
Biological Effect of Gamma Rays According to Exposure Time on Germination and Plant Growth in Wheat

 ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Gamma Irradiation
2.2. Germination Assays and Plant Growth
2.3. Chlorophyll Content Measurement
2.4. RNA Extraction and Gene Expression Analysis
2.5. Free Radical Content Measurement
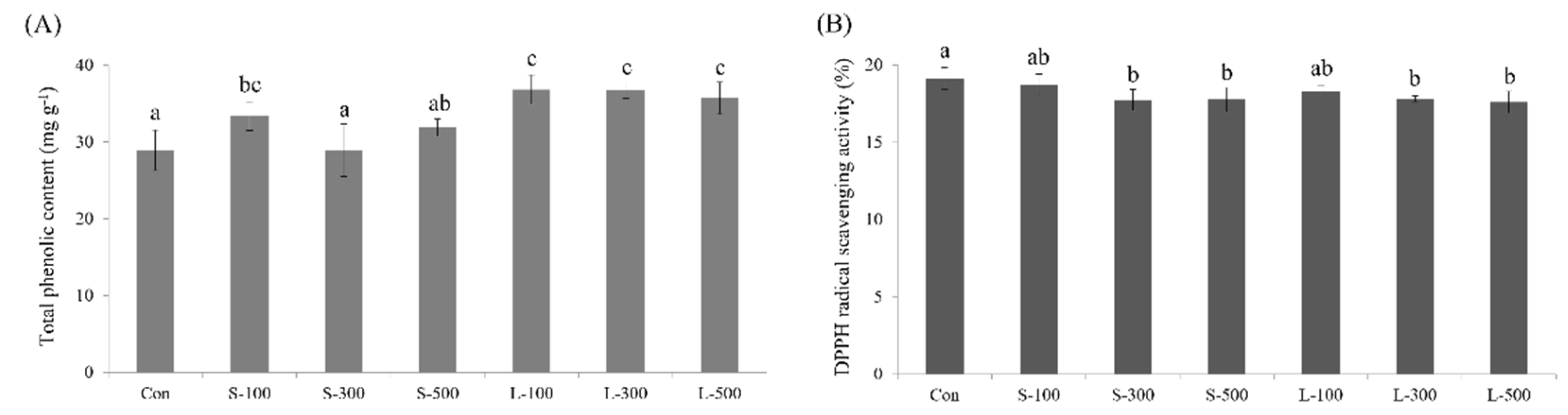
2.6. Total Phenolic Content
2.7. DPPH Radical Scavenging Activity
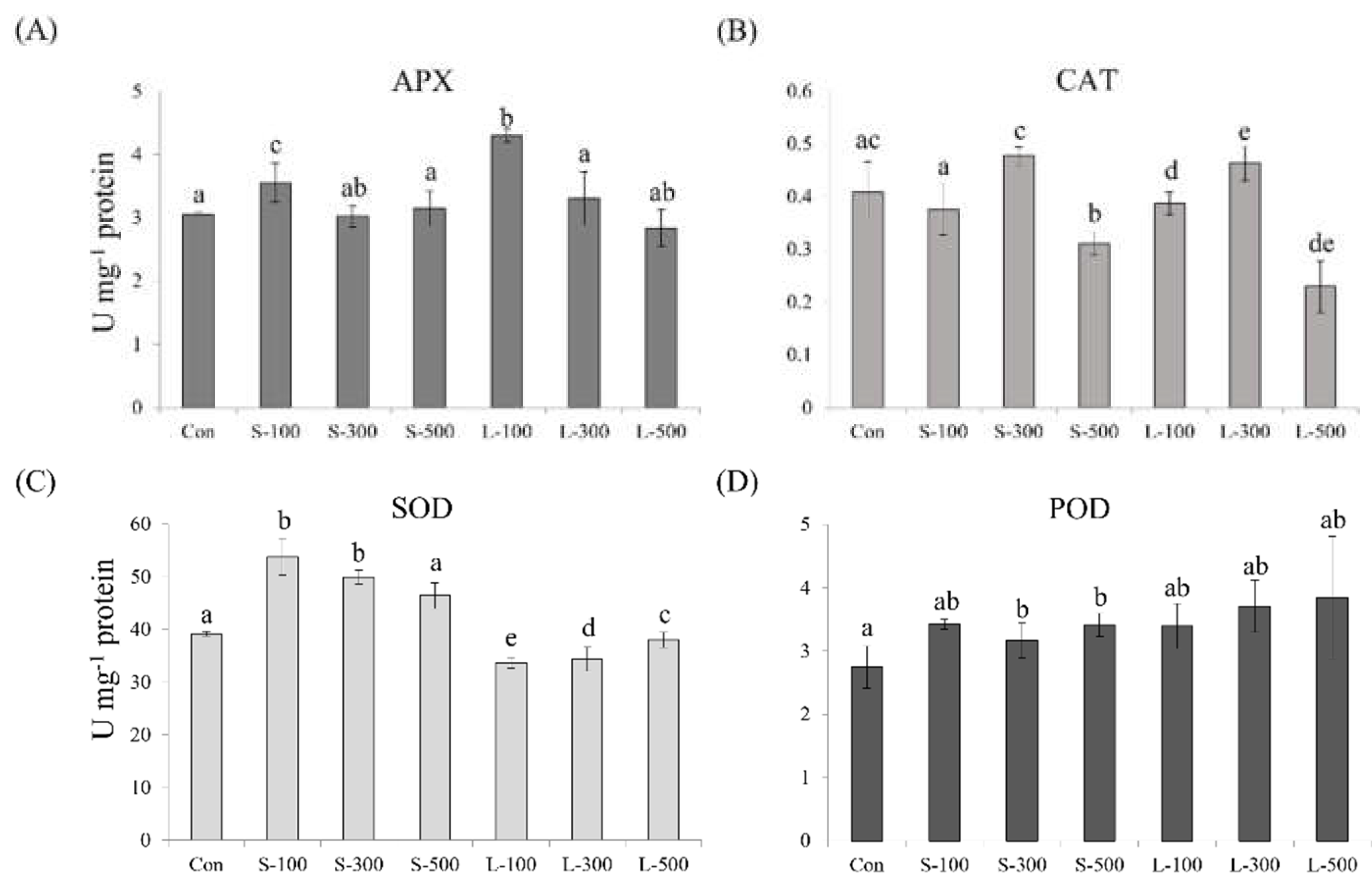
2.8. Antioxidant Enzyme Assay
2.9. Statistical Analysis
3. Results
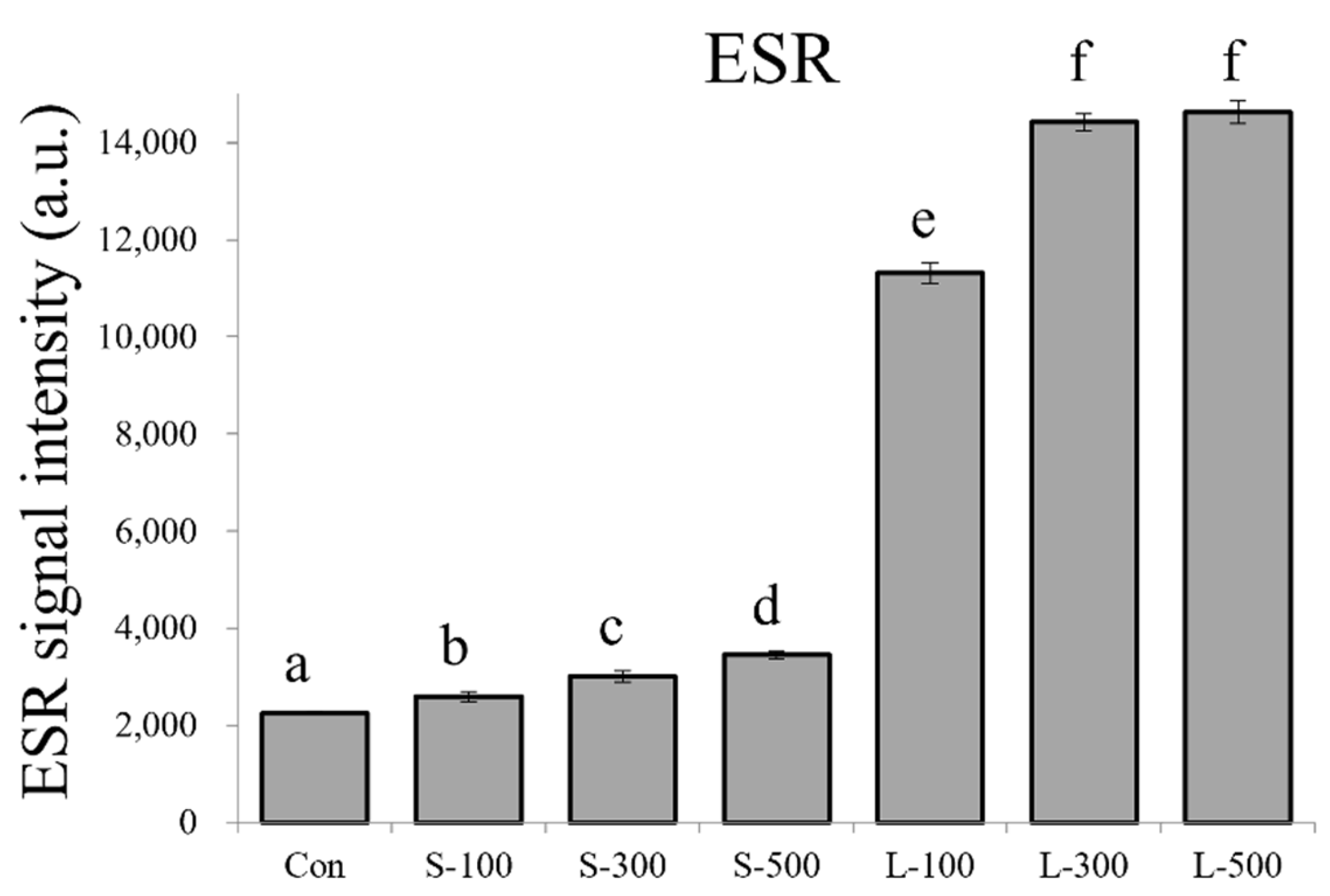
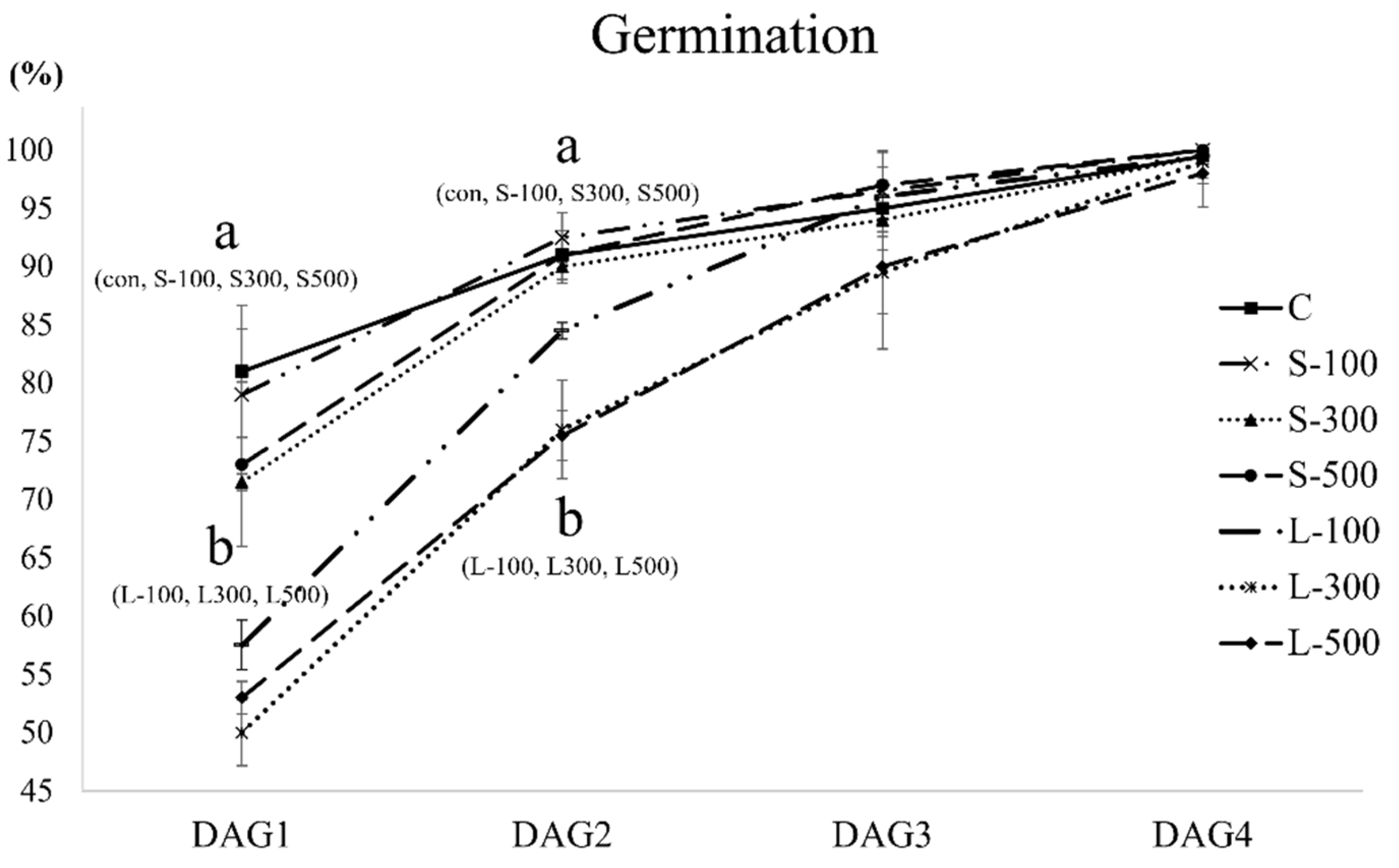
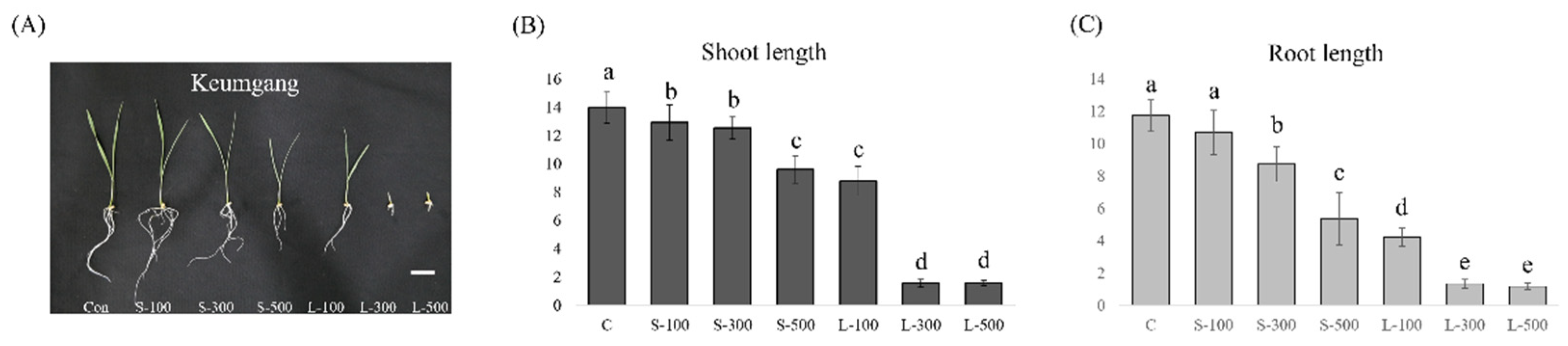
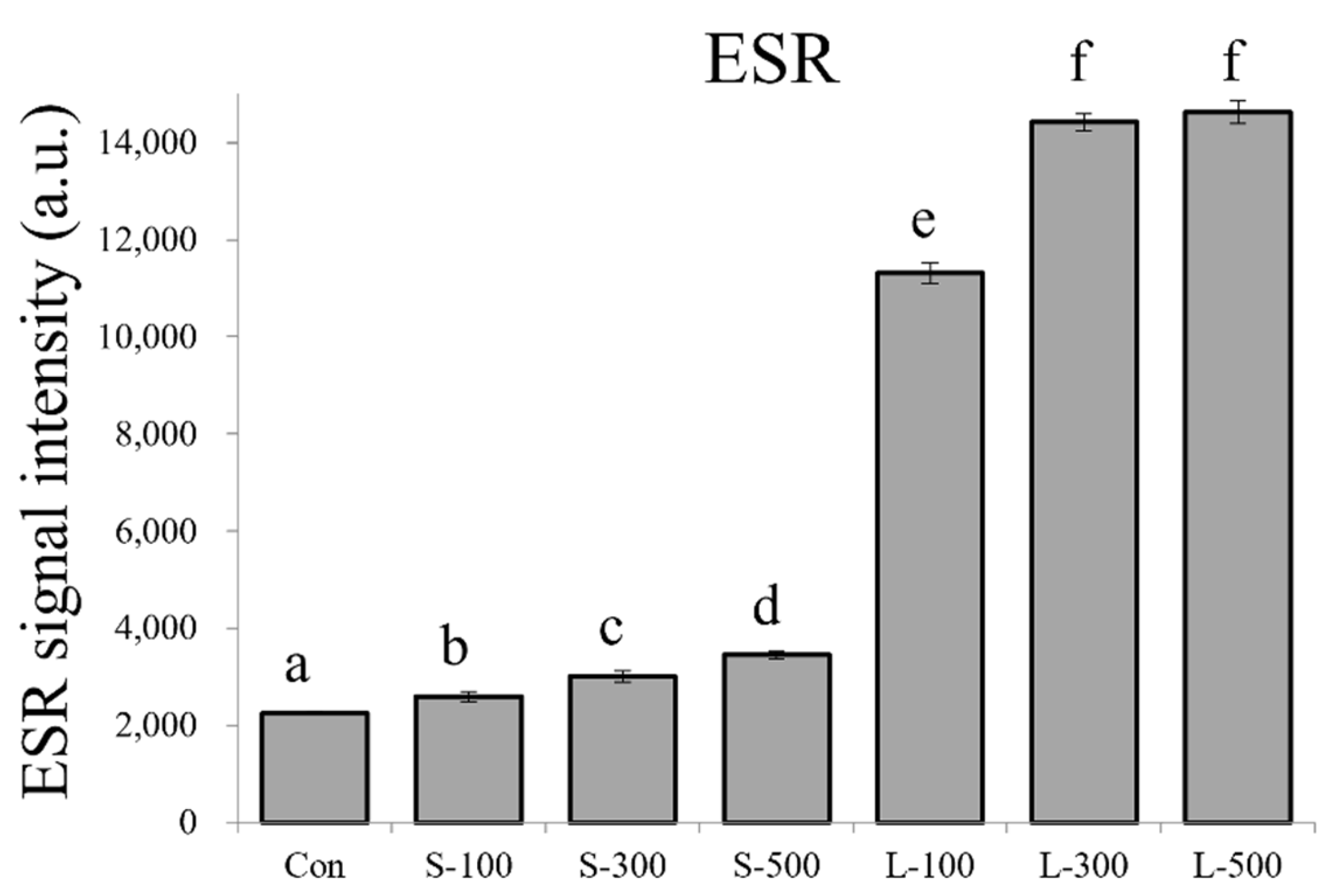
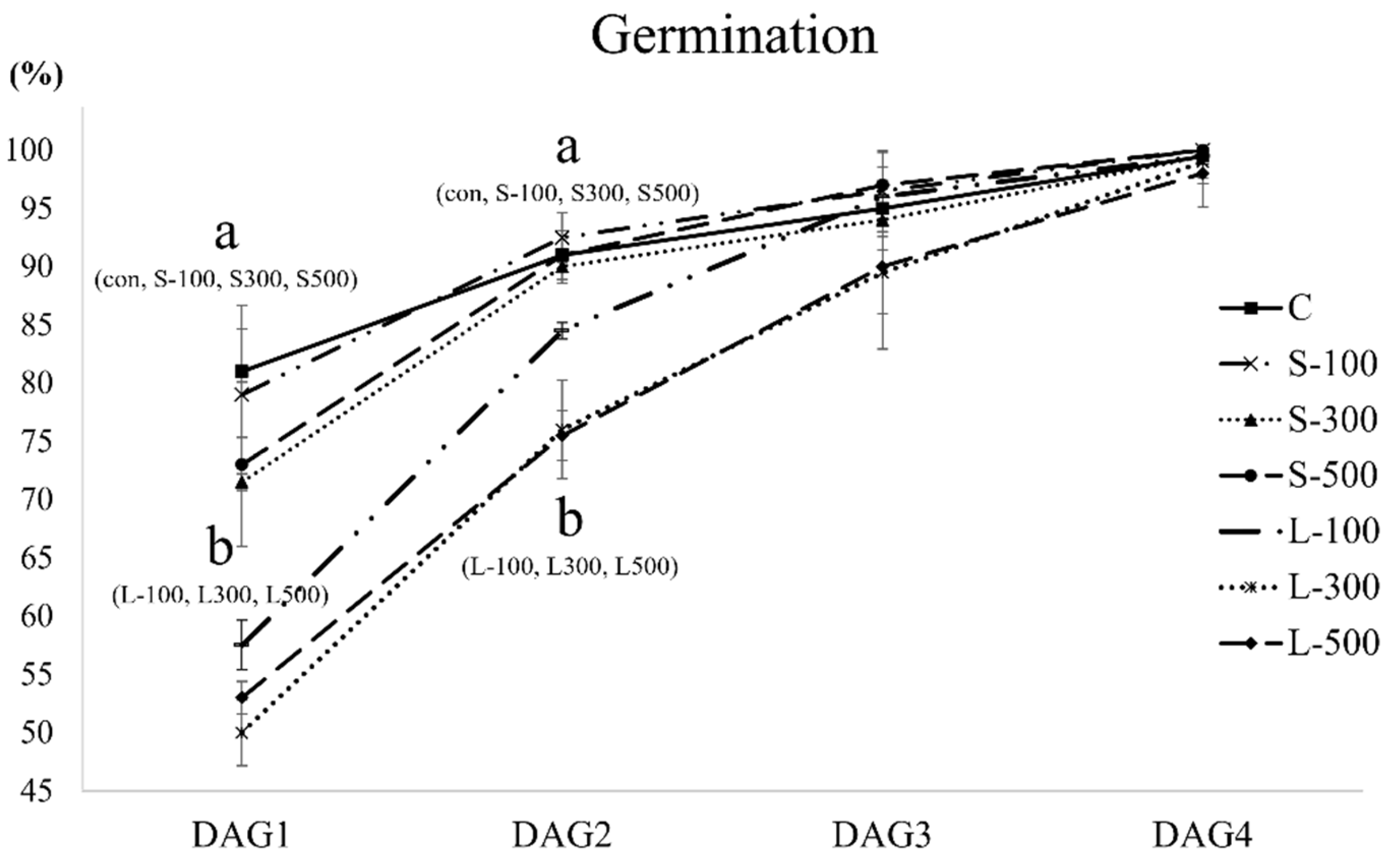
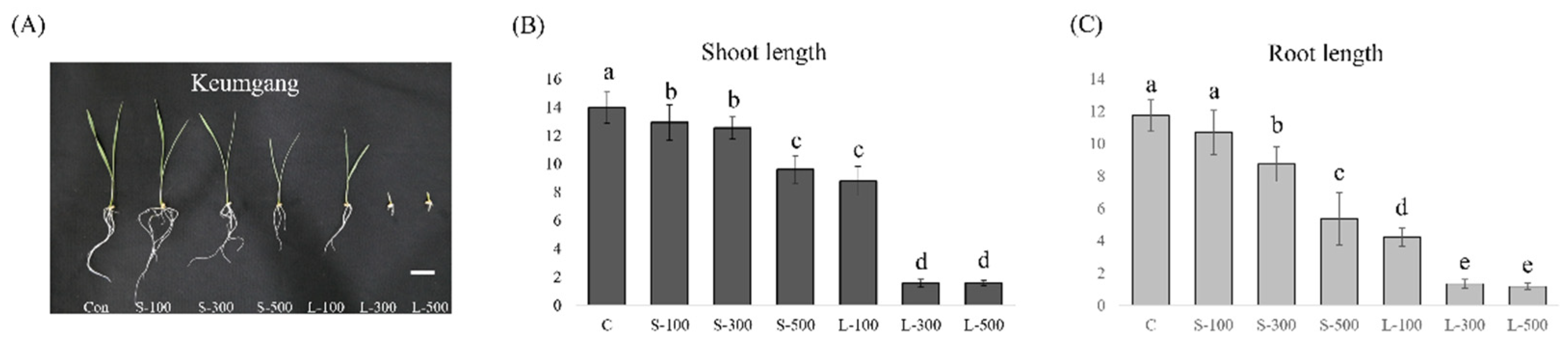
3.1. Free Radical Contents and Plant Growth
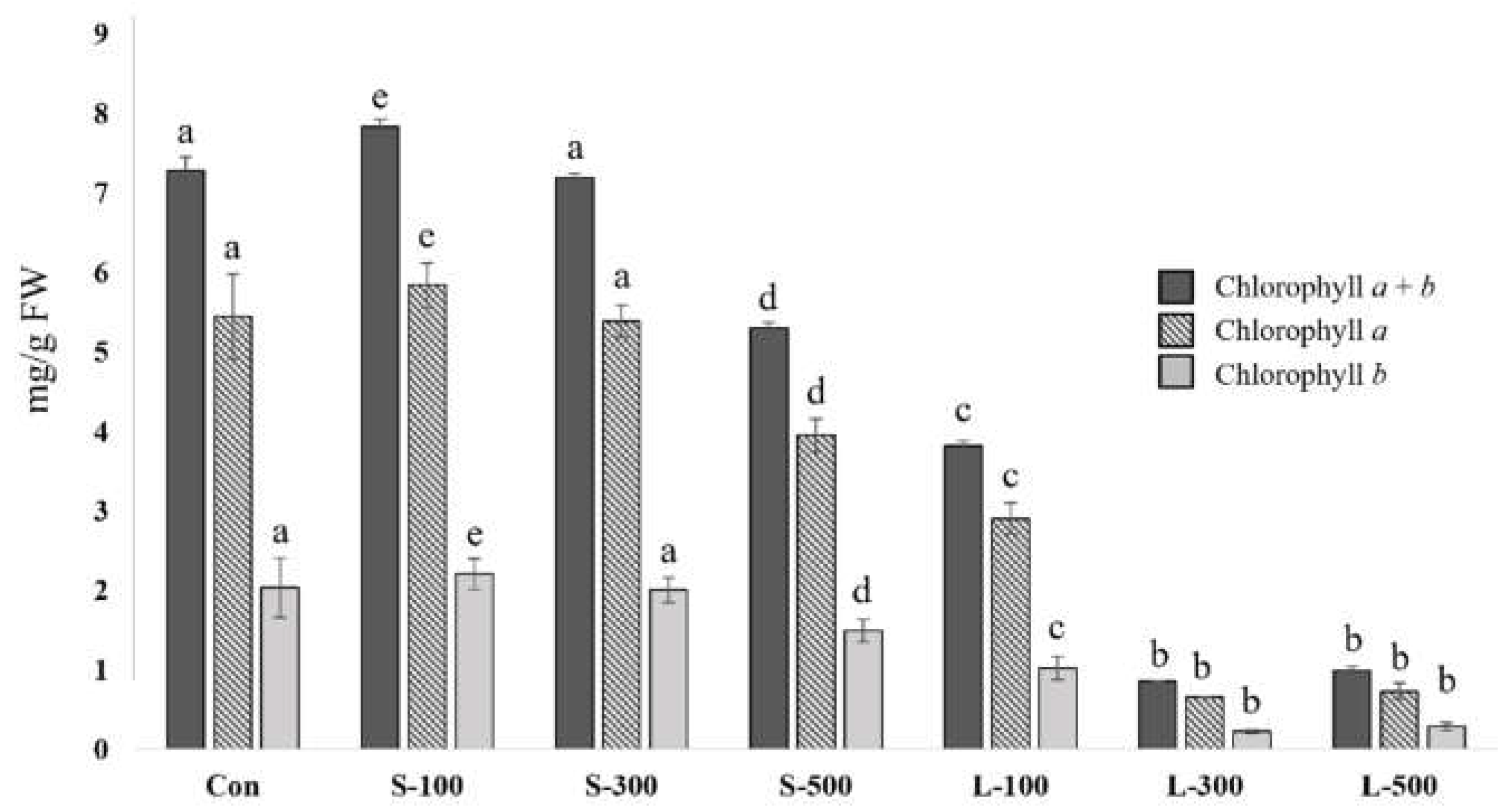
3.2. Chlorophyll Content Determination
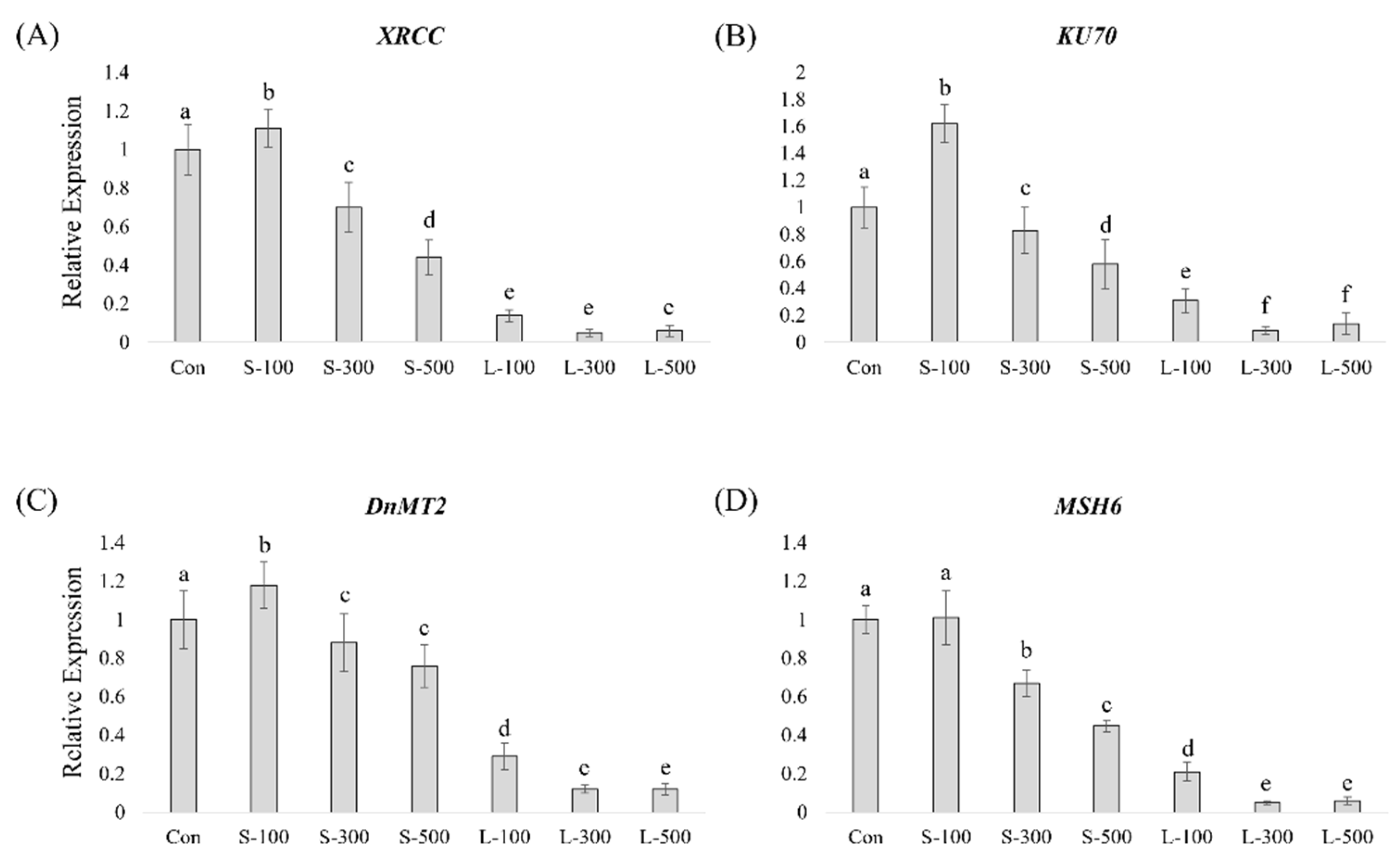
3.3. Gamma Radiation Effect on the Expression of DNA-Repair-Related Genes
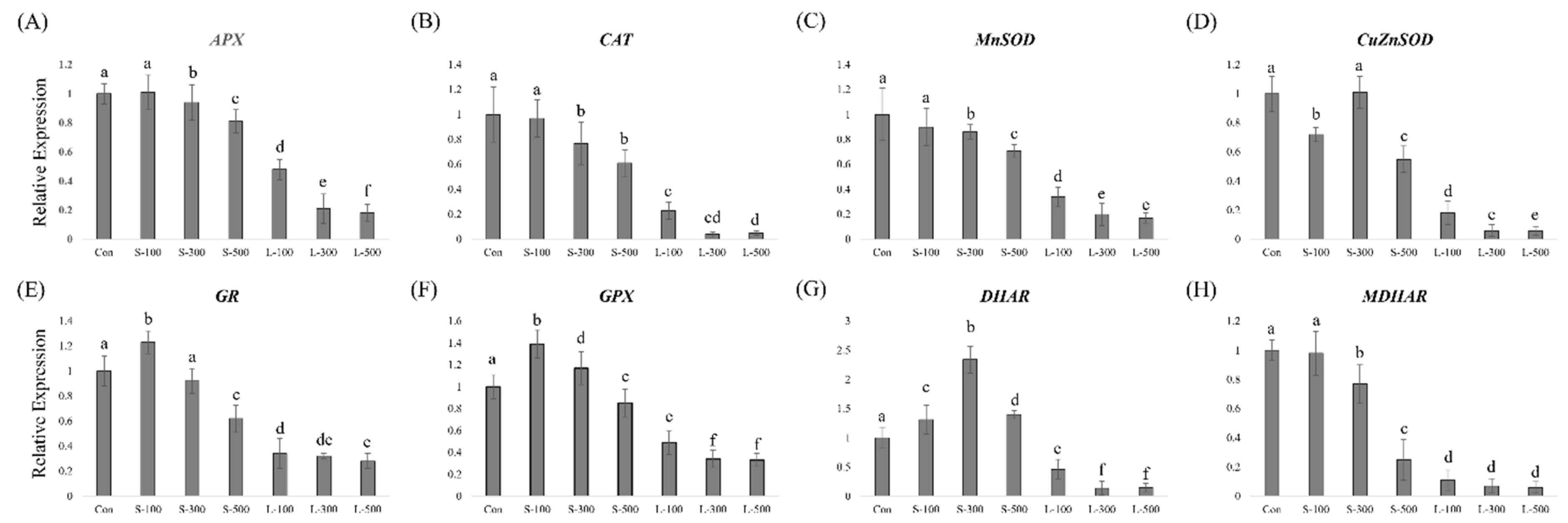
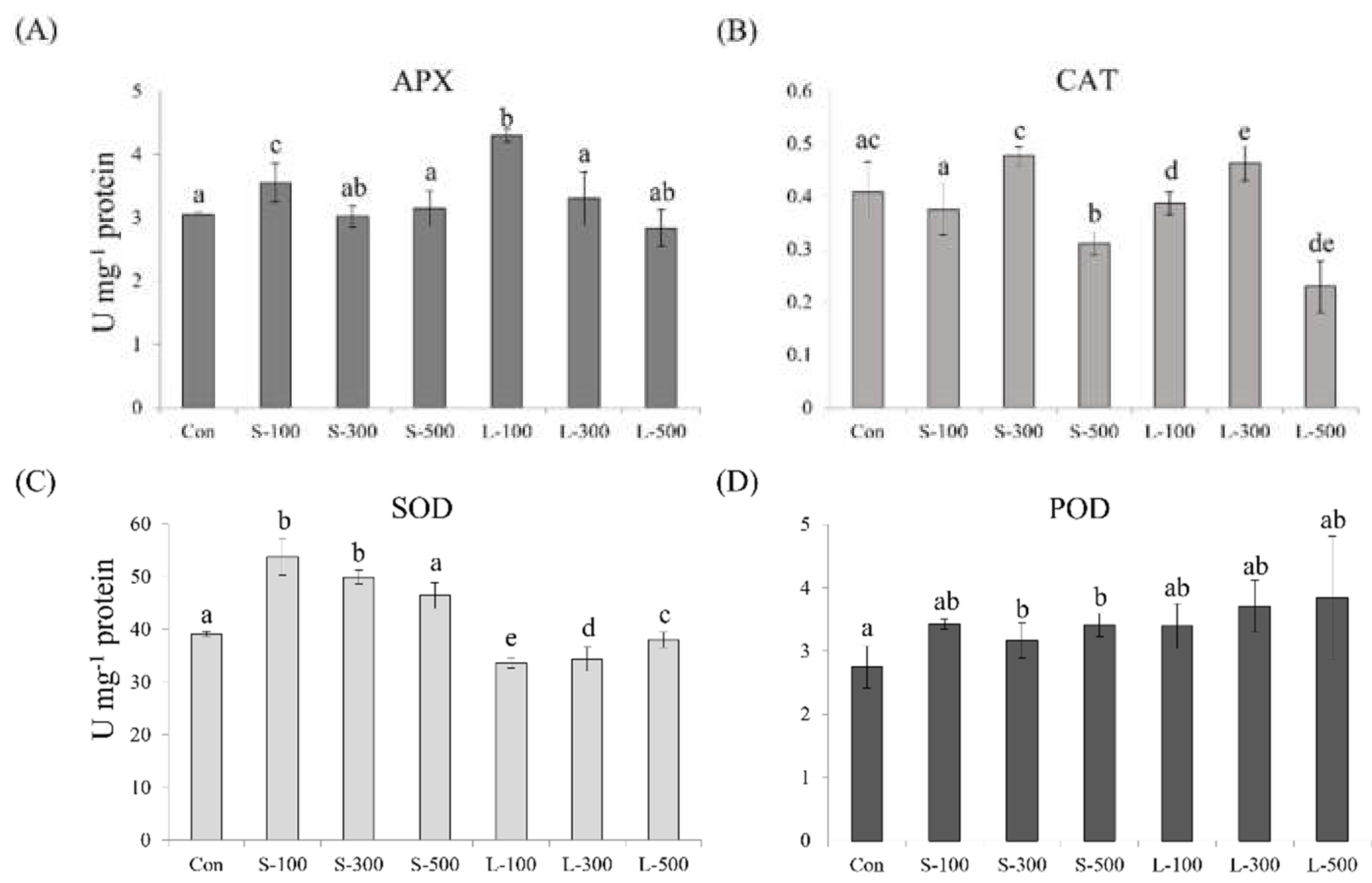
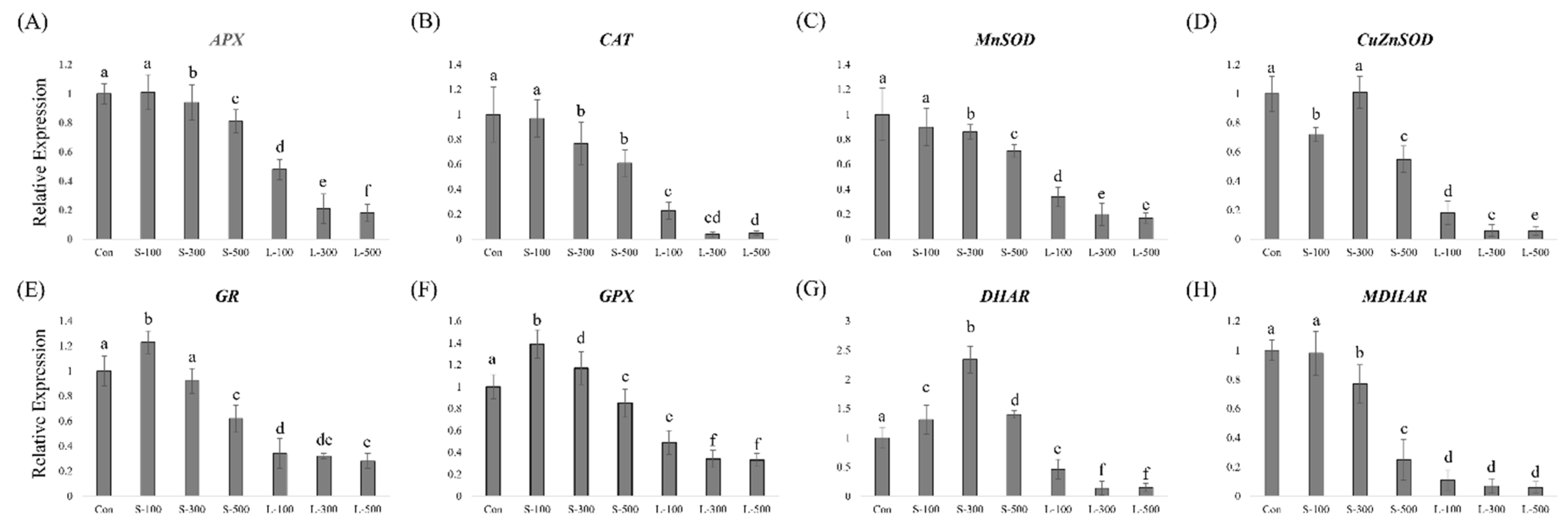
3.4. Antioxidant-Related Gene Expression Levels
3.5. Antioxidant Activity
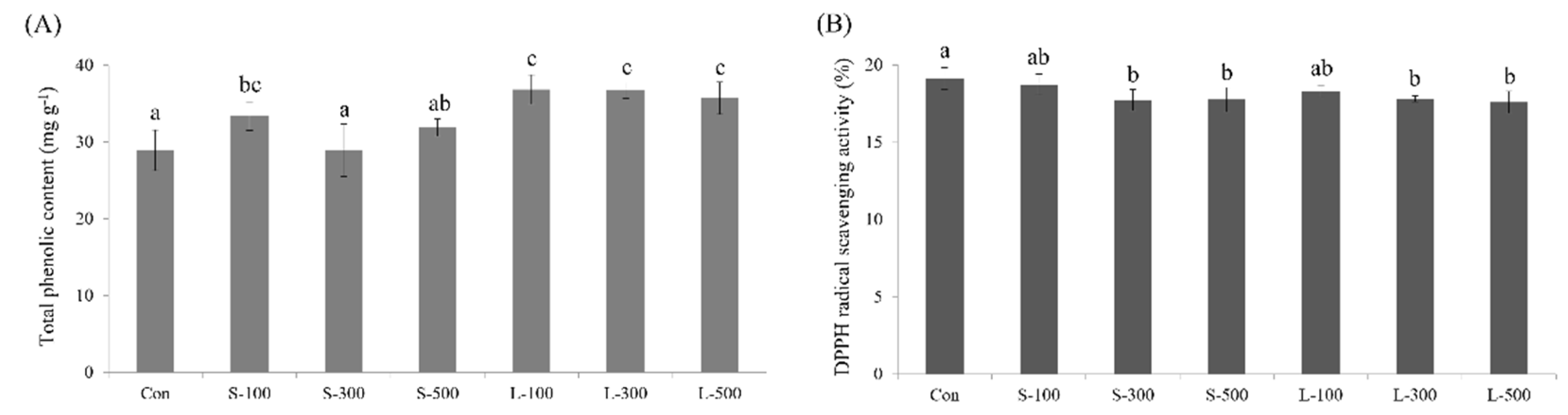
3.6. Total Phenolic Content and DPPH Free Radical Scavenging Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shiferaw, B.; Smale, M.; Braun, H.J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef]
- Venske, E.; Dos Santos, R.S.; Busanello, C.; Gustafson, P.; Costa de Oliveira, A. Bread wheat: A role model for plant domestication and breeding. Hereditas 2019, 156, 16. [Google Scholar] [CrossRef] [Green Version]
- Esnault, M.A.; Legue, F.; Chenal, C. Ionizing radiation: Advances in plant response. Environ. Exp. Bot. 2010, 68, 231–237. [Google Scholar] [CrossRef]
