
Hair Regeneration Effects of Lespedeza bicolor Extract In Vivo and In Vitro

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Cells
2.3. Cell Viability and Proliferation Assay
2.4. Western Blot Analysis
2.5. In Vivo Experiments
2.6. HPLC Analysis
2.7. Cell Cycle Analysis
2.8. Quantitative RT-PCR
2.9. Statistical Analysis
3. Results
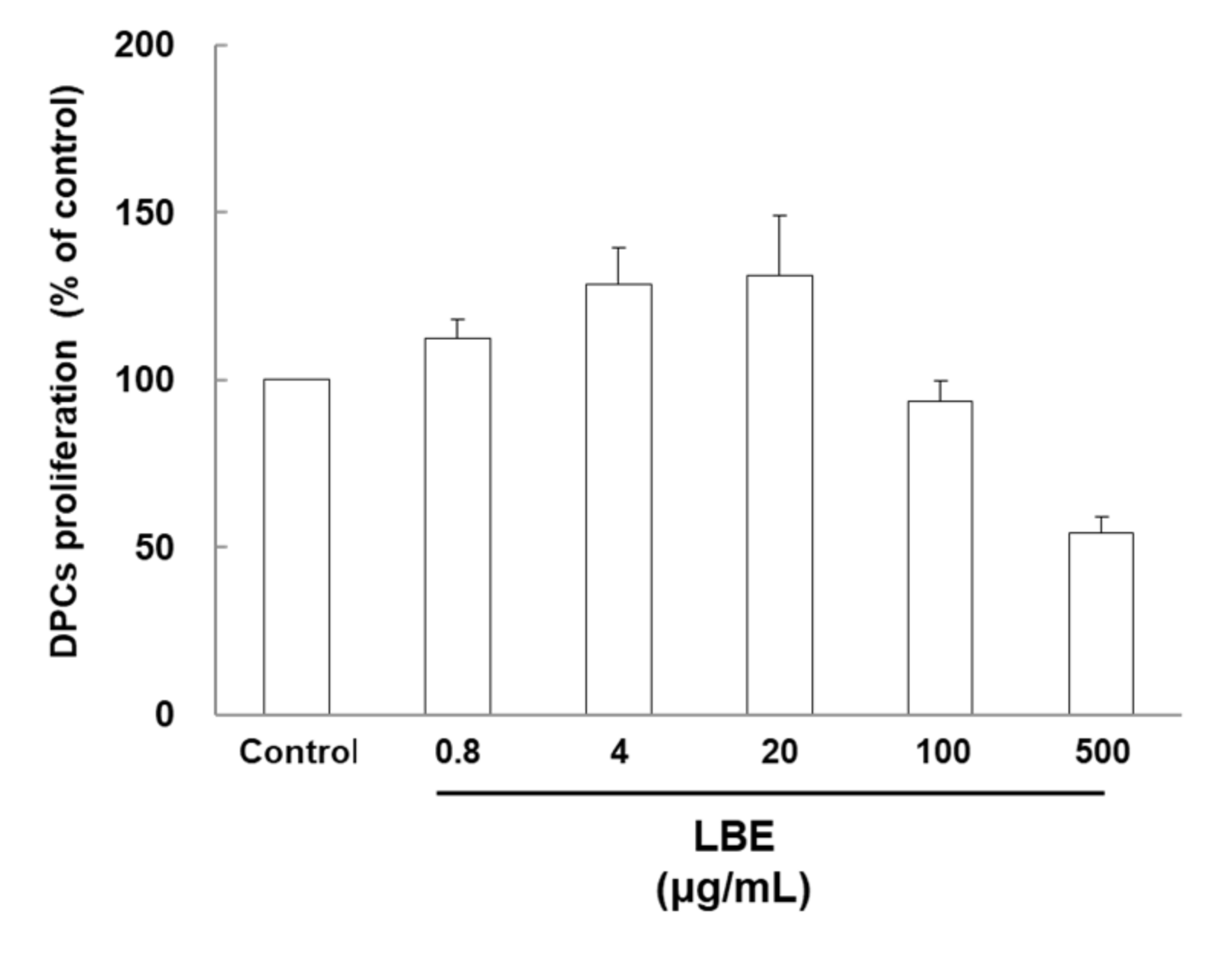
3.1. Effects of LBE on Human Hair Dermal Cell Proliferation
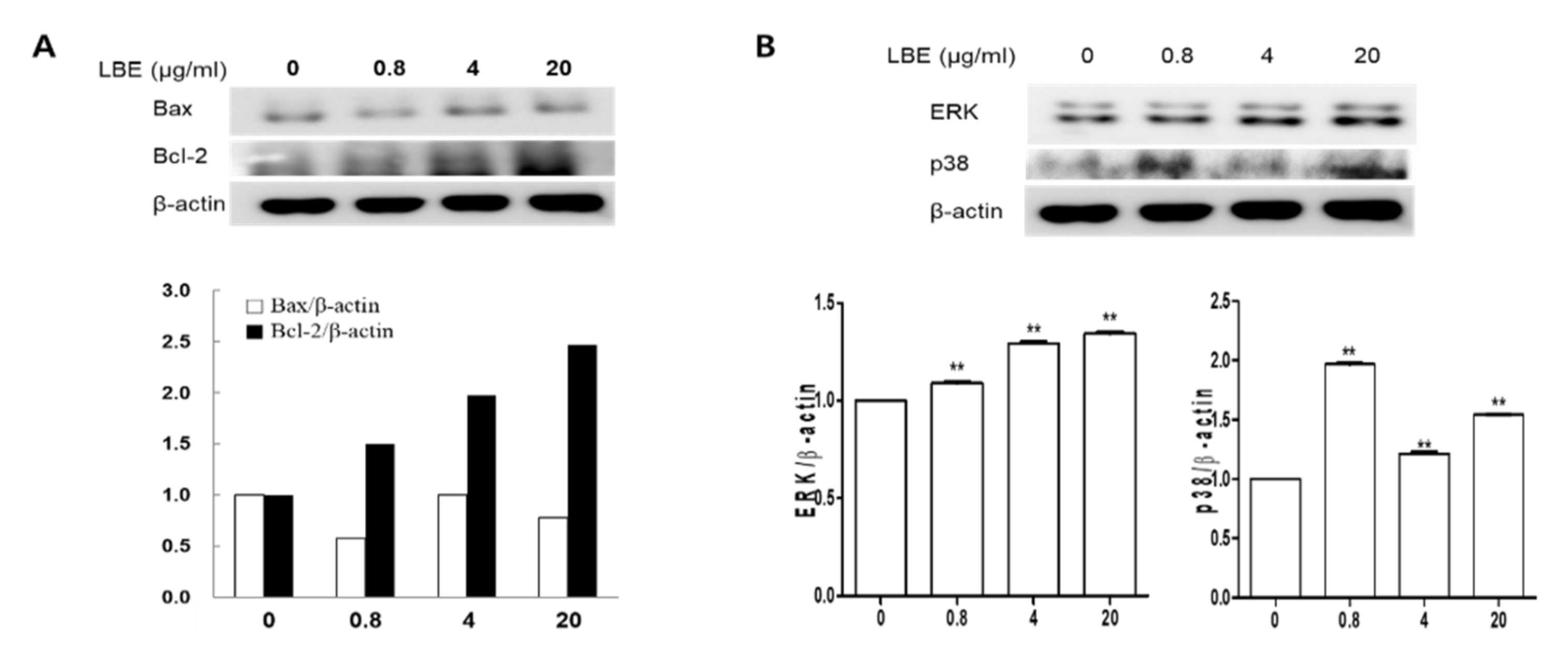
3.2. Effects of LBE on Protein Expressions in DPCs
3.3. Effects of LBE on Hair Regeneration from In Vivo
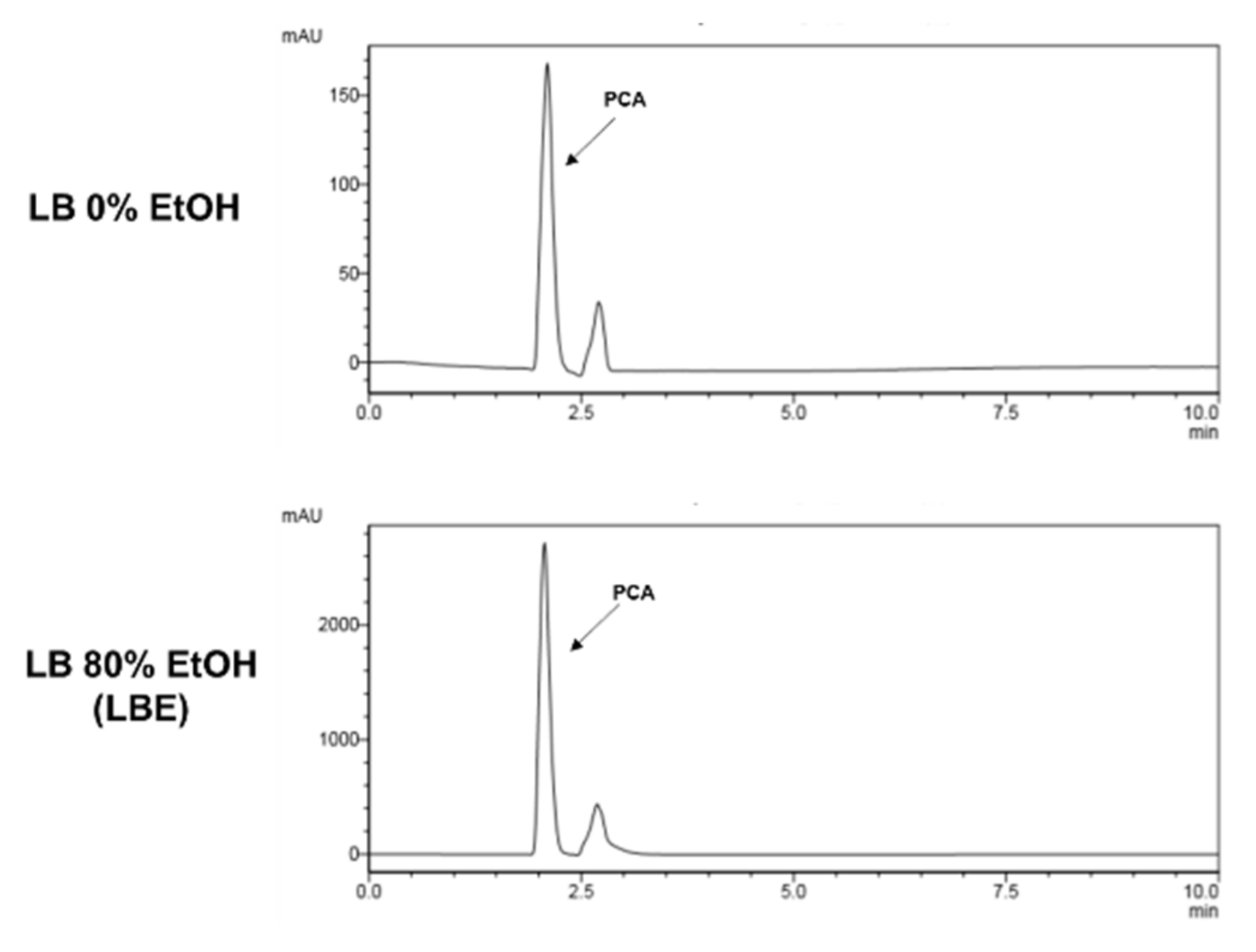
3.4. HPLC Analysis
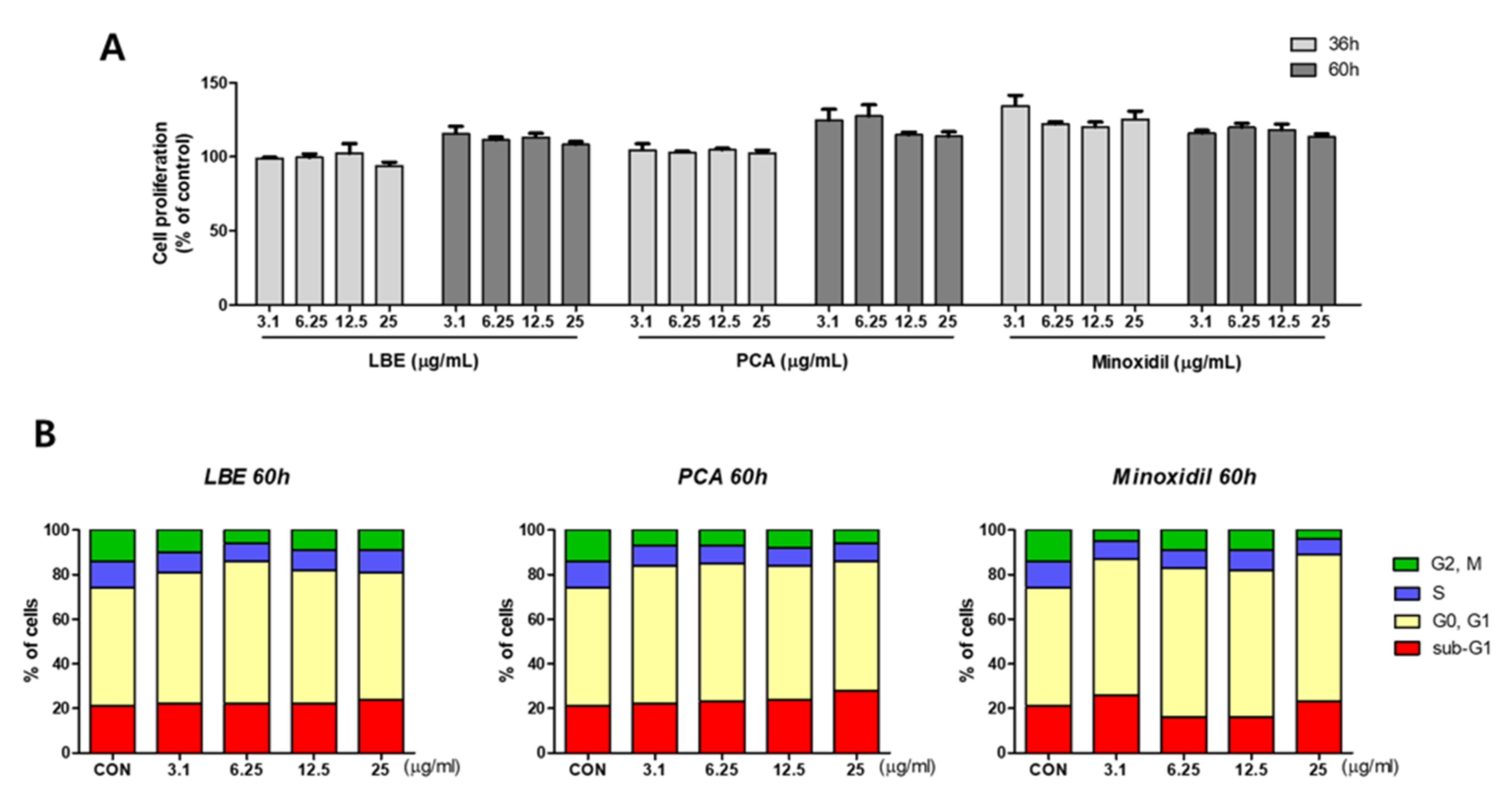
3.5. Effects of LBE on Cell Cycle Phase in DPCs
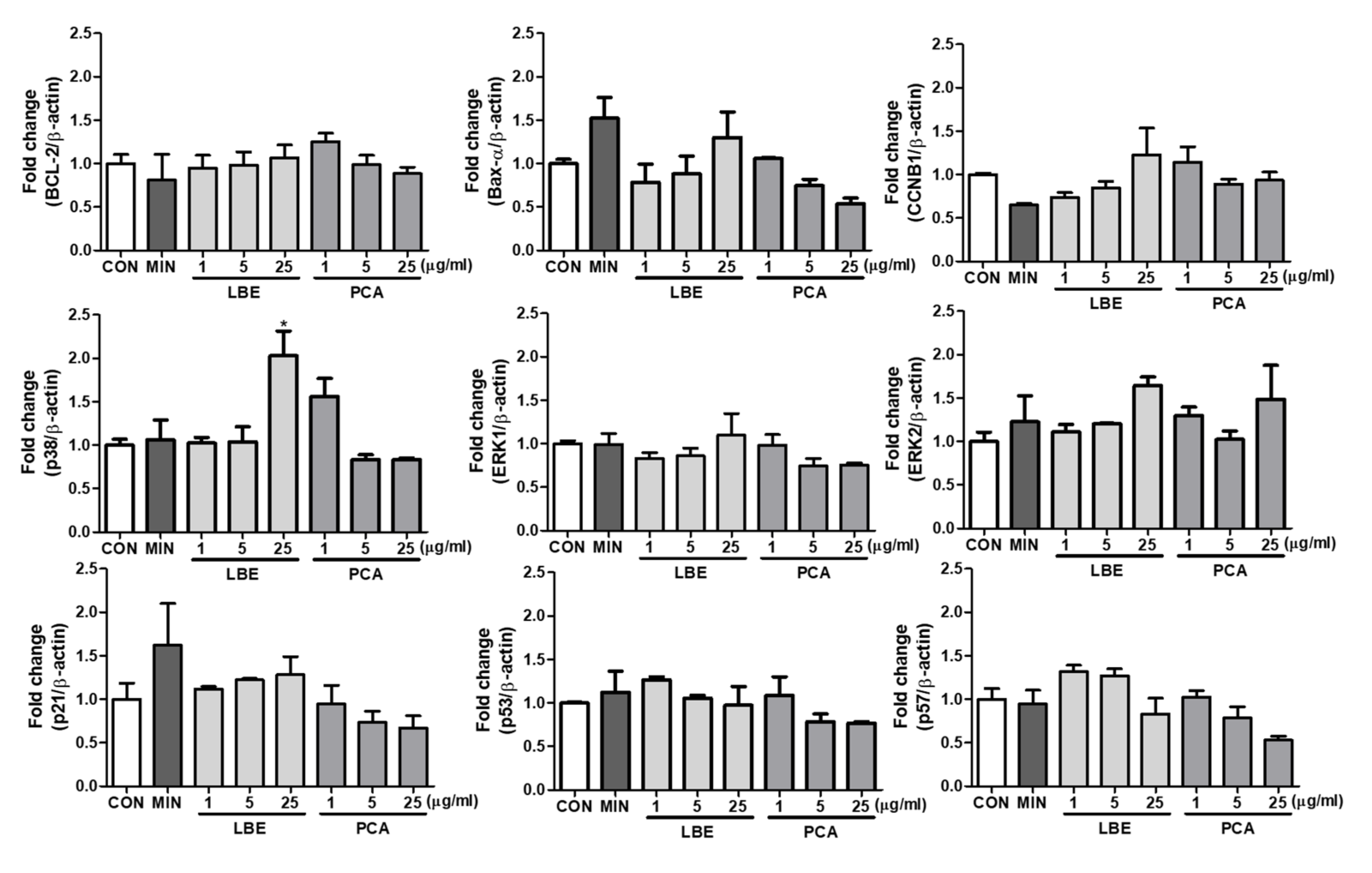
3.6. Effects of LBE on Cell Cycle Related Biomarkers in DPCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Greco, V.; Chen, T.; Rendl, M.; Schober, M.; Pasolli, H.A.; Stokes, N.; Dela Cruz-Racelis, J.; Fuchs, E. A two-step mechanism for stem cell activation during hair regeneration. Cell Stem Cell 2009, 4, 155–169. