
Black Fungi and Stone Heritage Conservation: Ecological and Metabolic Assays for Evaluating Colonization Potential and Responses to Traditional Biocides






Abstract
:
1. Introduction
2. Materials and Methods
2.1. Black Fungal Strains Selection and Identification
2.2. Thermal Preferences, Salt Tolerance, and Metabolic Assays
2.3. Marble Substrate Investigations
2.4. Selected Chemicals
2.5. Sensitiveness to Biocides
2.6. Statistical Analyses
3. Results
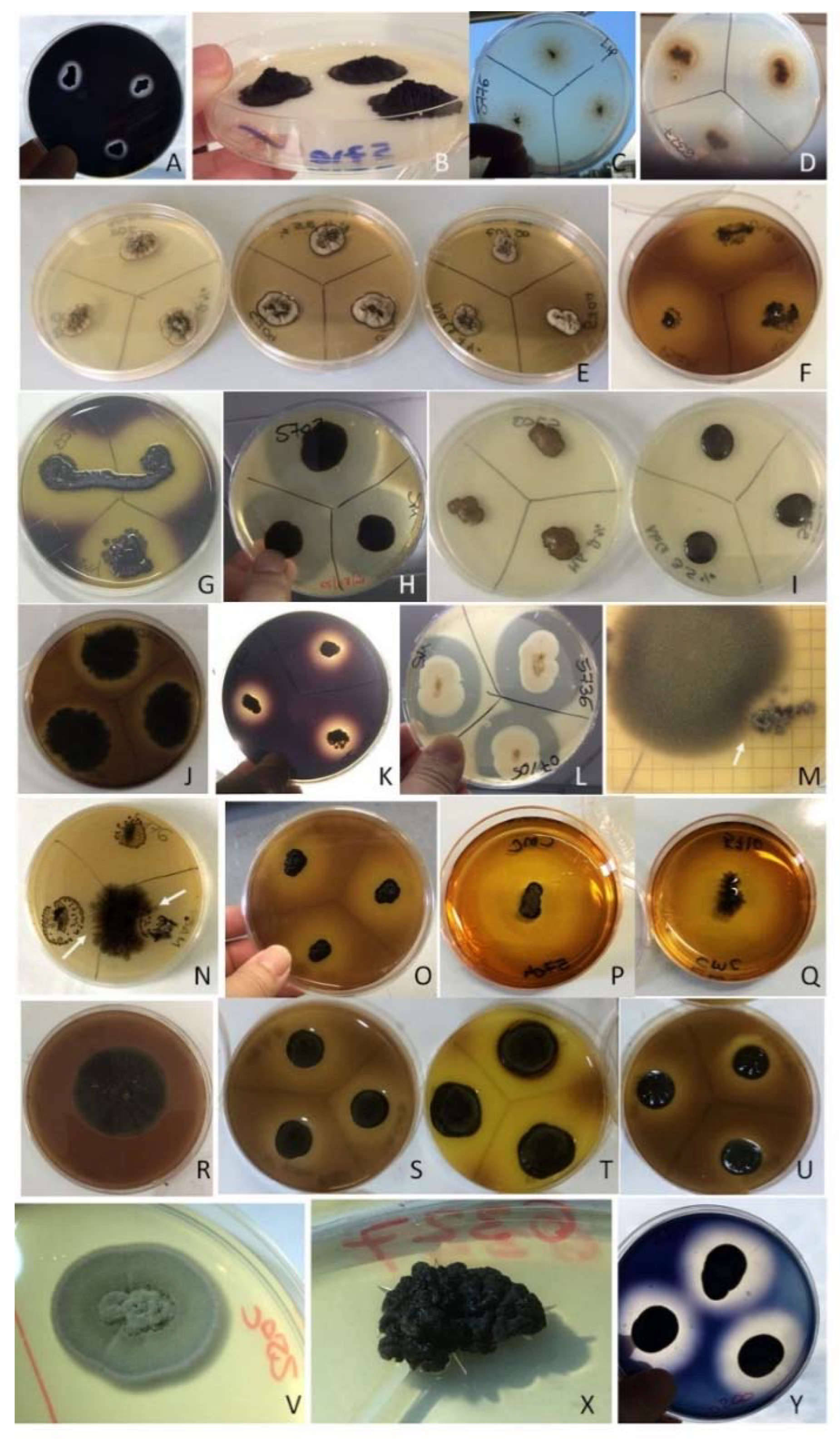
3.1. Identification and Selection of BF Species
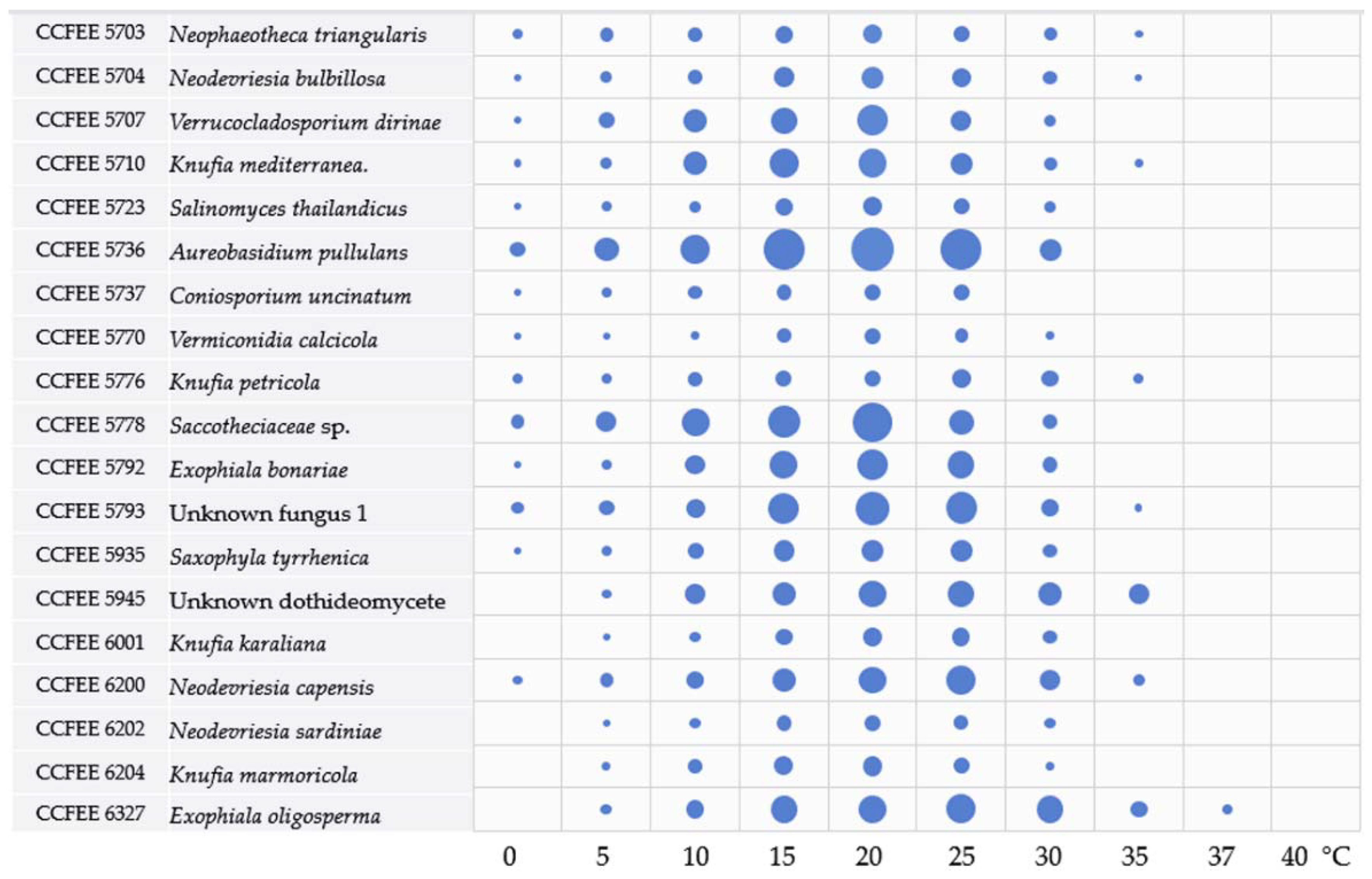
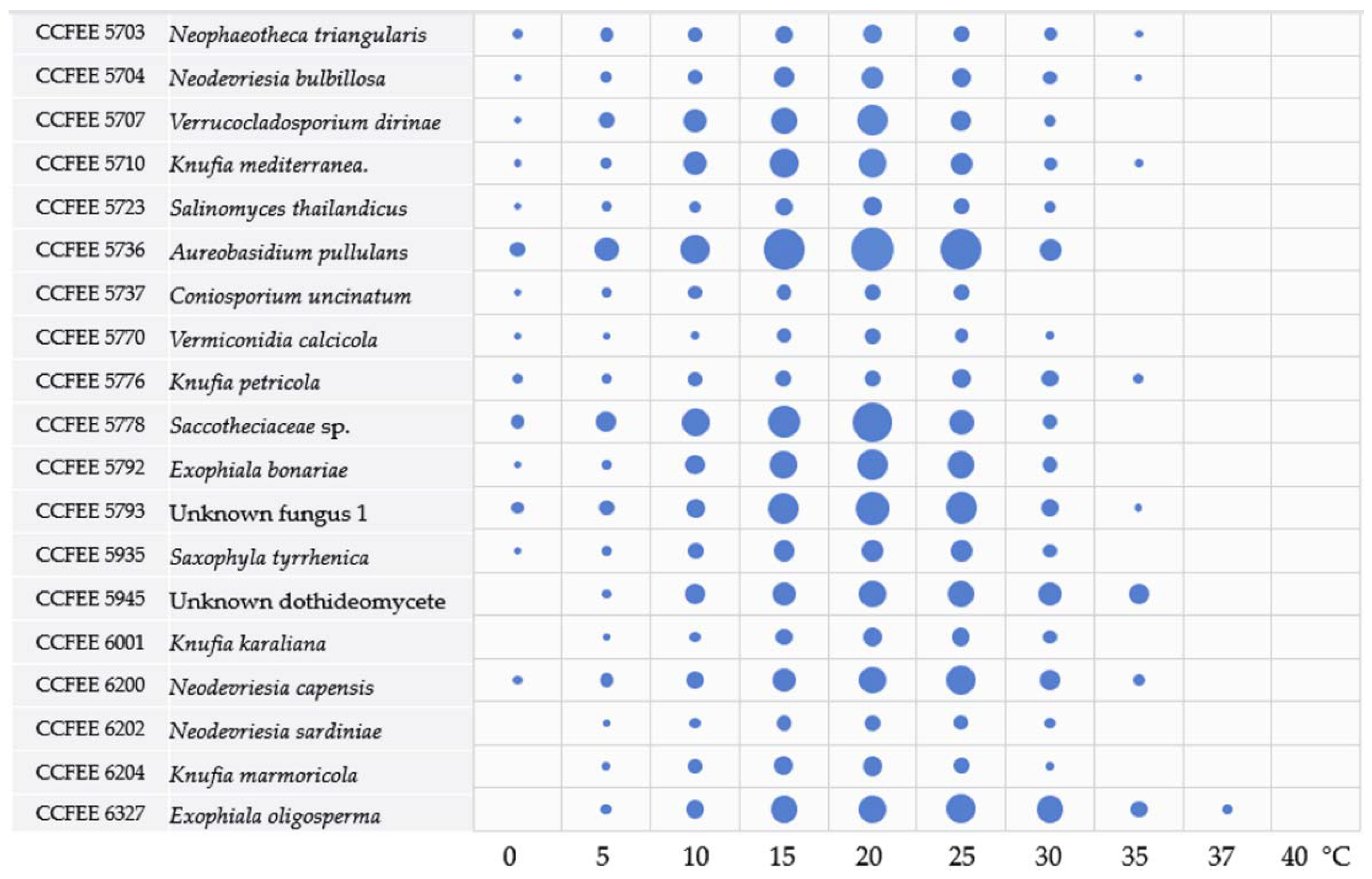
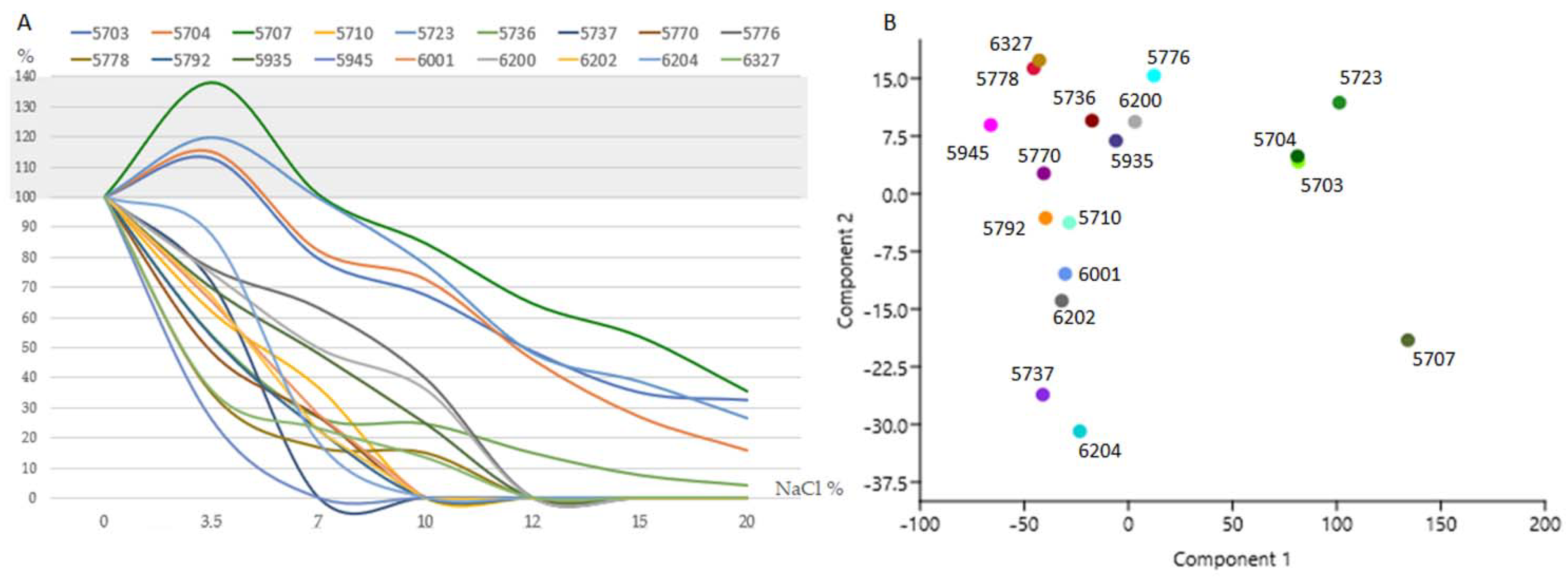
3.2. Main Ecological Traits Assessment: Thermal and Salt Tolerance
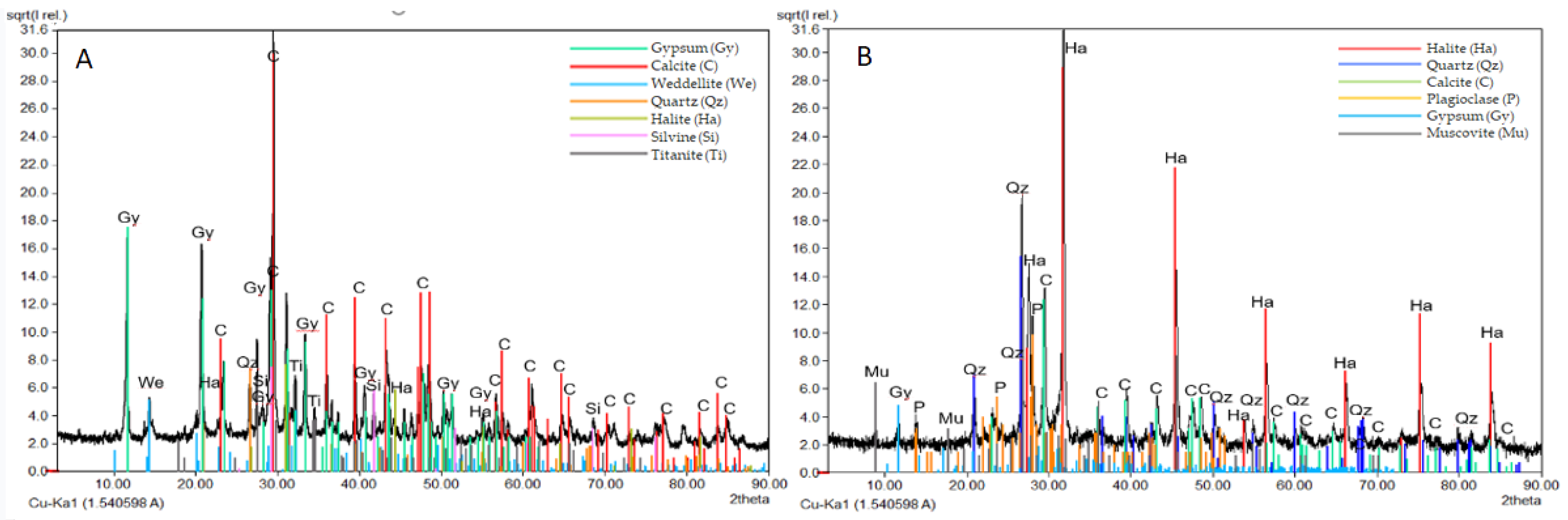
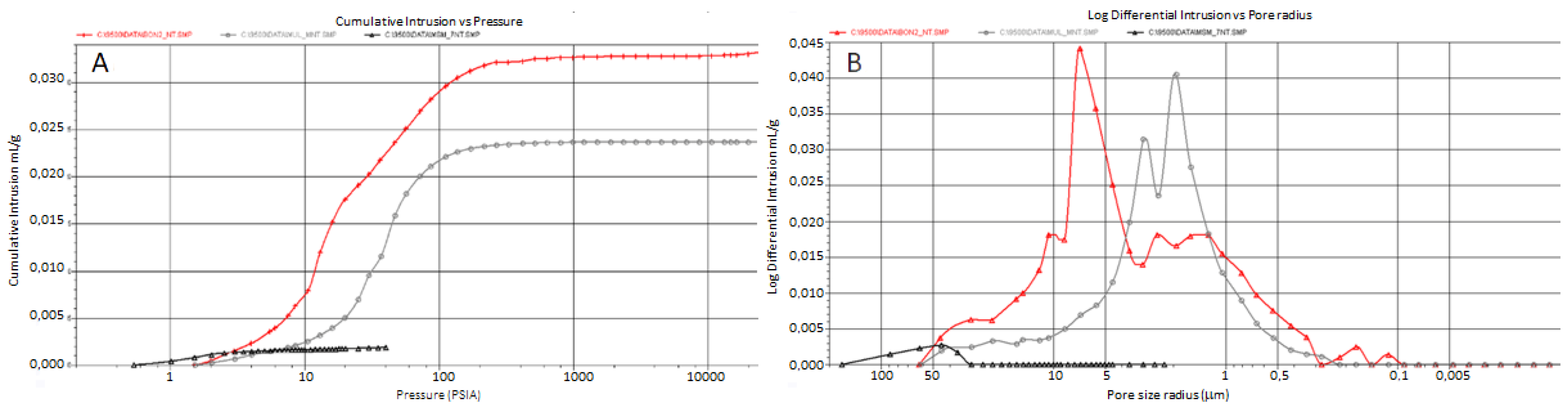
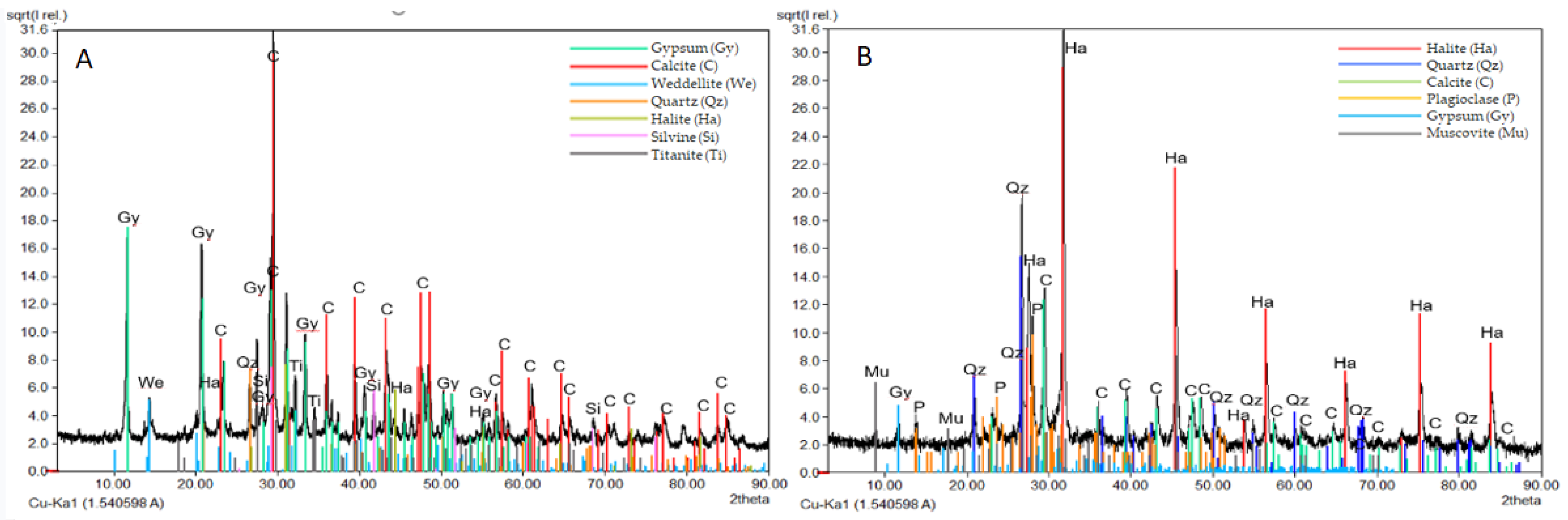
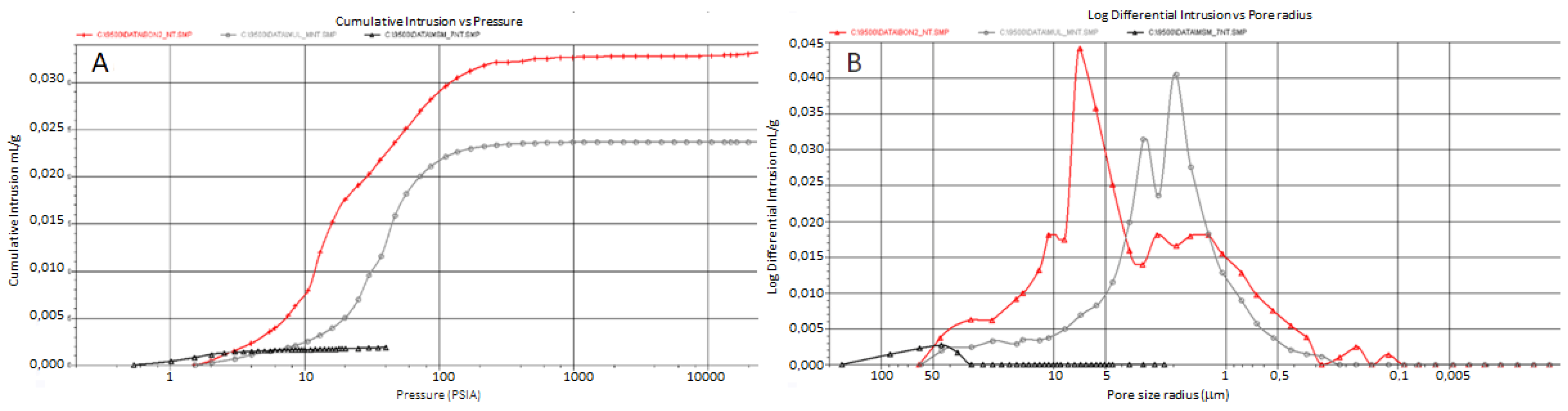
3.3. Mineralogical and Physical Investigations
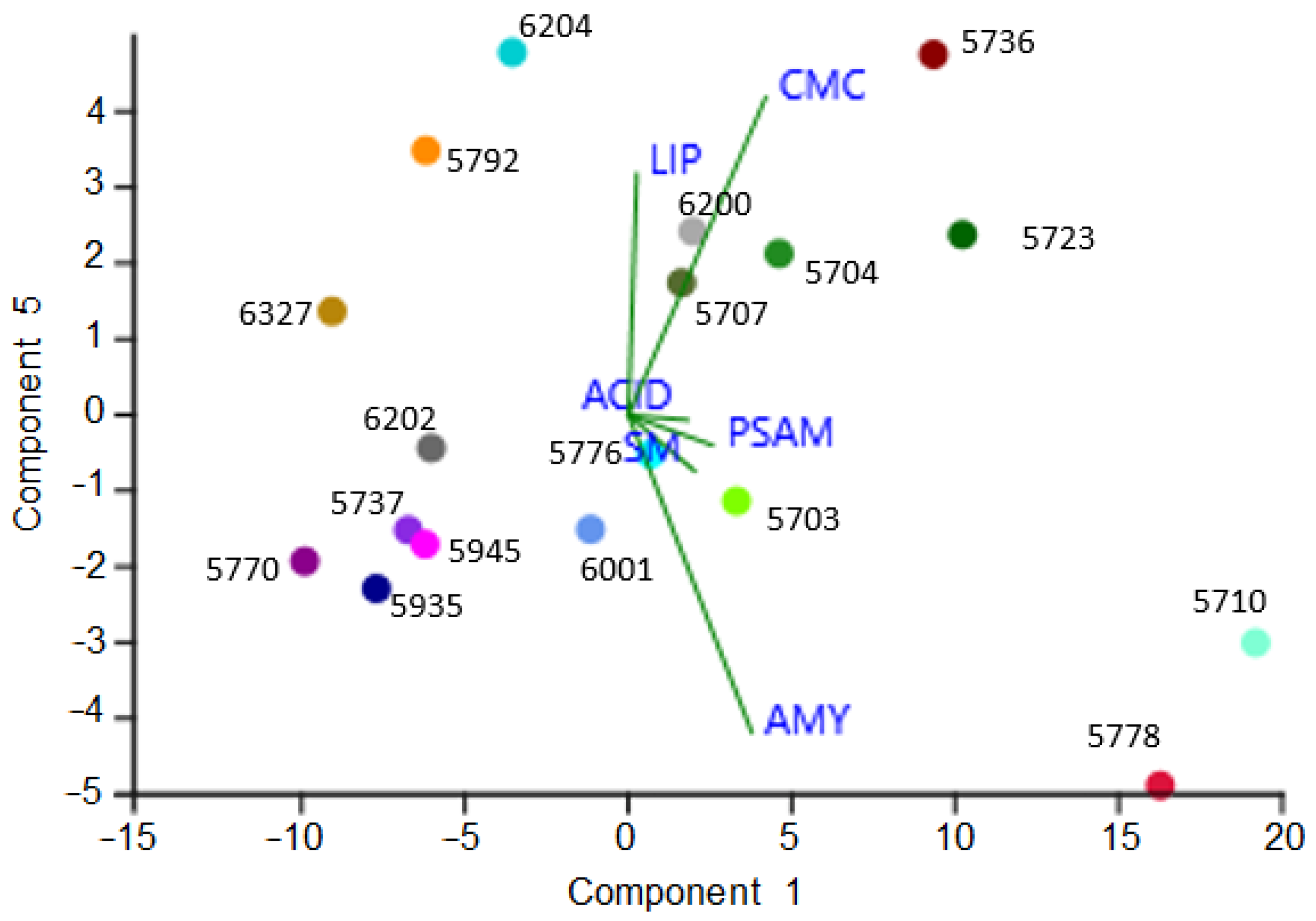
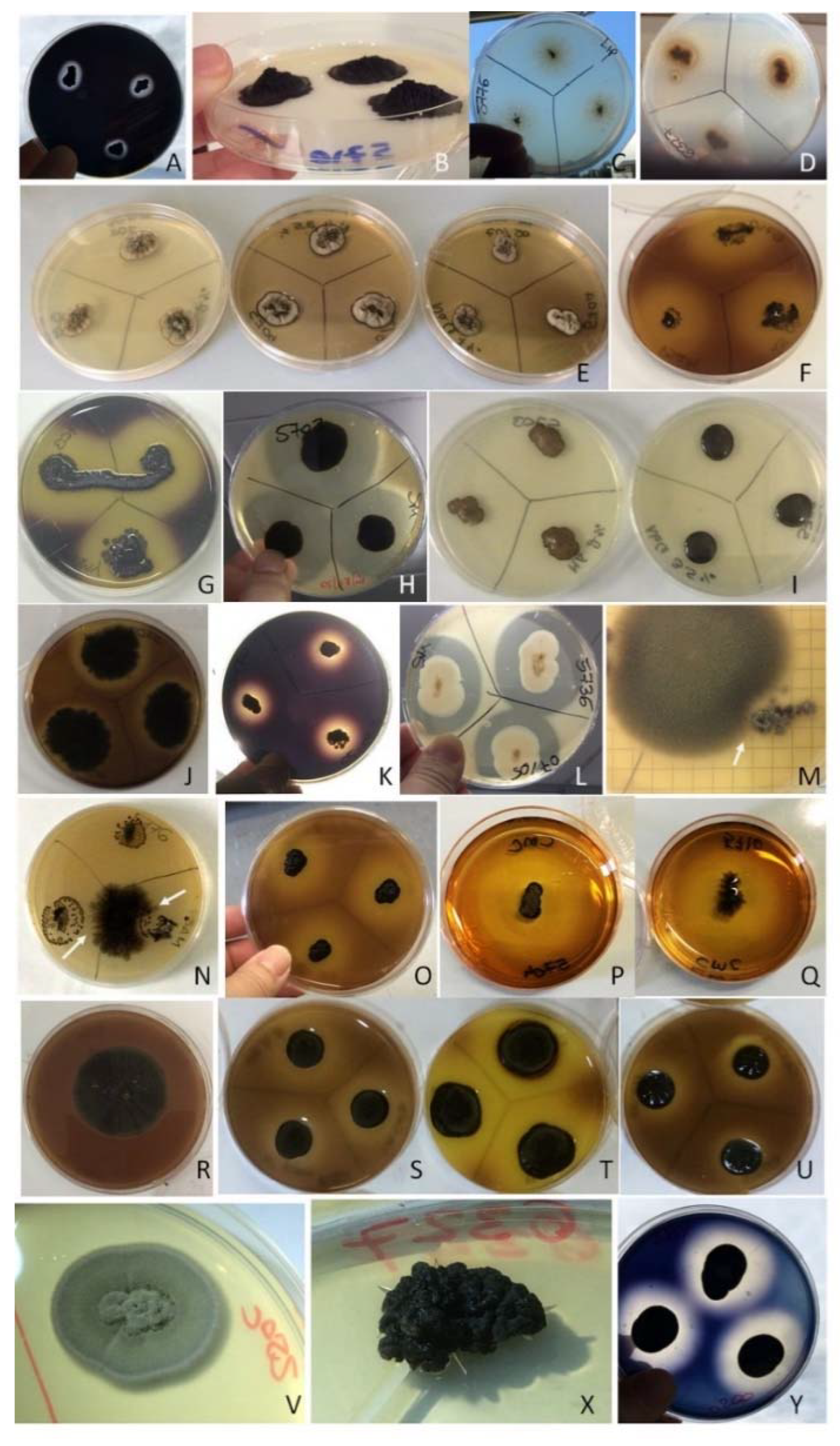
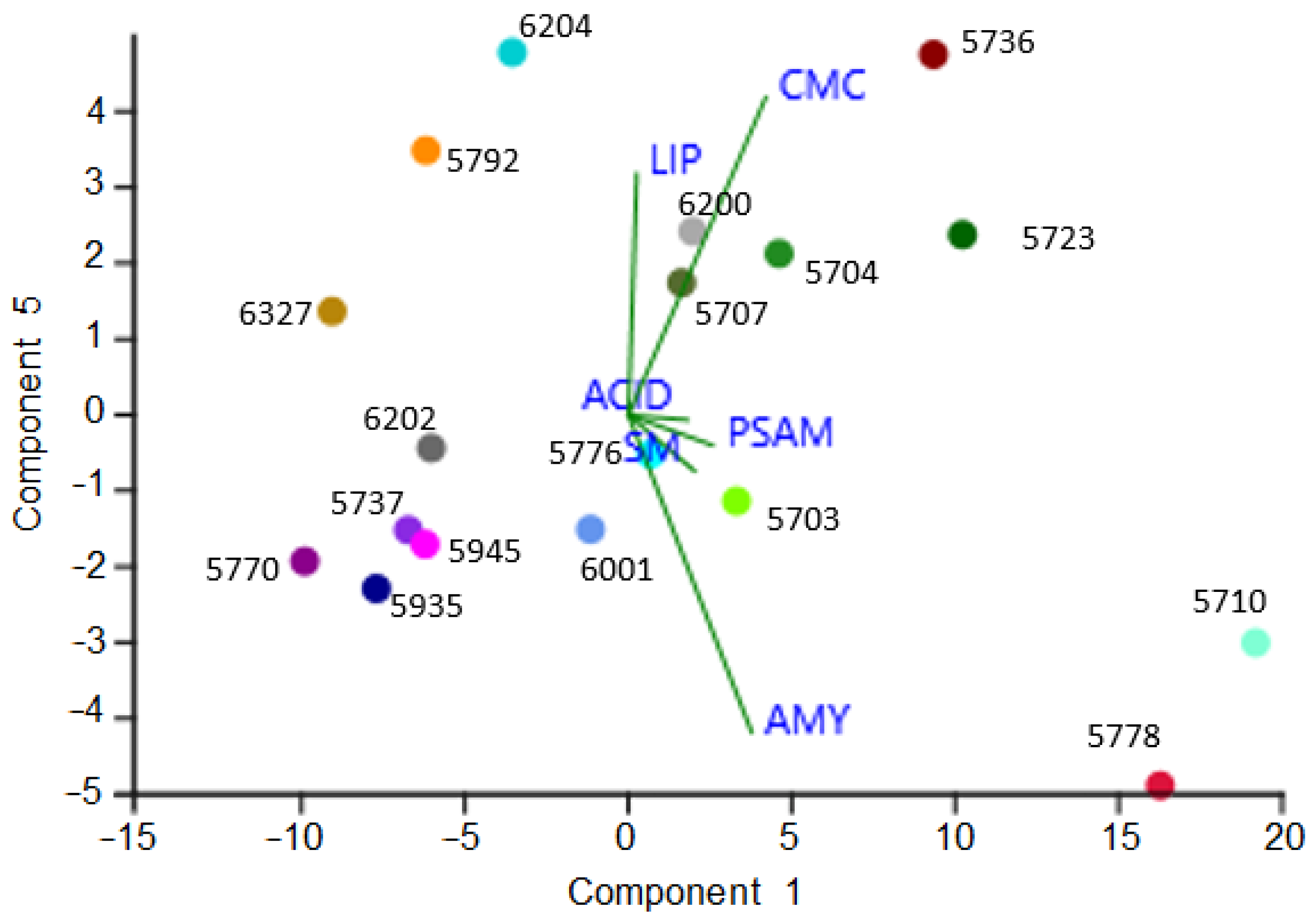
3.4. Detrimental Potential of the Investigated Species: The Metabolic Screenings
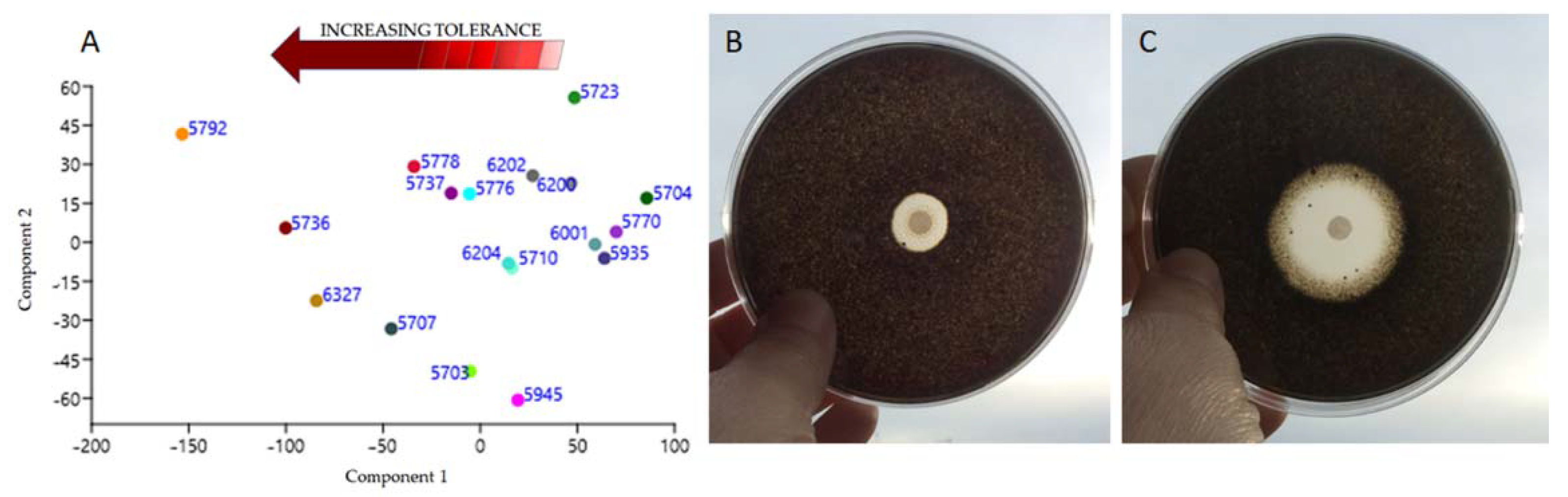
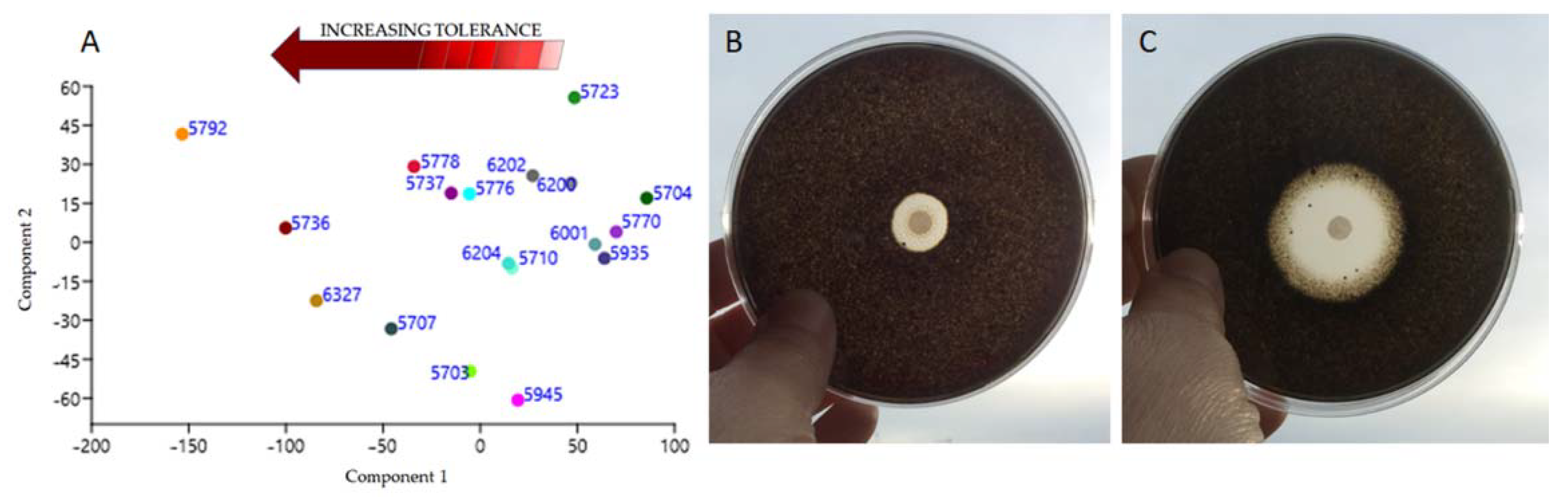
3.5. Sensitiveness to Traditional Biocides
4. Discussion
4.1. Selection and Identification of the BF Strains of Interest
4.2. Temperature and Osmotic Tolerance as Indicators of Environmental Preferences
4.3. Marble Deterioration Degree Selects the Resident Fungal Community
4.4. Metabolic Assay, Ecological Traits, and Detrimental Potential of the Investigated Species
4.5. Tolerance to Biocides
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caneva, G.; Ceschin, S. Ecology of biodeterioration. In Plant Biology for Cultural Heritage; Caneva, G., Nugari, M.P., Salvadori, O., Eds.; The Getty Conservation Institute: Los Angeles, CA, USA, 2009; pp. 35–58. [Google Scholar]