- Desouky, O.; Ding, N.; Zhou, G. Targeted and non-targeted effects of ionizing radiation. J. Radiat. Res. Appl. Sci. 2015, 8, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef]
- Wi, S.G.; Chung, B.Y.; Kim, J.S.; Kim, J.H.; Baek, M.H.; Lee, J.W.; Kim, Y.S. Effects of gamma irradiation on morphological changes and biological responses in plants. Micron 2007, 38, 553–564. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Ahmed, O.K.; El-Desouky, W. Effects of low doses γ-irradiation on oxidative stress and secondary metabolites production of rosemary (Rosmarinus officinalis L.) callus culture. Radiat. Phys. Chem. 2011, 80, 968–976. [Google Scholar] [CrossRef]
- Marcu, D.; Cristea, V.; Daraban, L. Dose-dependent effects of gamma radiation on lettuce (Lactuca sativa var. capitata) seedlings. Int. J. Radiat. Biol. 2013, 89, 219–223. [Google Scholar] [CrossRef]
- Li, F.; Shimizu, A.; Nishio, T.; Tsutsumi, N.; Kato, H. Comparison and characterization of mutations induced by gamma ray and carbon-ion irradiation in rice (Oryza sativa L.) using whole genome resequencing. G3 Genes Genomes Genet. 2019, 9, 3743–3751. [Google Scholar] [CrossRef] [Green Version]
- Han, A.R.; Hong, M.J.; Nam, B.; Kim, B.R.; Park, H.H.; Baek, I.; Kil, Y.S.; Nam, J.W.; Jin, C.H.; Kim, J.B. Comparison of flavonoid profiles in sprouts of radiation breeding wheat lines (Triticum aestivum L.). Agronomy 2020, 10, 1489. [Google Scholar] [CrossRef]
- Hong, M.J.; Kim, D.Y.; Nam, B.; Ahn, J.W.; Kwon, S.J.; Seo, Y.W.; Kim, J.B. Characterization of novel mutants of hexaploid wheat (Triticum aestivum L.) with various depths of purple grain color and antioxidant capacity. J. Sci. Food Agric. 2019, 99, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahimi, M.M.; Bahrani, A. Influence of gamma irradiation on some physiological characteristics and grain protein in wheat (Triticum aestivum L.). World Appl. Sci. J. 2011, 15, 654–659. [Google Scholar]
