
Current Research on the Effects of Non-Digestible Carbohydrates on Metabolic Disease

Abstract
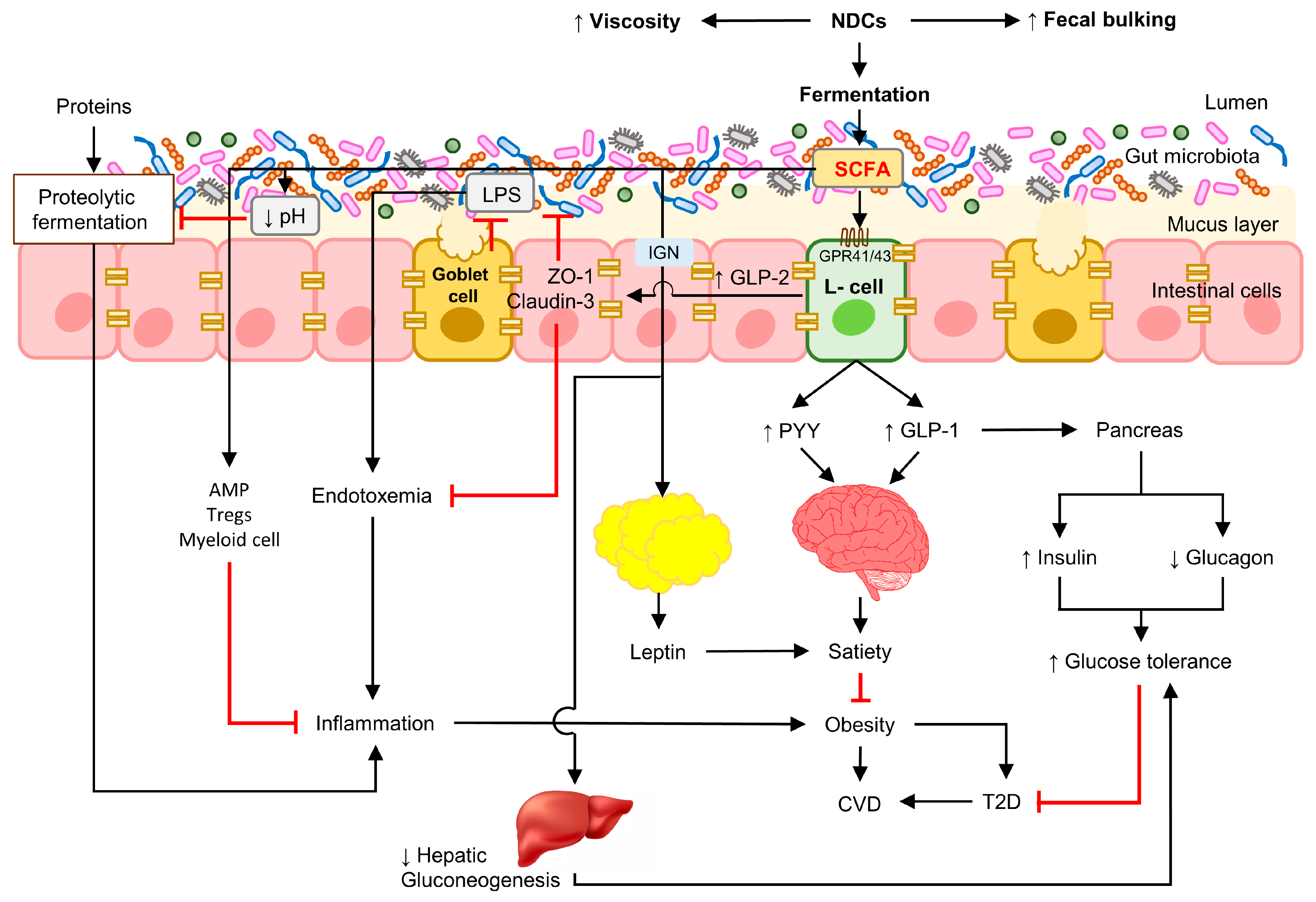
:1. Introduction
2. NDCs
3. NDC Characteristics Related to Health Benefits
4. NDCs and SCFAs
5. NDCs and MDs
5.1. Obesity
5.2. CVD
5.3. Diabetes

{kind=link}
| Types | Model | Dosage (g/day or %) | Duration (weeks) | Related Disease | Physiological Effects | References | |
|---|---|---|---|---|---|---|---|
| Soluble dietary fiber | |||||||
| Guar Gum | Human | ≥15 | 96 | L/M | ↓ Serum LDL-C and TC with cardiometabolic problems | [108] | |
| Pectin | |||||||
| Human | 650 or 1300 | 12 | Ob | ↓ Fasting BG, TG, cholesterols, AIP, HOMR-IR, insulin level, BW, body mass, leptin, and ghrelin. ↑ Adiponectin | [66] | |
| Human (Man) | 10 g | 3 h | IR | ↓ Plasma glucose, insulin, and iAUC | [101] | |
| Mice | 2% | 8 | Ob | ↓ Body and fat weight gain, dyslipidemea, hyperglycemia, and insulin resistance | [58] | |
| Rat (DB) | 0.25–2 (g/kg/day) | 4 | T2D | Improve glucose tolerance, hepaticglycogen content, BG, and blood lipid level. ↑ pAkt and ↓ GSK3β expression | [87] | ||
| Rat/Mice | 2–10% | 6–8 | Ob/NAFLD | Improve/restored adioistatic/adipokine sensitivity. Prevented the development of NAFLD, ↑ browning of adipose tissue | [57,59] | |
| Hydroxypropylmethylcellulose (HPMC) | Rat (ZDF) | 4–8% | 6 | DB/Ob | ↓ BG, urinary excretion of glucose, ketone bodies, epididymal fat pad, liver lipid, liver weight, adipose, and plasma cholesterol | [102,103] | |
| β-glucan | |||||||
| Human | 3–3.5 | 4 | CVD | ↓ LDL-c, TC, TC: HDL, non-HDL-c, and Framingham CVD risk | [81,82] | |
| Human | 3 or 5 | 5 | CVD | Change in microbiota profile: Bacteroides, Prevotella, and Dorea composition correlated with shifts of CVD risk factors: BMI, waist circumference, blood pressure, and TG levels | [80] | |
| Mice (HFD) | 2–5% | 12 | Ob | ↓ Weight gain and fat mass (2%), ↑ secretion of PYY and GLP-1 (5%) | [55] | ||
| Mice (HFD) | 0.4 (g/kg/day) | 10 | MD | ↓ IL-6 and IL-1β in plasma, ↑ HDL-c and ↓ BG, TC, LDL-c + VLDL-c, TG | [109] | |
| Psyllium husk | Human | 5 | 52 | Ob | ↓ BW | [67] | |
| Human (T2D) | 20 | 12 | T2D | ↓ BW, blood glucose, blood lipid, HbA1c, cholesterol, and TG | [88] | ||
| Human Human (NAFLD) | 9–10 10 | 8–10 12 | T2D/NAFLD NAFLD | ↓ Waist circumference, oxidized lipoproteins, calorie and carbohydrate intake, ALT, weight, and body fat ↓ BW, BMI | [68,110,111] | ||
| Inulin and inulin-type fructans | |||||||
| Human (Ob) | 8 | 12 | Ob | ↓ Hunger, desire to eat, and energy intake | [69] | |
| Human (T2D) | 0.1–10 | 6–8 | Ob/CVD | ↓ BW, waist circumference, serum TG, fat mass, fasting BG, HbA1c, LCL-c, TC/HDL-c and LDL-c/HDL-c | [70,71,112] | |
| Human (CVD) | 12 | 12 | CVD | ↓ IL-6 level, total p-cresyl sulfate (PCS) | [83] | |
| Rat | 32% | 12 | CVD | ↑ Hypertrophic of cardiomyocytes | [79] | ||
| Rat | 10% | 6 | Ob | ↓ Energy intake, BW, fat mass, plasma glucose, and GIP. ↑ PYY | [61] | ||
| Mice | 0.38 | 5 | Ob | ↓ Cecal content pH ad BW. ↑ Cecal SCFAs | [113] | ||
| High amylose starch (resistant starch II) | |||||||
| Human (T2D) | 6.8 or 25 | 8 | T2D | No significant different in fasting BG, ↓ fasting insulin level | [90] | |
| Human (women) | 0–30 | 4–6 h | IR | No significant different in fasting BG and insulin, ↓ the post-prandial glucose and insulin AUCs | [114,115] | ||
| Galactooligosaccharide (GOS) | Human | 5–18 | 2–3 | Ob | ↓ Colonic permeability, food intake, lipopolysaccharides, CRP, and BMI. ↑ antioxidative enzymes | [72,116] | |
| Mice | 0.083–0.83 | 6 | Ob/DB | ↓ BG, TC, TG, LDL-C, and liver lipid deposition. ↑ HDL-c, SCFAs | [104,105] | ||