- Miller, A.Z.; Sanmartín, P.; Pereira-Pardo, L.; Dionísio, A.; Sáiz-Jiménez, C.; Macedo, M.F.; Prieto, B. Bioreceptivity of building stones: A review. Sci. Total Environ. 2012, 426, 1–12. [Google Scholar] [CrossRef]
- Sanmartín, P.; Miller, A.Z.; Prieto, B.; Viles, H.A. Revisiting and reanalysing the concept of bioreceptivity 25 years on. Sci. Total Environ. 2021, 770, 145314. [Google Scholar] [CrossRef]
- Tonon, C.; Favero-Longo, S.E.; Matteucci, E.; Piervittori, R.; Croveri, P.; Appolonia, L.; Meirano, V.; Serino, M.; Elia, D. Microenvironmental features drive the distribution of lichens in the House of the Ancient Hunt, Pompeii, Italy. Int. Biodeterior. Biodegrad. 2019, 136, 71–81. [Google Scholar] [CrossRef]
- Isola, D.; Selbmann, L.; Meloni, P.; Maracci, E.; Onofri, S.; Zucconi, L. Detrimental rock black fungi and biocides: A study on the Monumental Cemetery of Cagliari. In Science and Technology for the Conservation of Cultural Heritage; Rogerio-Candelera, M.A., Lazzari, M., Cano, E., Eds.; CRC Press: London, UK, 2013; pp. 83–86. [Google Scholar]
- Onofri, S.; Zucconi, L.; Isola, D.; Selbmann, L. Rock-inhabiting fungi and their role in deterioration of stone monuments in the Mediterranean area. Plant. Biosyst. 2014, 148, 384–391. [Google Scholar] [CrossRef]
- Tonon, C.; Breitenbach, R.; Voigt, O.; Turci, F.; Gorbushina, A.A.; Favero-Longo, S.E. Hyphal morphology and substrate porosity -rather than melanization-drive penetration of black fungi into carbonate substrates. J. Cult. Herit. 2021, 48, 244–253. [Google Scholar] [CrossRef]
- Selbmann, L.; Isola, D.; Zucconi, L.; Onofri, S. Resistance to UV-B induced DNA damage in extreme-tolerant cryptoendolithic Antarctic fungi: Detection by PCR assays. Fungal Biol. 2011, 115, 937–944. [Google Scholar] [CrossRef]
- Selbmann, L.; Grube, M.; Onofri, S.; Isola, D.; Zucconi, L. Antarctic epilithic lichens as niches for black meristematic fungi. Biology 2013, 2, 784–797. [Google Scholar] [CrossRef] [Green Version]