- Wu, J.H.; Zhang, J.; Lan, F.; Fan, W.F.; Li, W. Morphological, cytological, and molecular variations induced by gamma rays in ground-grown chrysanthemum ‘Pinkling’. Can. J. Plant Sci. 2019, 100, 68–77. [Google Scholar] [CrossRef]
- Pallavi, B.; Nivas, S.K.; D’Souza, L.; Ganapathi, T.R.; Hegde, S. Gamma rays induced variations in seed germination, growth and phenotypic characteristics of Zinnia elegans var. Dreamland. Adv. Hort. Sci. 2017, 31, 267–273. [Google Scholar]
- Andrew-Peter-Leon, M.T.; Ramchander, S.; Kumar, K.K.; Muthamilarasan, M.; Pillai, M.A. Assessment of efficacy of mutagenesis of gamma irradiation in plant height and days to maturity through expression analysis in rice. PLoS ONE 2021, 16, e0245603. [Google Scholar]
- Hase, Y.; Satoh, K.; Seito, H.; Oono, Y. Genetic consequences of acute/chronic gamma and carbon ion irradiation of Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 336. [Google Scholar] [CrossRef] [Green Version]
- Borzouei, A.; Kafi, M.; Khazaei, H.; Naseriyan, B.; Majdabadi, A. Effects of gamma radiation on germination and physiological aspects of wheat (Triticum aestivum L.) seedlings. Pak. J. Bot. 2010, 42, 2281–2290. [Google Scholar]
- Melki, M.; Marouani, A. Effects of gamma rays irradiation on seed germination and growth of hard wheat. Environ. Chem. Lett. 2010, 8, 307–310. [Google Scholar] [CrossRef]
- Pane, F.J.D.; Lopez, S.C.; Cantamutto, M.Á.; Domenech, M.B.; Castro-Franco, M. Effect of different gamma irradiation doses on the germination and seedling growth of wheat and triticale cultivars. Aust. J. Crop Sci. 2018, 12, 1921–1926. [Google Scholar] [CrossRef]