| Polydextrose | Human (men) | 12 | 15–75 min | Ob | ↓ Energy intake in low protein group but not high protein group | [117] | |
| Insoluble dietary fiber | |||||||
| Cellulose | Rat | 10% | 24 | G/M | ↓ TG | [118] | |
| Soluble/Insoluble dietary fiber | |||||||
| Mixed plant cell wall fibers (corn starchhemicellulose) | Human | 10 g/day | 48 | T2D | Improve insulin release, peripheral insulin sensitivity, and blood glucose control | [106] | |
| Non-dietary fiber NDCs | |||||||
| Resistance starch III | Mice | 23% | 4 | T2D | Improve glucose and lipids profile (TC, TG, LDL, HDL) | [119] | |
| Resistance starch IV | Human | 30%/53.7% | 12/6 | CVD/T2D | Improve dyslipidemia and body composition. ↓ HbA1c, improve glycaemic control and cardiovascular risk without altering lipid metabolism | [84,85] | |
| Maltooligosaccharides (MOS) | Mice | 6 g/kg | 11 | Ob/DB | ↓ BW gain, adipose size, serum TC, TG, and insulin resistance | [62] | |
| Chitosan oligosaccharides (COS) | Mice | 200 mg/kg | 21 | MD | ↓ BG, TG, lipopolysaccharides, and adipose inflammation | [63] | |
| Bovine-milk oligosaccharides (BMO) | Mice | 6% | 6 | Ob | ↑ glucose tolerance, insulin secretion and HDL-C. ↓ BW, LBP, hepatic steatosis, gut permeability, total fat, mass, and adipocyte cell size | [64] | |
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pitsavos, C.; Panagiotakos, D.; Weinem, M.; Stefanadis, C. Diet, exercise and the metabolic syndrome. Rev. Diabet. Stud. 2006, 3, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhai, Y.; Zhao, J.; He, H.; Li, Y.; Liu, Y.; Feng, A.; Li, L.; Huang, T.; Xu, A.; et al. Impact of metabolic syndrome and it’s components on prognosis in patients with cardiovascular diseases: A meta-analysis. Front. Cardiovasc. Med. 2021, 8, 704145. [Google Scholar] [CrossRef] [PubMed]
- WHO. Noncommunicable Diseases Country Profiles 2018; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Saklayen, M.G. The global epidemic of the metabolic syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [Green Version]
- Batterham, R.L.; Cowley, M.A.; Small, C.J.; Herzog, H.; Cohen, M.A.; Dakin, C.L.; Wren, A.M.; Brynes, A.E.; Low, M.J.; Ghatei, M.A.; et al. Gut hormone PYY(3-36) physiologically inhibits food intake. Nature 2002, 418, 650–654. [Google Scholar] [CrossRef] [PubMed]
- D’Alessio, D.A.; Kahn, S.E.; Leusner, C.R.; Ensinck, J.W. Glucagon-like peptide 1 enhances glucose tolerance both by stimulation of insulin release and by increasing insulin-independent glucose disposal. J. Clin. Investig. 1994, 93, 2263–2266. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.P.; An, Z.; Wagner, C.; Lewis, A.G.; Cohen, E.B.; Li, B.; Mahbod, P.; Sandoval, D.; Perez-Tilve, D.; Tamarina, N.; et al. The role of β cell glucagon-like peptide-1 signaling in glucose regulation and response to diabetes drugs. Cell Metab. 2014, 19, 1050–1057. [Google Scholar] [CrossRef] [Green Version]
- Byrne, C.S.; Chambers, E.S.; Morrison, D.J.; Frost, G. The role of short chain fatty acids in appetite regulation and energy homeostasis. Int. J. Obes. 2015, 39, 1331–1338. [Google Scholar] [CrossRef] [Green Version]
- Brahe, L.K.; Astrup, A.; Larsen, L.H. Can we prevent obesity-related metabolic diseases by dietary modulation of the gut microbiota? Adv. Nutr. 2016, 7, 90–101. [Google Scholar] [CrossRef]
- Metabolic Syndrome: Mechanisms, Pathophysiology and Laboratory Assessment. Available online: https://www.abcam.com/content/metabolic-syndrome-mechanisms-pathophysiology-and-laboratory-assessment (accessed on 25 March 2022).