- Selbmann, L.; Isola, D.; Egidi, E.; Zucconi, L.; Gueidan, C.; de Hoog, G.S.; Onofri, S. Mountain tips as reservoirs for new rock-fungal entities: Saxomyces gen.nov. and four new species from Alps. Fungal Divers. 2014, 65, 167–182. [Google Scholar] [CrossRef] [Green Version]
- Marvasi, M.; Donnarumma, F.; Frandi, A.; Mastromei, G.; Sterflinger, K.; Tiano, P.; Perito, B. Black microcolonial fungi as deteriogens of two famous marble statues in Florence, Italy. Int. Biodeterior. Biodegrad. 2012, 68, 36–44. [Google Scholar] [CrossRef]
- Zakharova, K.; Tesei, D.; Marzban, G.; Dijksterhuis, J.; Wyatt, T.; Sterflinger, K. Microcolonial fungi on rocks: A life in constant drought? Mycopathologia 2013, 175, 537–547. [Google Scholar] [CrossRef] [Green Version]
- Dornieden, T.; Gorbushina, A.A.; Krumbein, W.E. Biodecay of cultural heritage as a space/time related ecological situation—An evaluation of a series of studies. Int. Biodeterior. Biodegrad. 2000, 46, 261–270. [Google Scholar] [CrossRef]
- Favero-Longo, S.E.; Gazzano, C.; Girlanda, M.; Castelli, D.; Tretiach, M.; Baiocchi, C.; Piervittori, R. Physical and chemical deterioration of silicate and carbonate rocks by meristematic microcolonial fungi and endolithic lichens (Chaetothyriomycetidae). Geomicrobiol. J. 2011, 28, 732–744. [Google Scholar] [CrossRef]
- Gorbushina, A.A.; Krumbein, W.E.; Hamann, C.H.; Panina, L.; Soukharjevski, S.; Wollenzien, U. Role of black fungi in color change and biodeterioration of antique marbles. Geomicrobiol. J. 1993, 11, 205–222. [Google Scholar] [CrossRef]
- Sert, H.B.; Sümbül, H.; Sterflinger, K. Microcolonial fungi from antique marbles in Perge/Side/Termessos (Antalya/Turkey). Antonie van Leeuwenhoek 2007, 91, 217–227. [Google Scholar] [CrossRef]