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Tredget, E.E.; Wu, Y. Dynamic signals for hair follicle development and regeneration. Stem Cells Dev. 2012, 21, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Myung, P.; Ito, M. Dissecting the bulge in hair regeneration. J. Clin. Investig. 2012, 122, 448–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Nuaimi, Y.; Baier, G.; Watson, R.E.; Chuong, C.M.; Paus, R. The cycling hair follicle as an ideal systems biology research model. Exp. Dermatol. 2010, 19, 707–713. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.C.; Cotsarelis, G. Review of hair follicle dermal cells. J. Dermatol. Sci 2010, 57, 2–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, K.; Stephenson, T.J.; Messenger, A.G. Differences in hair follicle dermal papilla volume are due to extracellular matrix volume and cell number: Implications for the control of hair follicle size and androgen responses. J. Investig. Dermatol. 1999, 113, 873–877. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J.A.; Sinclair, R.; Harrap, S.B. Androgenetic alopecia: Pathogenesis and potential for therapy. Expert Rev. Mol. Med. 2002, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hunt, N.; McHale, S. The psychological impact of alopecia. BMJ 2005, 331, 951–953. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, R.; Patel, M.; Dawson, T.L., Jr.; Yazdabadi, A.; Yip, L.; Perez, A.; Rufaut, N.W. Hair loss in women: Medical and cosmetic approaches to increase scalp hair fullness. Br. J. Dermatol. 2011, 165 (Suppl. 3), 12–18. [Google Scholar] [CrossRef]
- Arca, E.; Acikgoz, G.; Tastan, H.B.; Kose, O.; Kurumlu, Z. An open, randomized, comparative study of oral finasteride and 5% topical minoxidil in male androgenetic alopecia. Dermatology 2004, 209, 117–125. [Google Scholar] [CrossRef]
- McClellan, K.J.; Markham, A. Finasteride: A review of its use in male pattern hair loss. Drugs 1999, 57, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Trueb, R.M.; Itin, P.; Itin und Schweizerische Arbeitsgruppe für Trichologie. Photographic documentation of the effectiveness of 1 mg. oral finasteride in treatment of androgenic alopecia in the man in routine general practice in Switzerland. Praxis 2001, 90, 2087–2093. [Google Scholar] [PubMed]
- Paus, R.; Haslam, I.S.; Sharov, A.A.; Botchkarev, V.A. Pathobiology of chemotherapy-induced hair loss. Lancet Oncol. 2013, 14, e50–e59. [Google Scholar] [CrossRef]