- Lim, S.; Eckel, R.H. Pharmacological treatment and therapeutic perspectives of metabolic syndrome. Rev. Endocr. Metab. Disord. 2014, 15, 329–341. [Google Scholar] [CrossRef]
- Lakka, T.A.; Laaksonen, D.E. Physical activity in prevention and treatment of the metabolic syndrome. Appl. Physiol. Nutr. Metab. 2007, 32, 76–88. [Google Scholar] [CrossRef]
- Feldeisen, S.E.; Tucker, K.L. Nutritional strategies in the prevention and treatment of metabolic syndrome. Appl. Physiol. Nutr. Metab. 2007, 32, 46–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Iglesia, R.; Loria-Kohen, V.; Zulet, M.A.; Martinez, J.A.; Reglero, G.; de Molina, A.R. Dietary strategies implicated in the prevention and treatment of metabolic syndrome. Int. J. Mol. Sci. 2016, 17, 1877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Diet, Nutrition and the Prevention of Chronic Diseases; World Health Organization: Geneva, Switzerland, 2003; Volume 916. [Google Scholar]
- Martínez-González, M.A.; Fernández-Jarne, E.; Serrano-Martínez, M.; Marti, A.; Martinez, J.A.; Martín-Moreno, J.M. Mediterranean diet and reduction in the risk of a first acute myocardial infarction: An operational healthy dietary score. Eur. J. Nutr. 2002, 41, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Yue, Q.; Wang, Z.; Tang, X.; Zhao, C.; Li, K.; Su, L.; Zhang, S.; Sun, X.; Liu, X.; Zhao, L. Hypolipidemic Effects of Fermented Seaweed Extracts by Saccharomyces cerevisiae and Lactiplantibacillus plantarum. Front. Microbiol. 2021, 12, 772585. [Google Scholar] [CrossRef] [PubMed]
- Gabbia, D.; De Martin, S. Brown seaweeds for the management of metabolic syndrome and associated diseases. Molecules 2020, 25, 4182. [Google Scholar] [CrossRef]
- FDA. Science Review of Isolated and Synthetic Non-Digestible Carbohydrates; U.S. Food Drug Administration: Silver Spring, MD, USA, 2016; 129p.
- McRae, M.P. Dietary Fiber Is Beneficial for the Prevention of Cardiovascular Disease: An Umbrella Review of Meta-analyses. J. Chiropr. Med. 2017, 16, 289–299. [Google Scholar] [CrossRef]
- Food and Drug Administration. Review of the Scientific Evidence on the Physiological Effects of Certain Non-Digestible Carbohydrates; Food Drug Administration: Silver Spring, MD, USA, 2018; pp. 1–52.
- Champ, M. Resistant starch. In Starch in Food: Structure, Function and Applications; CRC Press: Boca Raton, FL, USA, 2004; pp. 560–574. ISBN 9781855737310. [Google Scholar]
- Mudgil, D.; Barak, S. Composition, properties and health benefits of indigestible carbohydrate polymers as dietary fiber: A review. Int. J. Biol. Macromol. 2013, 61, 1–6. [Google Scholar] [CrossRef]
- Phillips, G.O.; Cui, S.W. An introduction: Evolution and finalisation of the regulatory definition of dietary fibre. Food Hydrocoll. 2011, 25, 139–143. [Google Scholar] [CrossRef]
- Williams, B.A.; Mikkelsen, D.; Flanagan, B.M.; Gidley, M.J. “Dietary fibre”: Moving beyond the “soluble/insoluble” classification for monogastric nutrition, with an emphasis on humans and pigs. J. Anim. Sci. Biotechnol. 2019, 10, 45. [Google Scholar] [CrossRef] [Green Version]
- Viuda-Martos, M.; López-Marcos, M.C.; Fernández-López, J.; Sendra, E.; López-Vargas, J.H.; Perez-Álvarez, J.A. Role of fiber in cardiovascular diseases: A review. Compr. Rev. Food Sci. Food Saf. 2010, 9, 240–258. [Google Scholar] [CrossRef]
- Mudgil, D.; Barak, S.; Khatkar, B.S. Guar gum: Processing, properties and food applications—A Review. J. Food Sci. Technol. 2014, 51, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, M.M.; Aldughpassi, A.D.H.; Sidhu, J.S.; Al-Foudari, M.Y.; Al-Othman, A.R.A. Effect of psyllium husk addition on the instrumental texture and consumer acceptability of high-fiber wheat pan bread and buns. Ann. Agric. Sci. 2021, 66, 75–80. [Google Scholar] [CrossRef]
- Shi, Z.; Zhang, Y.; Phillips, G.O.; Yang, G. Utilization of bacterial cellulose in food. Food Hydrocoll. 2014, 35, 539–545. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Steed, H.; Macfarlane, S. Bacterial metabolism and health-related effects of galacto-oligosaccharides and other prebiotics. J. Appl. Microbiol. 2008, 104, 305–344. [Google Scholar] [CrossRef] [PubMed]
- Homayouni, A.; Amini, A.; Keshtiban, A.K.; Mortazavian, A.M.; Esazadeh, K.; Pourmoradian, S. Resistant starch in food industry: A changing outlook for consumer and producer. Starch-Stärke 2014, 66, 102–114. [Google Scholar] [CrossRef]
- Marlett, J.A. Sites and mechanisms for the hypocholesterolemic actions of soluble dietary fiber sources. Adv. Exp. Med. Biol. 1997, 427, 109–121. [Google Scholar] [CrossRef]
- Streppel, M.T.; Arends, L.R.; van ’t Veer, P.; Grobbee, D.E.; Geleijnse, J.M. Dietary fiber and blood pressure: A meta-analysis of randomized placebo-controlled trials. Arch. Intern. Med. 2005, 165, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Institute of Medicine (US) Standing Committee on the Scientific Evaluation of Dietary Reference Intakes. Dietary Reference Intakes. In Dietary Reference Intakes for Calcium, Phosphorus, Magnesium, Vitamin D and Fluoride; National Academies Press: Washington, DC, USA, 1997; Volume 55, ISBN 030908525X. [Google Scholar]
- Mann, J.I.; Cummings, J.H. Possible implications for health of the different definitions of dietary fibre. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 226–229. [Google Scholar] [CrossRef]
- Prado, S.B.R.; Castro-Alves, V.C.; Ferreira, G.F.; Fabi, J.P. Ingestion of Non-digestible carbohydrates from plant-source foods and decreased risk of colorectal cancer: A review on the biological effects and the mechanisms of action. Front. Nutr. 2019, 6, 72. [Google Scholar] [CrossRef]