- Toreno, G.; Isola, D.; Meloni, P.; Carcangiu, G.; Selbmann, L.; Onofri, S.; Caneva, G.; Zucconi, L. Biological colonization on stone monuments: A new low impact cleaning method. J. Cult. Herit. 2018, 30, 100–109. [Google Scholar] [CrossRef]
- Caneva, G.; Isola, D.; Lee, H.J.; Chung, Y.J. Biological risk for hypogea: Shared data from Etruscan tombs in Italy and ancient tombs of the Baekje dynasty in Republic of Korea. Appl. Sci. 2020, 10, 6104. [Google Scholar] [CrossRef]
- Caneva, G.; Bartoli, F.; Savo, V.; Futagami, Y.; Strona, G. Combining statistical tools and ecological assessments in the study of biodeterioration patterns of stone temples in Angkor (Cambodia). Sci. Rep. 2016, 6, 32601. [Google Scholar] [CrossRef] [Green Version]
- Pinna, D. Coping with Biological Growth on Stone Heritage Objects: Methods, Products, Applications, and Perspectives; Apple Academic Press CRC Press: Oakville, ON, Canada, 2017; 360p. [Google Scholar]
- Caneva, G.; Nugari, M.P.; Pinna, D.; Salvadori, O. Il Controllo del Degrado Biologico: I Biocidi nel Restauro dei Materiali Lapidei; Nardini: Firenze, Italy, 1996; pp. 1–200. [Google Scholar]
- Nugari, M.P.; Salvadori, O. Biocides and treatment of stone: Limitation and future prospects. In Art, Biology and Conservation: Biodeterioration of Works of Art; The Met Fifth Avenue: New York, NY, USA, 2003; pp. 518–535. [Google Scholar]
- Young, M.E.; Alakomi, H.L.; Fortune, I.; Gorbushina, A.A.; Krumbein, W.E.; Maxwell, I.; McCullagh, C.; Robertson, P.; Saarela, M.; Valero, J.; et al. Development of a biocidal treatment regime to inhibit biological growths on cultural heritage: BIODAM. Environ. Geol. 2008, 56, 631–641. [Google Scholar] [CrossRef]