- Lee, S.J.; Hossaine, M.D.; Park, S.C. A potential anti-inflammation activity and depigmentation effect of Lespedeza bicolor extract and its fractions. Saudi J. Biol. Sci. 2016, 23, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Maximov, O.B.; Kulesh, N.I.; Stepanenko, L.S.; Dmitrenok, P.S. New prenylated isoflavanones and other constituents of Lespedeza bicolor. Fitoterapia 2004, 75, 96–98. [Google Scholar] [CrossRef]
- Woo, H.S.; Kim, D.W.; Curtis-Long, M.J.; Lee, B.W.; Lee, J.H.; Kim, J.Y.; Kang, J.E.; Park, K.H. Potent inhibition of bacterial neuraminidase activity by pterocarpans isolated from the roots of Lespedeza bicolor. Bioorg. Med. Chem. Lett. 2011, 21, 6100–6103. [Google Scholar] [CrossRef]
- Do, M.H.; Lee, J.H.; Wahedi, H.M.; Pak, C.; Lee, C.H.; Yeo, E.J.; Lim, Y.; Ha, S.K.; Choi, I.; Kim, S.Y. Lespedeza bicolor ameliorates endothelial dysfunction induced by methylglyoxal glucotoxicity. Phytomedicine 2017, 36, 26–36. [Google Scholar] [CrossRef]
- Ko, Y.H.; Shim, K.Y.; Kim, S.K.; Seo, J.Y.; Lee, B.R.; Hur, K.H.; Kim, Y.J.; Kim, S.E.; Do, M.H.; Parveen, A.; et al. Lespedeza bicolor Extract Improves Amyloid Beta25—35-Induced Memory Impairments by Upregulating BDNF and Activating Akt, ERK, and CREB Signaling in Mice. Planta Med. 2019, 85, 1363–1373. [Google Scholar] [CrossRef]