- Myhrstad, M.C.W.; Tunsjø, H.; Charnock, C.; Telle-Hansen, V.H. Dietary Fiber, Gut Microbiota, and Metabolic Regulation—Current Status in Human Randomized Trials. Nutrients 2020, 12, 859. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.H. Control of lymphocyte functions by gut microbiota-derived short-chain fatty acids. Cell. Mol. Immunol. 2021, 18, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The impact of dietary fiber on gut microbiota in host health and disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, E.S.; Byrne, C.S.; Morrison, D.J.; Murphy, K.G.; Preston, T.; Tedford, C.; Garcia-Perez, I.; Fountana, S.; Serrano-Contreras, J.I.; Holmes, E.; et al. Dietary supplementation with inulin-propionate ester or inulin improves insulin sensitivity in adults with overweight and obesity with distinct effects on the gut microbiota, plasma metabolome and systemic inflammatory responses: A randomised cross-over t. Gut 2019, 68, 1430–1438. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; van der Beek, C.M.; Jocken, J.W.E.; Goossens, G.H.; Holst, J.J.; Olde Damink, S.W.M.; Lenaerts, K.; Dejong, C.H.C.; Blaak, E.E. Colonic infusions of short-chain fatty acid mixtures promote energy metabolism in overweight/obese men: A randomized crossover trial. Sci. Rep. 2017, 7, 2360. [Google Scholar] [CrossRef]
- Savage, A.P.; Adrian, T.E.; Carolan, G.; Chatterjee, V.K.; Bloom, S.R. Effects of peptide YY (PYY) on mouth to caecum intestinal transit time and on the rate of gastric emptying in healthy volunteers. Gut 1987, 28, 166–170. [Google Scholar] [CrossRef]
- De Vadder, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef] [Green Version]
- Nie, Q.; Hu, J.; Gao, H.; Fan, L.; Chen, H.; Nie, S. Polysaccharide from Plantago asiatica L. attenuates hyperglycemia, hyperlipidemia and affects colon microbiota in type 2 diabetic rats. Food Hydrocoll. 2019, 86, 34–42. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Allison, C.; Gibson, G.R. Effect of pH on protease activities in the large intestine. Lett. Appl. Microbiol. 2008, 7, 161–164. [Google Scholar] [CrossRef]
- Verspreet, J.; Damen, B.; Broekaert, W.F.; Verbeke, K.; Delcour, J.A.; Courtin, C.M. A critical look at prebiotics within the dietary fiber concept. Annu. Rev. Food Sci. Technol. 2016, 7, 167–190. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; O’Riordan, M.X.D. Regulation of bacterial pathogenesis by intestinal short-chain fatty acids. Adv. Appl. Microbiol. 2013, 85, 93–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cani, P.D.; Possemiers, S.; Van de Wiele, T.; Guiot, Y.; Everard, A.; Rottier, O.; Geurts, L.; Naslain, D.; Neyrinck, A.; Lambert, D.M.; et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut 2009, 58, 1091–1103. [Google Scholar] [CrossRef] [Green Version]
- Yeh, T.-L.; Chen, H.-H.; Tsai, S.-Y.; Lin, C.-Y.; Liu, S.-J.; Chien, K.-L. The Relationship between metabolically healthy obesity and the risk of cardiovascular disease: A systematic review and meta-analysis. J. Clin. Med. 2019, 8, 1228. [Google Scholar] [CrossRef] [Green Version]
- Ruhee, R.; Suzuki, K. Dietary fiber and its effect on obesity: A Review Article. Adv. Med. Res. 2018, 1, 1–13. [Google Scholar] [CrossRef]
- Alexander, C.; Swanson, K.S.; Fahey, G.C.; Garleb, K.A. Perspective: Physiologic Importance of Short-Chain Fatty Acids from Nondigestible Carbohydrate Fermentation. Adv. Nutr. 2019, 10, 576–589. [Google Scholar] [CrossRef]
- Miyamoto, J.; Watanabe, K.; Taira, S.; Kasubuchi, M.; Li, X.; Irie, J.; Itoh, H.; Kimura, I. Barley β-glucan improves metabolic condition via short-chain fatty acids produced by gut microbial fermentation in high fat diet fed mice. PLoS ONE 2018, 13, e0196579. [Google Scholar] [CrossRef]
- Islam, A.; Civitarese, A.E.; Hesslink, R.L.; Gallaher, D.D. Viscous dietary fiber reduces adiposity and plasma leptin and increases muscle expression of fat oxidation genes in rats. Obesity 2012, 20, 349–355. [Google Scholar] [CrossRef]
- García-Carrizo, F.; Picó, C.; Rodríguez, A.M.; Palou, A. High-esterified pectin reverses metabolic malprogramming, improving sensitivity to adipostatic/adipokine hormones. J. Agric. Food Chem. 2019, 67, 3633–3642. [Google Scholar] [CrossRef]
- Zhan, J.; Liang, Y.; Liu, D.; Ma, X.; Li, P.; Zhai, W.; Zhou, Z.; Wang, P. Pectin reduces environmental pollutant-induced obesity in mice through regulating gut microbiota: A case study of p,p′-DDE. Environ. Int. 2019, 130, 104861. [Google Scholar] [CrossRef] [PubMed]
- Houron, C.; Ciocan, D.; Trainel, N.; Mercier-Nomé, F.; Hugot, C.; Spatz, M.; Perlemuter, G.; Cassard, A.M. Gut microbiota reshaped by pectin treatment improves liver steatosis in obese mice. Nutrients 2021, 13, 3725. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Scholte, J.; Borewicz, K.; van den Bogert, B.; Smidt, H.; Scheurink, A.J.W.; Gruppen, H.; Schols, H.A. Effects of pectin supplementation on the fermentation patterns of different structural carbohydrates in rats. Mol. Nutr. Food Res. 2016, 60, 2256–2266. [Google Scholar] [CrossRef] [PubMed]
- Cluny, N.L.; Eller, L.K.; Keenan, C.M.; Reimer, R.A.; Sharkey, K.A. Interactive effects of oligofructose and obesity predisposition on gut hormones and microbiota in diet-induced obese rats. Obesity 2015, 23, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, X.; Wang, S.; Li, H.; Lu, Z.; Shi, J.; Xu, Z. Mannan-oligosaccharide modulates the obesity and gut microbiota in high-fat diet-fed mice. Food Funct. 2018, 9, 3916–3929. [Google Scholar] [CrossRef]