- Cappitelli, F.; Nosanchuk, J.D.; Casadevall, A.; Toniolo, L.; Brusetti, L.; Florio, S.; Principi, P.; Borin, S.; Sorlini, C. Synthetic consolidants attacked by melanin-producing fungi: Case study of the biodeterioration of Milan (Italy) Cathedral marble treated with acrylics. Appl. Environ. Microbiol. 2007, 73, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Cappitelli, F.; Villa, F.; Sanmartín, P. Interactions of microorganisms and synthetic polymers in cultural heritage conservation. Int. Biodeterior. Biodegrad. 2021, 163, 105282. [Google Scholar] [CrossRef]
- De Leo, F.; Urzì, C. Fungal colonization on treated and untreated stone surfaces. In Molecular Biology and Cultural Heritage; Saiz-Jimenez, C., Ed.; Swets & Zeitlinger BV: Lisse, The Netherlands, 2003; pp. 213–218. [Google Scholar]
- Sterflinger, K. Fungi: Their role in deterioration of cultural heritage. Fungal Biol. Rev. 2010, 24, 47–55. [Google Scholar] [CrossRef]
- Nai, C.; Wong, H.Y.; Pannenbecker, A.; Broughton, W.J.; Benoit, I.; de Vries, R.P.; Gueidan, C.; Gorbushina, A.A. Nutritional physiology of a rock-inhabiting, model microcolonial fungus from an ancestral lineage of the Chaetothyriales (Ascomycetes). Fungal Genet. Biol. 2013, 56, 54–66. [Google Scholar] [CrossRef]
- Voigt, O.; Knabe, N.; Nitsche, S.; Erdmann, E.A.; Schumacher, J.; Gorbushina, A.A. An advanced genetic toolkit for exploring the biology of the rock-inhabiting black fungus Knufia petricola. Sci. Rep. 2020, 10, 22021. [Google Scholar] [CrossRef]
- Trovão, J.; Portugal, A. Current knowledge on the fungal degradation abilities profiled through biodeteriorative plate essays. Appl. Sci. 2021, 11, 4196. [Google Scholar] [CrossRef]
- Isola, D.; Zucconi, L.; Onofri, S.; Caneva, G.; De Hoog, G.S.; Selbmann, L. Extremotolerant rock inhabiting black fungi from Italian monumental sites. Fungal Divers. 2016, 76, 75–96. [Google Scholar] [CrossRef]
- Meloni, P.; Vacca, G.; Massidda, L.; Carcangiu, G.; Mameli, P.; Cocco, O.; Toreno, G. Vulnerabilità dei beni culturali: Sistemi informatici finalizzati alla conservazione programmata e preventiva di monumenti in marmo. In Pensare la Prevenzione: Manufatti, Usi, Ambienti; Ricerche, A., Ed.; Parco Scientifico Tecnologico di Venezia: Marghera-Venezia, Italy, 2010; pp. 385–392. [Google Scholar]