- ISTA (International Seed Testing Association). International rules for seed testing. Seed Sci. Technol. 1996, 24, 155–202. [Google Scholar]
- Esechie, H. Interaction of salinity and temperature on the germination of sorghum. J. Agron. Crop Sci. 1994, 172, 194–199. [Google Scholar] [CrossRef]
- Ellis, R.H.; Roberts, E.H. The quantification of ageing and survival in orthodox seeds. Seed Sci. Technol. 1981, 2, 373–409. [Google Scholar]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-vis spectroscopy. Curr. Protoc. Food Analyt. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophyll and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Coklar, H.; Akbulut, M. Anthocyanins and phenolic compounds of Mahonia aquifolium berries and their contributions to antioxidant activities. J. Funct. Foods 2017, 35, 166–174. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell. Physiol. 1981, 22, 867–880. [Google Scholar]
- Aebi, H.E. Catalasein Methods of Enzymatic Analysis, 3rd ed.; Bergmeyer, H.U., Ed.; Verlag Chemie: Weinheim, Germany, 1983; pp. 273–286. [Google Scholar]
- Kwak, S.S.; Kim, S.K.; Lee, M.S.; Jung, K.H.; Park, I.H.; Liu, J.R. Acidic peroxidases from suspension-cultures of sweet potato. Phytochemistry 1995, 39, 981–984. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, S.S.; Jaafar, H.; Ibrahim, R.; Rahmat, A.; Aziz, M.A.; Philip, E. Effects of acute gamma irradiation on physiological traits and flavonoid accumulation of Centella asiatica. Molecules 2011, 16, 4994–5007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcu, D.; Damian, G.; Cosma, C.; Cristea, V. Gamma radiation effects on seed germination, growth and pigment content, and ESR study of induced free radicals in maize (Zea mays). J. Biol. Phys. 2013, 39, 