- Zheng, J.; Cheng, G.; Li, Q.; Jiao, S.; Feng, C.; Zhao, X.; Yin, H.; Du, Y.; Liu, H. Chitin oligosaccharide modulates gut microbiota and attenuates high-fat-diet-induced metabolic syndrome in mice. Mar. Drugs 2018, 16, 66. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, M.K.; Ronveaux, C.C.; Rust, B.M.; Newman, J.W.; Hawley, M.; Barile, D.; Mills, D.A.; Raybould, H.E. Prebiotic milk oligosaccharides prevent development of obese phenotype, impairment of gut permeability, and microbial dysbiosis in high fat-fed mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G474–G487. [Google Scholar] [CrossRef]
- Isken, F.; Klaus, S.; Osterhoff, M.; Pfeiffer, A.F.H.; Weickert, M.O. Effects of long-term soluble vs. insoluble dietary fiber intake on high-fat diet-induced obesity in C57BL/6J mice. J. Nutr. Biochem. 2010, 21, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Capomolla, A.S.; Janda, E.; Paone, S.; Parafati, M.; Sawicki, T.; Mollace, R.; Ragusa, S.; Mollace, V. Atherogenic index reduction and weight loss in metabolic syndrome patients treated with A Novel Pectin-Enriched Formulation of Bergamot Polyphenols. Nutrients 2019, 11, 1271. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.; Ho, S.; Gahler, R.J.; Wood, S. Effect on body weight and composition in overweight/obese Australian adults over 12 months consumption of two different types of fibre supplementation in a randomized trial. Nutr. Metab. 2016, 13, 82. [Google Scholar] [CrossRef] [Green Version]
- Akbarian, S.-A.; Asgary, S.; Feizi, A.; Iraj, B.; Askari, G. Comparative study on the effect of Plantago psyllium and Ocimum basilicum seeds on anthropometric measures in nonalcoholic fatty liver patients. Int. J. Prev. Med. 2016, 7, 114. [Google Scholar] [CrossRef] [PubMed]
- Reimer, R.A.; Willis, H.J.; Tunnicliffe, J.M.; Park, H.; Madsen, K.L.; Soto-Vaca, A. Inulin-type fructans and whey protein both modulate appetite but only fructans alter gut microbiota in adults with overweight/obesity: A randomized controlled trial. Mol. Nutr. Food Res. 2017, 61, 1700484. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.M.; da Silva, N.B.M.; Chaves, J.B.P.; Alfenas, R.d.C.G. Consumption of yacon flour improves body composition and intestinal function in overweight adults: A randomized, double-blind, placebo-controlled clinical trial. Clin. Nutr. ESPEN 2019, 29, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolucci, A.C.; Hume, M.P.; Martínez, I.; Mayengbam, S.; Walter, J.; Reimer, R.A. Prebiotics reduce body fat and alter intestinal microbiota in children who are overweight or with obesity. Gastroenterology 2017, 153, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Morel, F.B.; Dai, Q.; Ni, J.; Thomas, D.; Parnet, P.; Fança-Berthon, P. α-Galacto-oligosaccharides dose-dependently reduce appetite and decrease inflammation in overweight adults. J. Nutr. 2015, 145, 2052–2059. [Google Scholar] [CrossRef] [Green Version]
- Buil-Cosiales, P.; Zazpe, I.; Toledo, E.; Corella, D.; Salas-Salvadó, J.; Diez-Espino, J.; Ros, E.; Fernandez-Creuet Navajas, J.; Santos-Lozano, J.M.; Arós, F.; et al. Fiber intake and all-cause mortality in the Prevención con Dieta Mediterránea (PREDIMED) study. Am. J. Clin. Nutr. 2014, 100, 1498–1507. [Google Scholar] [CrossRef]
- Kokubo, Y.; Iso, H.; Saito, I.; Yamagishi, K.; Ishihara, J.; Inoue, M.; Tsugane, S.; JPHC Study Group. Dietary fiber intake and risk of cardiovascular disease in the Japanese population: The Japan Public Health Center-based study cohort. Eur. J. Clin. Nutr. 2011, 65, 1233–1241. [Google Scholar] [CrossRef] [Green Version]
- Soliman, G.A. Dietary Fiber, Atherosclerosis, and Cardiovascular Disease. Nutrients 2019, 11, 1155. [Google Scholar] [CrossRef] [Green Version]
- Buttar, H.S.; Li, T.; Ravi, N. Prevention of cardiovascular diseases: Role of exercise, dietary interventions, obesity and smoking cessation. Exp. Clin. Cardiol. 2005, 10, 229–249. [Google Scholar]
- Al-Lahham, S.H.; Peppelenbosch, M.P.; Roelofsen, H.; Vonk, R.J.; Venema, K. Biological effects of propionic acid in humans; metabolism, potential applications and underlying mechanisms. Biochim. Biophys. Acta 2010, 1801, 1175–1183. [Google Scholar] [CrossRef]
- Lee Kennedy, R.; Vangaveti, V.; Jarrod, G.; Shashidhar, V.; Shashidhar, V.; Baune, B.T. Review: Free fatty acid receptors: Emerging targets for treatment of diabetes and its complications. Ther. Adv. Endocrinol. Metab. 2010, 1, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Sarfaraz, S.; Singh, S.; Hawke, A.; Clarke, S.T.; Ramdath, D.D. Effects of High-Fat Diet Induced Obesity and Fructooligosaccharide Supplementation on Cardiac Protein Expression. Nutrients 2020, 12, 3404. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ames, N.P.; Tun, H.M.; Tosh, S.M.; Jones, P.J.; Khafipour, E. High molecular weight barley β-glucan alters gut microbiota toward reduced cardiovascular disease risk. Front. Microbiol. 2016, 7, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolever, T.M.S.; Rahn, M.; Dioum, E.; Spruill, S.E.; Ezatagha, A.; Campbell, J.E.; Jenkins, A.L.; Chu, Y. An oat β-glucan beverage reduces LDL cholesterol and cardiovascular disease risk in men and women with borderline high cholesterol: A double-blind, randomized, controlled clinical trial. J. Nutr. 2021, 151, 2655–2666. [Google Scholar] [CrossRef]