- Meloni, P.; Vacca, G.; Massidda, L.; Carcangiu, G.; Mameli, P.; Toreno, G. Architettura in un Sistema GIS per la conservazione programmata delle opere all’aperto del cimitero storico di Bonaria in Cagliari (Sardegna). In Proceedings of the Atti del IV Convegno di Monitoraggio e Conservazione preventiva dei Beni Culturali, Fac. di Ingegneria, Cassino, Italy, 27–28 May 2010; pp. 76–82. [Google Scholar]
- Meloni, P.; Toreno, G.; Carcangiu, G.; Cocco, O.; Deiana, M.; Murru, A. Sinergie per la conservazione degli spazi della memoria: L’esperienza didattica dei cantieri pilota del cimitero monumentale di Bonaria in Cagliari. In Atti 2° Congresso Specialistico Internazionale sui Cimiteri Monumentali: Conoscenza, Conservazione e Restyling; Cicop Italia—Centro Internazionale per la Conservazione del Patrimonio Architettonico: Firenze, Italy, 2013; pp. 166–171. [Google Scholar]
- Isola, D.; Bartoli, F.; Langone, S.; Ceschin, S.; Zucconi, L.; Caneva, G. Plant DNA Barcode as a Tool for Root Identification in Hypogea: The Case of the Etruscan Tombs of Tarquinia (Central Italy). Plants 2021, 10, 1138. [Google Scholar] [CrossRef]
- Isola, D.; Zucconi, L.; Cecchini, A.; Caneva, G. Dark-pigmented biodeteriogenic fungi in Etruscan hypogeal tombs: New data on their culture-dependent diversity, favouring conditions, and resistance to biocidal treatments. Fungal Biol. 2021, 125, 609–620. [Google Scholar] [CrossRef]
- Gražulis, S.; Daškevič, A.; Merkys, A.; Chateigner, D.; Lutterotti, L.; Quiros, M.; Serebryanaya, N.R.; Moeck, P.; Downs, R.T.; Le Bail, A. Crystallography Open Database (COD): An open-access collection of crystal structures and platform for world-wide collaboration. Nucleic Acids Res. 2012, 40, D420–D427. [Google Scholar] [CrossRef]