- Park, J.E.; Lee, H.; Kim, S.Y.; Lim, Y. Lespedeza bicolor Extract Ameliorated Renal Inflammation by Regulation of NLRP3 Inflammasome-Associated Hyperinflammation in Type 2 Diabetic Mice. Antioxidants 2020, 9, 148. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Tan, J.; Zou, B.; Liu, X.; Yu, Y. Exploring the potential effect and mechanisms of protocatechuic acid on human hair follicle melanocytes. Acta Pharm. 2020, 70, 539–549. [Google Scholar] [CrossRef]
- Krzysztoforska, K.; Mirowska-Guzel, D.; Widy-Tyszkiewicz, E. Pharmacological effects of protocatechuic acid and its therapeutic potential in neurodegenerative diseases: Review on the basis of in vitro and in vivo studies in rodents and humans. Nutr. Neurosci. 2019, 22, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Hornedo-Ortega, R.; Alvarez-Fernandez, M.A.; Cerezo, A.B.; Richard, T.; Troncoso, A.M.A.; Garcia-Parrilla, M.A.C. Protocatechuic Acid: Inhibition of Fibril Formation, Destabilization of Preformed Fibrils of Amyloid-beta and alpha-Synuclein, and Neuroprotection. J. Agric. Food Chem. 2016, 64, 7722–7732. [Google Scholar] [CrossRef] [PubMed]
- Chase, H.B. Growth of the hair. Physiol. Rev. 1954, 34, 113–126. [Google Scholar] [CrossRef] [PubMed]