- Ho, H.V.T.; Sievenpiper, J.L.; Zurbau, A.; Blanco Mejia, S.; Jovanovski, E.; Au-Yeung, F.; Jenkins, A.L.; Vuksan, V. The effect of oat β-glucan on LDL-cholesterol, non-HDL-cholesterol and apoB for CVD risk reduction: A systematic review and meta-analysis of randomised-controlled trials. Br. J. Nutr. 2016, 116, 1369–1382. [Google Scholar] [CrossRef] [Green Version]
- Armani, R.G.; Carvalho, A.B.; Ramos, C.I.; Hong, V.; Bortolotto, L.A.; Cassiolato, J.L.; Oliveira, N.F.; Cieslarova, Z.; do Lago, C.L.; Klassen, A.; et al. Effect of fructooligosaccharide on endothelial function in CKD patients: A randomized controlled trial. Nephrol. Dial. Transplant. 2022, 37, 85–91. [Google Scholar] [CrossRef]
- Nichenametla, S.N.; Weidauer, L.A.; Wey, H.E.; Beare, T.M.; Specker, B.L.; Dey, M. Resistant starch type 4-enriched diet lowered blood cholesterols and improved body composition in a double blind controlled cross-over intervention. Mol. Nutr. Food Res. 2014, 58, 1365–1369. [Google Scholar] [CrossRef]
- Mesa García, M.D.; García-Rodríguez, C.E.; de la Cruz Rico, M.; Aguilera, C.M.; Pérez-Rodríguez, M.; Pérez-de-la-Cruz, A.J.; Gil, Á. A new fructose-free, resistant-starch type IV-enriched enteral formula improves glycaemic control and cardiovascular risk biomarkers when administered for six weeks to elderly diabetic patients. Nutr. Hosp. 2017, 34, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Hartley, L.; May, M.D.; Loveman, E.; Colquitt, J.L.; Rees, K. Dietary fibre for the primary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2016, 1, CD011472. [Google Scholar] [CrossRef]
- Ibrügger, S.; Kristensen, M.; Poulsen, M.W.; Mikkelsen, M.S.; Ejsing, J.; Jespersen, B.M.; Dragsted, L.O.; Engelsen, S.B.; Bügel, S. Extracted Oat and Barley β-Glucans Do Not Affect Cholesterol Metabolism in Young Healthy Adults. J. Nutr. 2013, 143, 1579–1585. [Google Scholar] [CrossRef] [Green Version]
- Noureddin, S.; Mohsen, J.; Payman, A. Effects of psyllium vs. placebo on constipation, weight, glycemia, and lipids: A randomized trial in patients with type 2 diabetes and chronic constipation. Complement. Ther. Med. 2018, 40, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Darooghegi Mofrad, M.; Mozaffari, H.; Mousavi, S.M.; Sheikhi, A.; Milajerdi, A. The effects of psyllium supplementation on body weight, body mass index and waist circumference in adults: A systematic review and dose-response meta-analysis of randomized controlled trials. Crit. Rev. Food Sci. Nutr. 2020, 60, 859–872. [Google Scholar] [CrossRef] [PubMed]
- Dainty, S.A.; Klingel, S.L.; Pilkey, S.E.; McDonald, E.; McKeown, B.; Emes, M.J.; Duncan, A.M. Resistant Starch Bagels Reduce Fasting and Postprandial Insulin in Adults at Risk of Type 2 Diabetes. J. Nutr. 2016, 146, 2252–2259. [Google Scholar] [CrossRef] [Green Version]
- Post, R.E.; Mainous, A.G., 3rd; King, D.E.; Simpson, K.N. Dietary fiber for the treatment of type 2 diabetes mellitus: A meta-analysis. J. Am. Board Fam. Med. 2012, 25, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Lewis, G.; Wang, B.; Shafiei Jahani, P.; Hurrell, B.P.; Banie, H.; Aleman Muench, G.R.; Maazi, H.; Helou, D.G.; Howard, E.; Galle-Treger, L.; et al. Dietary Fiber-induced microbial short chain fatty acids suppress ILC2-dependent airway inflammation. Front. Immunol. 2019, 10, 2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jovanovski, E.; Khayyat, R.; Zurbau, A.; Komishon, A.; Mazhar, N.; Sievenpiper, J.L.; Blanco Mejia, S.; Ho, H.V.T.; Li, D.; Jenkins, A.L.; et al. Should viscous fiber supplements be considered in diabetes control? Results from a systematic review and meta-analysis of randomized controlled trials. Diabetes Care 2019, 42, 755–766. [Google Scholar] [CrossRef]
- Davison, K.M.; Temple, N.J. Cereal fiber, fruit fiber, and type 2 diabetes: Explaining the paradox. J. Diabetes Complicat. 2018, 32, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Slavin, J.L. Dietary fiber and body weight. Nutrition 2005, 21, 411–418. [Google Scholar] [CrossRef]
- Mudgil, D. The Interaction between Insoluble and Soluble Fiber. In Dietary Fiber for the Prevention of Cardiovascular Disease; Academic Press: Cambridge, MA, USA, 2017; pp. 35–59. ISBN 9780128051306. [Google Scholar]
- Weickert, M.O.; Pfeiffer, A.F.H. Metabolic effects of dietary fiber consumption and prevention of diabetes. J. Nutr. 2008, 138, 439–442. [Google Scholar] [CrossRef]
- Weickert, M.O.; Pfeiffer, A.F.H. Impact of dietary fiber consumption on insulin resistance and the prevention of type 2 diabetes. J. Nutr. 2018, 148, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Consortium, T.I. Dietary fibre and incidence of type 2 diabetes in eight European countries: The EPIC-InterAct Study and a meta-analysis of prospective studies. Diabetologia 2015, 58, 1394–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandalia, M.; Garg, A.; Lutjohann, D.; von Bergmann, K.; Grundy, S.M.; Brinkley, L.J. Beneficial effects of high dietary fiber intake in patients with type 2 diabetes mellitus. N. Engl. J. Med. 2000, 342, 1392–1398. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.; Gu, X.; Stebbins, N.; Crandall, P.; Ricke, S.; Lee, S. Effects of Soybean Pectin on Blood Glucose and Insulin Responses in Healthy Men; University of Arkansas: Little Rock, AR, USA, 2015. [Google Scholar] [CrossRef]