625–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jan, S.; Parween, T.; Siddiqi, T.O. Effects of gamma radiation on morphological, biochemical, and physiological aspects of plants and plant products. Environ. Rev. 2012, 20, 17–39. [Google Scholar] [CrossRef] [Green Version]
- Caplin, N.; Willey, N. Ionizing radiation, higher plants, and radioprotection: From acute high doses to chronic low doses. Front. Plant Sci. 2018, 9, 847. [Google Scholar] [CrossRef]
- Badr, A.; El-Shazly, H.H.; Halawa, M. Cytological effects of gamma radiation and its impact on growth and yield of M1 and M2 plants of cowpea cultivars. Cytologia 2014, 79, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Hakala, K.; Jauhiainen, L.; Koskela, T.; Käyhkö, P.; Vorne, V. Sensitivity of crops to increased ultraviolet radiation in northern growing conditions. J. Agron. Crop Sci. 2002, 188, 8–18. [Google Scholar] [CrossRef]
- Gudkov, I.N.; Grodzinsky, D.M. Cell radiosensitivity variation in synchronously-dividing root meristems of Pisum sativum L. and Zea mays L. during the mitotic cycle. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1982, 41, 401–409. [Google Scholar] [CrossRef]
- Hong, M.J.; Kim, J.B.; Yoon, Y.H.; Kim, S.H.; Ahn, J.W.; Jeong, I.Y.; Kang, S.Y.; Seo, Y.W.; Kim, D.S. The effects of chronic gamma irradiation on oxidative stress response and the expression of anthocyanin biosynthesis-related genes in wheat (Triticum aestivum). Int. J. Radiat. Biol. 2014, 90, 1218–1228. [Google Scholar] [CrossRef]
- Kim, D.Y.; Hong, M.J.; Park, C.S.; Seo, Y.W. The effects of chronic radiation of gamma ray on protein expression and oxidative stress in Brachypodium distachyon. Int. J. Radiat. Biol. 2015, 91, 407–419. [Google Scholar] [CrossRef]