- El-Domyati, M.; Attia, S.; Saleh, F.; Bassyouni, M.I.; El-Fakahany, H.; Abdel-Wahab, H. Proliferation, DNA repair and apoptosis in androgenetic alopecia. J. Eur. Acad. Dermatol. Venereol. 2009, 23, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Boyera, N.; Galey, I.; Bernard, B.A. Biphasic effects of minoxidil on the proliferation and differentiation of normal human keratinocytes. Skin Pharmacol. 1997, 10, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Han, J.H.; Kwon, O.S.; Chung, J.H.; Cho, K.H.; Eun, H.C.; Kim, K.H. Effect of minoxidil on proliferation and apoptosis in dermal papilla cells of human hair follicle. J. Dermatol. Sci. 2004, 34, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Jung, H.A.; Choi, J.S.; Kim, Y.S.; Kang, S.S.; Kim, H.P. Anti-inflammatory activity of the constituents of the roots of Aralia continentalis. Arch. Pharm. Res. 2009, 32, 1237–1243. [Google Scholar] [CrossRef]
- Muller-Rover, S.; Handjiski, B.; van der Veen, C.; Eichmuller, S.; Foitzik, K.; McKay, I.A.; Stenn, K.S.; Paus, R. A comprehensive guide for the accurate classification of murine hair follicles in distinct hair cycle stages. J. Investig. Dermatol. 