- Brockman, D.A.; Chen, X.; Gallaher, D.D. Hydroxypropyl methylcellulose, a viscous soluble fiber, reduces insulin resistance and decreases fatty liver in Zucker Diabetic Fatty rats. Nutr. Metab. 2012, 9, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, S.C.; Anderson, W.H.K.; Albers, D.R.; Langhorst, M.L.; Young, S.A. Effect of hydroxypropyl methylcellulose on obesity and glucose metabolism in a diet-induced obesity mouse model. J. Diabetes 2011, 3, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Lyu, W.; Xie, M.; Yuan, Q.; Ye, H.; Hu, B.; Zhou, L.; Zeng, X. Effects of α-Galactooligosaccharides from Chickpeas on High-Fat-Diet-Induced Metabolic Syndrome in Mice. J. Agric. Food Chem. 2017, 65, 3160–3166. [Google Scholar] [CrossRef]
- Sangwan, V.; Tomar, S.K.; Ali, B.; Singh, R.R.B.; Singh, A.K. Hypoglycaemic effect of galactooligosaccharides in alloxan-induced diabetic rats. J. Dairy Res. 2015, 82, 70–77. [Google Scholar] [CrossRef]
- Hanai, H.; Ikuma, M.; Sato, Y.; Iida, T.; Hosoda, Y.; Matsushita, I.; Nogaki, A.; Yamada, M.; Kaneko, E. Long-term Effects of Water-soluble Corn Bran Hemicellulose on Glucose Tolerance in Obese and Non-obese Patients: Improved Insulin Sensitivity and Glucose Metabolism in Obese Subjects. Biosci. Biotechnol. Biochem. 1997, 61, 1358–1361. [Google Scholar] [CrossRef]
- Chen, C.; Zeng, Y.; Xu, J.; Zheng, H.; Liu, J.; Fan, R.; Zhu, W.; Yuan, L.; Qin, Y.; Chen, S.; et al. Therapeutic effects of soluble dietary fiber consumption on type 2 diabetes mellitus. Exp. Ther. Med. 2016, 12, 1232–1242. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Sun, Y.; Santos, H.O.; Găman, M.A.; Bhat, L.T.; Cui, Y. Effects of guar gum supplementation on the lipid profile: A systematic review and meta-analysis of randomized controlled trials. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 3271–3281. [Google Scholar] [CrossRef]
- Chen, G.; Chen, D.; Zhou, W.; Peng, Y.; Chen, C.; Shen, W.; Zeng, X.; Yuan, Q. Improvement of metabolic Syndrome in High-Fat Diet-Induced Mice by Yeast β-Glucan Is Linked to Inhibited Proliferation of Lactobacillus and Lactococcus in gut microbiota. J. Agric. Food Chem. 2021, 69, 7581–7592. [Google Scholar] [CrossRef]
- Akbarzadeh, Z.; Nourian, M.; Askari, G.; Maracy, M.R. The effect of Psyllium on body composition measurements and liver enzymes in overweight or obese adults with nonalcoholic fatty liver disease (NAFLD). Int. J. Adv. Biotechnol. Res. 2016, 7, 1545–1554. [Google Scholar]
- Ricklefs-Johnson, K.; Johnston, C.S.; Sweazea, K.L. Ground flaxseed increased nitric oxide levels in adults with type 2 diabetes: A randomized comparative effectiveness study of supplemental flaxseed and psyllium fiber. Obes Med. 2017, 5, 16–24. [Google Scholar] [CrossRef]
- Aliasgharzadeh, A.; Khalili, M.; Mirtaheri, E.; Pourghassem Gargari, B.; Tavakoli, F.; Abbasalizad Farhangi, M.; Babaei, H.; Dehghan, P. A Combination of Prebiotic Inulin and Oligofructose Improve Some of Cardiovascular Disease Risk Factors in Women with Type 2 Diabetes: A Randomized Controlled Clinical Trial. Adv. Pharm. Bull. 2015, 5, 507–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huazano-García, A.; Shin, H.; López, M.G. Modulation of Gut Microbiota of Overweight Mice by Agavins and Their Association with Body Weight Loss. Nutrients 2017, 9, 821. [Google Scholar] [CrossRef] [Green Version]
- Gower, B.A.; Bergman, R.; Stefanovski, D.; Darnell, B.; Ovalle, F.; Fisher, G.; Sweatt, S.K.; Resuehr, H.S.; Pelkman, C. Baseline insulin sensitivity affects response to high-amylose maize resistant starch in women: A randomized, controlled trial. Nutr. Metab. 2016, 13, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahat-Rozenbloom, S.; Fernandes, J.; Cheng, J.; Gloor, G.B.; Wolever, T.M.S. The acute effects of inulin and resistant starch on postprandial serum short-chain fatty acids and second-meal glycemic response in lean and overweight humans. Eur. J. Clin. Nutr. 2017, 71, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Krumbeck, J.A.; Rasmussen, H.E.; Hutkins, R.W.; Clarke, J.; Shawron, K.; Keshavarzian, A.; Walter, J. Probiotic Bifidobacterium strains and galactooligosaccharides improve intestinal barrier function in obese adults but show no synergism when used together as synbiotics. Microbiome 2018, 6, 121. [Google Scholar] [CrossRef]
- Soong, Y.Y.; Lim, W.X.; Leow, M.K.S.; Siow, P.C.; Teh, A.L.; Henry, C.J. Combination of soya protein and polydextrose reduces energy intake and glycaemic response via modulation of gastric emptying rate, ghrelin and glucagon-like peptide-1 in Chinese. Br. J. Nutr. 2016, 115, 2130–2137. [Google Scholar] [CrossRef] [Green Version]
- Pastuszewska, B.; Taciak, M.; Tuśnio, A.; Misztal, T.; Ochtabińska, A. Physiological effects of long-term feeding diets supplemented with potato fibre or cellulose to adult rats. Arch. Anim. Nutr. 2010, 64, 155–169. [Google Scholar] [CrossRef]
- Nugraheni, M.; Hamidah, S.; Auliana, R. A potential of coleus tuberosus crackers rich in resistant starch type 3 improves glucose and lipid profile of alloxan –induced diabetic mice. Curr. Res. Nutr. Food Sci. 2017, 5, 308–319. [Google Scholar] [CrossRef]
- Armstrong, H.; Mander, I.; Zhang, Z.; Armstrong, D.; Wine, E. Not All Fibers Are Born Equal; Variable Response to Dietary Fiber Subtypes in IBD. Front. Pediatr. 2021, 8, 924. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, C.M.; Livingston, K.A.; Obin, M.; Roberts, S.B.; Chung, M.; McKeown, N.M. Dietary fiber and the human gut microbiota: Application of evidence mapping methodology. Nutrients 2017, 9, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, B.A.; Grant, L.J.; Gidley, M.J.; Mikkelsen, D. Gut fermentation of dietary fibres: Physico-chemistry of plant cell walls and implications for health. Int. J. Mol. Sci. 2017, 18, 2203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, J.; Kim, S.; Cabatbat, J.J.T.; Jang, S.; Jin, Y.S.; Jung, G.Y.; Chia, N.; Kim, P.J. Global metabolic interaction network of the human gut microbiota for context-specific community-scale analysis. Nat. Commun. 2017, 8, 15393. [Google Scholar] [CrossRef] [Green Version]