- Czaja-Szewczak, B. Burial tunics from Naqlun. In Christianity and Monasticism in the Fayoum Oasis; Gabra, G., Ed.; Essays of the 2004 International Symposium of the Saint Mark Foundation and the Saint Shenouda the Archimandrite Coptic Society in Honor of Martin Krause; American University in Cairo Press: Cairo, Egypt, 2005; pp. 133–142. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 9 December 2021).
- Czachura, P.; Owczarek-Kościelniak, M.; Piątek, M. Salinomyces polonicus: A moderately halophilic kin of the most extremely halotolerant fungus Hortaea werneckii. Fungal Biol. 2021, 125, 459–468. [Google Scholar] [CrossRef]
- Isola, D.; Scano, A.; Orrù, G.; Prenafeta-Boldú, F.X.; Zucconi, L. Hydrocarbon-contaminated sites: Is there something more than Exophiala xenobiotica? New insights into black fungal diversity using the long cold incubation method. J. Fungi 2021, 7, 817. [Google Scholar] [CrossRef]
- Sterflinger, K.; Little, B.; Pinar, G.; Pinzari, F.; de los Rios, A.; Gu, J.D. Future directions and challenges in biodeterioration research on historic materials and cultural properties. Int. Biodeterior. Biodegrad. 2018, 129, 10–12. [Google Scholar] [CrossRef]
- Martin-Sanchez, P.M.; Miller, A.Z.; Saiz-Jimenez, C. Lascaux Cave: An example of fragile ecological balance in subterranean environments. In Microbial Life in Cave Systems; Engel, A.S., Ed.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2015; pp. 279–301. [Google Scholar] [CrossRef]
- Urzì, C.; De Leo, F.; Krakova, L.; Pangallo, D.; Bruno, L. Effects of biocide treatments on the biofilm community in Domitilla’s catacombs in Rome. Sci. Total Environ. 2016, 572, 252–262. [Google Scholar] [CrossRef]
- Mazzoli, R.; Giuffrida, M.G.; Pessione, E. Back to the past: “Find the guilty bug—Microorganisms involved in the biodeterioration of archeological and historical artifacts”. Appl. Microbiol. Biotechnol. 2018, 102, 6393–6407. [Google Scholar] [CrossRef]