2001, 117, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Hendrix, S.; Handjiski, B.; Peters, E.M.; Paus, R. A guide to assessing damage response pathways of the hair follicle: Lessons from cyclophosphamide-induced alopecia in mice. J. Investig. Dermatol. 2005, 125, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Shin, W.S.; Ho, J. Fructus panax ginseng extract promotes hair regeneration in C57BL/6 mice. J. Ethnopharmacol. 2011, 138, 340–344. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Sequences | Accession No. | |
|---|---|---|---|
| Bax-α | Forward | TGATGGACGGGTCCGGG | NM_138763.4 |
| Reverse | CAAAGGGCCCCTGTCTTCA | ||
| Bcl-2 | Forward | CGGAGGCTGGGATGCCTTTGT | NM_000633.3 |
| Reverse | CAAGCTCCCACCAGGGCCAAA | ||
| ERK1 | Forward | ACTCCAAAGCCCTTGACCTG | NM_002746.3 |
| Reverse | CTTCAGCCGCTCCTTAGGTA | ||
| ERK2 | Forward | CCCGTCTTGGCTTATCCACT | NM_002745.5 |
| Reverse | TACATACTGCCGCAGGTCAC | ||
| p21 | Forward | TCACTGTCTTGTACCCTTGTGC | NM_001220777.2 |
| Reverse | GGCGTTTGGAGTGGTAGAAA | ||
| p27 | Forward | CTTGCCCGAGTTCTACTACAGAC | NM_004064.5 |
| Reverse | CAAATGCGTGTCCTCAGAGTTAG | ||
| p38 | Forward | TATGCGTCTGACAGGAACAC | NM_001315.3 |
| Reverse | GATCGGCCACTGGTTCATCA | ||
| p53 | Forward | CGTGTGGAGTATTTGGATGAC | NM_000546.6 |
| Reverse | TTGTAGTGGATGGTGGTACAGTC | ||
| p57 | Forward | GATTTCTTCGCCAAGCGCAA | NM_000076.2 |
