I.; Costache, T.; Rambu, D.; Ungureanu-Iuga, M.; Oroian, M.; Mironeasa, S.; et al. Advances in the Characterization of Usnea barbata (L.) Weber ex F.H. Wigg from Călimani Mountains, Romania. Appl. Sci. 2022, 12, 4234. https://doi.org/10.3390/app12094234
Popovici V, Bucur L, Gîrd CE, Calcan SI, Cucolea EI, Costache T, Rambu D, Ungureanu-Iuga M, Oroian M, Mironeasa S, et al. Advances in the Characterization of Usnea barbata (L.) Weber ex F.H. Wigg from Călimani Mountains, Romania. Applied Sciences. 2022; 12(9):4234. https://doi.org/10.3390/app12094234
Chicago/Turabian StylePopovici, Violeta, Laura Bucur, Cerasela Elena Gîrd, Suzana Ioana Calcan, Elena Iulia Cucolea, Teodor Costache, Dan Rambu, Mădălina Ungureanu-Iuga, Mircea Oroian, Silvia Mironeasa, and et al. 2022. "Advances in the Characterization of Usnea barbata (L.) Weber ex F.H. Wigg from Călimani Mountains, Romania" Applied Sciences 12, no. 9: 4234. https://doi.org/10.3390/app12094234
APA StylePopovici, V., Bucur, L., Gîrd, C. E., Calcan, S. I., Cucolea, E. I., Costache, T., Rambu, D., Ungureanu-Iuga, M., Oroian, M., Mironeasa, S., Schröder, V., Ozon, E. A., Caraiane, A., & Badea, V. (2022). Advances in the Characterization of Usnea barbata (L.) Weber ex F.H. Wigg from Călimani Mountains, Romania. Applied Sciences, 12(9), 4234. https://doi.org/10.3390/app12094234