- Irfaq, M.; Nawab, K. Effect of gamma irradiation on some morphological characteristics on three wheat (Triticum aestivum L.) cultivars. J. Biol. Sci. 2001, 1, 935–937. [Google Scholar]
- Preuss, S.B.; Britt, A.B. A DNA-damage-induced cell cycle checkpoint in Arabidopsis. Genetics 2003, 164, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Von Well, E.; Fossey, A.; Booyse, M. Efficiency of energy conversion and growth of gamma irradiated embryos and young seedlings of Triticum monococcum L. cultivar Einkorn. J. Radiat. Res. Appl. Sci. 2018, 11, 75–82. [Google Scholar] [CrossRef]
- Ananthaswamy, H.N.; Vakil, U.K.; Sreenivasan, A. Biochemical changes in gamma-irradiated wheat during germination. Radiat. Bot. 1971, 11, 1–12. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Han, B.; Gu, J.; Zhao, L.; Guo, H.; Xie, Y.; Zhao, S.; Song, X.; Han, L.; Liu, L. Factors affecting the radiosensitivity of hexaploid wheat to γ-irradiation: Radiosensitivity of hexaploidy wheat (Triticum aestivum L.). PLoS ONE 2016, 11, e0161700. [Google Scholar] [CrossRef]
- Kovalchuk, O.; Arkhipov, A.; Barylyak, I.; Karachov, I.; Titov, V.; Hohn, B.; Kovalchuk, I. Plants experiencing chronic internal exposure to ionizing radiation exhibit higher frequency of homologous recombination than acutely irradiated plants. Mutat. Res. 2000, 449, 47–56. [Google Scholar] [CrossRef]
- Vandenhove, H.; Vanhoudt, N.; Cuypers, A.; Van Hees, M.; Wannijn, J.; Horemans, N. Life-cycle chronic gamma exposure of Arabidopsis thaliana induces growth effects but no discernable effects on oxidative stress pathways. Plant. Physiol. Biochem. 2010, 48, 778–786. [Google Scholar] [CrossRef]
- Choi, H.I.; Han, S.M.; Jo, Y.D.; Hong, M.J.; Kim, S.H.; Kim, J.B. Effects of acute and chronic gamma irradiation on the cell biology and physiology of rice plants. Plants 2021, 10, 439. [Google Scholar] [CrossRef]