| Type | Structure | Source | Properties | References | |
|---|---|---|---|---|---|
| Soluble dietary fiber | |||||
| Guar Gum | A linear chain of β 1,4-linked mannose residues to which galactose residues are 1,6-linked at every second mannose | Seeds of the drought tolerant plant | Gel-forming, thickening, and stabilization. | [27] | |
| Locust Bean Gum | Galactomannan composed of galactose and mannose units combined through glycosidic linkages | Seeds of the carob tree | Film-forming. | [21] | |
| Pectin | Chain of α-(1 → 4)-linked d-galacturonic acid units interrupted by the insertion of (1 → 2)-linked l-rhamnopyranosyl residues in adjacent or alternate positions | Cell walls and intracellular tissues of fruits and vegetables | Emulsifier, gelling agent, thickener, stabilizer, and fat or sugar replacer in low-calorie foods | [23] | |
| Hydroxypropylmethylcellulose (HPMC) | Propylene glycol ether of methylcellulose | Film forming, stabilizing, and thickening. | [21] | ||
| β-Glucan | Mixed-linkage polysaccharide (1 → 3), (1 → 4) β-d-glucan | Cell walls of oats, barley, rye and wheat | Altering foods structure, texture, and viscosity | [23] | |
| Psyllium husk | Arabinoxylan with (1 → 4) and (1 → 3) xylopyranose backbones | Seeds of Plantango ovata | Gel-forming, produce low-calorie, and high fiber foods | [28] | |
| Arabinoxylan | Diversely composed (1 → 4)-β-d-xylan polymer | Wheat | Film-forming, balance of carbohydrate-rich foods, improve the viscosity, texture, sensory characteristics, and shelf-life of food products | [24] | |
| Alginate | Linear unbranched polysaccharides which contain different amounts of (1 → 4′)-linked β-d-mannuronic acid and α-l-guluronic acid residues | Brown seaweeds | Gelling, viscosifying, and stabilizing | [21] | |
| Inulin and inulin-type fructans | A mixture of linear fructose polymers with different chain length and a glucose molecule at each C2 end | Chicory roots | Bulking agent in foods, improve the texture, mouthfeel, taste, and replace sugar or fat. | [21] | |
| High amylose starch (resistant starch II) | d-Glucose units linked by R-1,4/R-1,6 glucosidic bonds | Raw starch (green banana and raw potatoes) | Increase of food’s functional properties does not change its sensory characteristics. | [22] | |
| Galactooligosaccharide (GOS) | β-Linked galactose moieties with galactose or glucose at the reducing end. | Soybeans and lactose from cow’s milk | Improve the texture of foods and as a bulking agent. | [30] | |
| Polydextrose | A polysaccharide composed of randomly cross-linked glucose units with all types of glycosidic bonding | Produced from the naturally occurring components: glucose, sorbitol, and citric acid | Bulking agent, stabilizer, thickener, and humectant | [21] | |
| Resistant maltodextrin/dextrin (resistant starch V) | Oligosaccharides of glucose molecules that are joined by digestible linkages and non-digestible α-1,2 and α-1,3 linkages | Corn, wheat, potato, and tapioca | Increase the nutritional value of food | [31] | |
| Insoluble dietary fiber | |||||
| Cellulose | Linear homopolymer of β-(1 → 4) linked β-d-glucose residues | Cell wall of plant (vegetables, fruits, and cereals) | Increase the content of fiber in food, thickening, gelling, and stabilizing | [29] | |
| Soluble/Insoluble dietary fiber | |||||
| Mixed plant cell wall fibers | Cellulose, hemicelluloses, and pectin | Fruits, vegetables, grains, legumes, pulses, nuts, and other plants | Increase the viscosity or gel strength | [21] | |
| Non-dietary fiber NDCs | |||||
| Resistant starch I | Physically embedded starch | Seeds or legumes and unprocessed whole grains | Ingredients for creating fibre-rich food, increase swelling, viscosity, and gel- forming capacity | [22] | |
| Resistant starch III | Regenerated starch | Starch-containing foods are cooked and cooled (corn starches, pasta, stale bread) | Improves texture, strength, and crispness in baked goods and extruded products such as cereals and snack foods | [22] | |
| Resistant starch IV | Chemically modified starch | Chemically modified starches food (breads and cakes) | Improve taste and texture, increase swelling, viscosity, and gel- forming capacity | [22] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chanmuang, S.; Nguyen, Q.-A.; Kim, H.-J. Current Research on the Effects of Non-Digestible Carbohydrates on Metabolic Disease. Appl. Sci. 2022, 12, 3768. https://doi.org/10.3390/app12083768
Chanmuang S, Nguyen Q-A, Kim H-J. Current Research on the Effects of Non-Digestible Carbohydrates on Metabolic Disease. Applied Sciences. 2022; 12(8):3768. https://doi.org/10.3390/app12083768
Chicago/Turabian StyleChanmuang, Saoraya, Quynh-An Nguyen, and Hyun-Jin Kim. 2022. "Current Research on the Effects of Non-Digestible Carbohydrates on Metabolic Disease" Applied Sciences 12, no. 8: 3768. https://doi.org/10.3390/app12083768
APA StyleChanmuang, S., Nguyen, Q.-A., & Kim, H.-J. (2022). Current Research on the Effects of Non-Digestible Carbohydrates on Metabolic Disease. Applied Sciences, 12(8), 3768. https://doi.org/10.3390/app12083768