| Reverse | GGGGCTCTTTGGGCTCTAAAT | ||
| CCNB1 (Cyclin B1) | Forward | TCCAGTTATGCAGCACCTGGCTA | NM_031966.4 |
| Reverse | TGCCACAGCCTTGGCTAAATCTT | ||
| β-actin | Forward | GGACTTCGAGCAAGAGATGG | NM_001199954.1 |
| Reverse | TGTGTTGGGGTACAGGTCTTTG | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, S.-Y.; Kwon, J.-E.; Kim, S.; Lee, Y.-G.; Park, S.; Kang, S.-C. Hair Regeneration Effects of Lespedeza bicolor Extract In Vivo and In Vitro. Appl. Sci. 2022, 12, 2863. https://doi.org/10.3390/app12062863
Shin S-Y, Kwon J-E, Kim S, Lee Y-G, Park S, Kang S-C. Hair Regeneration Effects of Lespedeza bicolor Extract In Vivo and In Vitro. Applied Sciences. 2022; 12(6):2863. https://doi.org/10.3390/app12062863
Chicago/Turabian StyleShin, Seung-Yeon, Jeong-Eun Kwon, Seonyu Kim, Yeong-Geun Lee, Soojin Park, and Se-Chan Kang. 2022. "Hair Regeneration Effects of Lespedeza bicolor Extract In Vivo and In Vitro" Applied Sciences 12, no. 6: 2863. https://doi.org/10.3390/app12062863
APA StyleShin, S.-Y., Kwon, J.-E., Kim, S., Lee, Y.-G., Park, S., & Kang, S.-C. (2022). Hair Regeneration Effects of Lespedeza bicolor Extract In Vivo and In Vitro. Applied Sciences, 12(6), 2863. https://doi.org/10.3390/app12062863