- Hong, M.J.; Kim, D.Y.; Ahn, J.W.; Kang, S.Y.; Seo, Y.W.; Kim, J.B. Comparison of radiosensitivity response to acute and chronic gamma irradiation in colored wheat. Genet. Mol. Biol. 2018, 41, 611–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Song, M.; Lee, K.J.; Hwang, S.G.; Jang, C.S.; Kim, J.B.; Kim, S.H.; Ha, B.K.; Kang, S.Y.; Kim, D.S. Genome-wide transcriptome profiling of ROS scavenging and signal transduction pathways in rice (Oryza sativa L.) in response to different types of ionizing radiation. Mol. Biol. Rep. 2012, 39, 11231–11248. [Google Scholar] [CrossRef] [PubMed]
- Ptácek, O.; Stavreva, D.A.; Kim, J.K.; Gichner, T. Induction and repair of DNA damage as measured by the comet assay and the yield of somatic mutations in gamma-irradiated tobacco seedlings. Mutat. Res. 2001, 491, 17–23. [Google Scholar] [CrossRef]
- Nishiguchi, M.; Nanjo, T.; Yoshida, K. The effects of gamma irradiation on growth and expression of genes encoding DNA repair-related proteins in Lombardy poplar (Populus nigra var. italica). J. Environ. Radioact. 2012, 109, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Mannuss, A.; Trapp, O.; Puchta, H. Gene regulation in response to DNA damage. Biochim. Biophys. Acta 2012, 1819, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Nazarenko, M.; Lykholat, Y.; Grygoryuk, I.; Khromikh, N. Optimal doses and concentrations of mutagens for winter wheat breeding purposes. Part I. Grain productivity. J. Cent. Eur. Agric. 2018, 19, 194–205. [Google Scholar] [CrossRef]
- Dwinanda, P.; Syukur, S.; Suliansyah, I. Induction of mutations with gamma ray radiation to improve the characteristics of wheat (Triticum aestivum L.) genotype IS-Jarissa. IOP Conf. Ser. Earth Environ. Sci. 2020, 497, 012013. [Google Scholar] [CrossRef]
- Sen, Y.; Ozturk, I.; Yaycili, O.; Alikamanoglu, S. Drought tolerance in irradiated wheat mutants studied by genetic and biochemical marker. J. Plant Growth Regul. 2017, 36, 669–679. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Germination Rate Index (%) | Mean Germination Time (Days) | |
|---|---|---|
| C | 88.54 ± 1.97 | 1.295 ± 0.017 |
| S-100 | 87.96 ± 3.42 | 1.320 ± 0.000 |
| S-300 | 83.46 ± 0.14 | 1.425 ± 0.040 |
| S-500 | 84.75 ± 4.23 | 1.390 ± 0.023 |
| L-100 | 75.71 ± 0.91 | 1.600 ± 0.058 |
| L-300 | 69.88 ± 2.26 | 1.805 ± 0.040 |
| L-500 | 71.08 ± 0.77 | 1.735 ± 0.029 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, M.J.; Kim, D.Y.; Jo, Y.D.; Choi, H.-I.; Ahn, J.-W.; Kwon, S.-J.; Kim, S.H.; Seo, Y.W.; Kim, J.-B. Biological Effect of Gamma Rays According to Exposure Time on Germination and Plant Growth in Wheat. Appl. Sci. 2022, 12, 3208. https://doi.org/10.3390/app12063208
Hong MJ, Kim DY, Jo YD, Choi H-I, Ahn J-W, Kwon S-J, Kim SH, Seo YW, Kim J-B. Biological Effect of Gamma Rays According to Exposure Time on Germination and Plant Growth in Wheat. Applied Sciences. 2022; 12(6):3208. https://doi.org/10.3390/app12063208
Chicago/Turabian StyleHong, Min Jeong, Dae Yeon Kim, Yeong Deuk Jo, Hong-Il Choi, Joon-Woo Ahn, Soon-Jae Kwon, Sang Hoon Kim, Yong Weon Seo, and Jin-Baek Kim. 2022. "Biological Effect of Gamma Rays According to Exposure Time on Germination and Plant Growth in Wheat" Applied Sciences 12, no. 6: 3208. https://doi.org/10.3390/app12063208
APA StyleHong, M. J., Kim, D. Y., Jo, Y. D., Choi, H.-I., Ahn, J.-W., Kwon, S.-J., Kim, S. H., Seo, Y. W., & Kim, J.-B. (2022). Biological Effect of Gamma Rays According to Exposure Time on Germination and Plant Growth in Wheat. Applied Sciences, 12(6), 3208. https://doi.org/10.3390/app12063208