- Jurado, V.; Gonzalez-Pimentel, J.L.; Hermosin, B.; Saiz-Jimenez, C. Biodeterioration of Salón de Reinos, Museo Nacional del Prado, Madrid, Spain. Appl. Sci. 2021, 11, 8858. [Google Scholar] [CrossRef]
- Ma, W.; Wu, F.; Tian, T.; He, D.; Zhang, Q.; Gu, J.-D.; Duan, Y.; Ma, D.; Wang, W.; Feng, H. Fungal diversity and its contribution to the biodeterioration of mural paintings in two 1700-Year-Old Tombs of China. Int. Biodeterior. Biodegrad. 2020, 152, 104972. [Google Scholar] [CrossRef]
- Savković, Ž.; Unković, N.; Stupar, M.; Franković, M.; Jovanović, M.; Erić, S.; Šarić, K.; Stanković, S.; Dimkić, I.; Vukojević, J.; et al. Diversity and biodeteriorative potential of fungal dwellers on ancient stone stela. Int. Biodeterior. Biodegrad. 2016, 115, 212–223. [Google Scholar] [CrossRef]
- Trovão, J.; Tiago, I.; Catarino, L.; Gil, F.; Portugal, A. In vitro analyses of fungi and dolomitic limestone interactions: Bioreceptivity and biodeterioration assessment. Int. Biodeterior. Biodegrad. 2020, 155, 105107. [Google Scholar] [CrossRef]
- Unković, N.; Erić, S.; Šarić, K.; Stupar, M.; Savković, Ž.; Stanković, S.; Dimkić, I.; Vukojević, J.; Grbić, M.L. Biogenesis of secondary mycogenic minerals related to wall paintings deterioration process. Micron 2017, 100, 1–9. [Google Scholar] [CrossRef]
- Sun, W.; Su, L.; Yang, S.; Sun, J.; Liu, B.; Fu, R.; Wu, B.; Liu, X.; Cai, L.; Guo, L.; et al. Unveiling the hidden diversity of rock-inhabiting fungi: Chaetothyriales from China. J. Fungi 2020, 6, 187. [Google Scholar] [CrossRef]
- Sazanova, K.V.; Zelenskaya, M.S.; Vlasov, A.D.; Bobir, S.Y.; Yakkonen, K.L.; Vlasov, D.Y. Microorganisms in superficial deposits on the stone monuments in Saint Petersburg. Microorganisms 2022, 10, 316. [Google Scholar] [CrossRef]
- Selbmann, L.; Onofri, S.; Zucconi, L.; Isola, D.; Rottigni, M.; Ghiglione, C.; Piazza, P.; Alvaro, M.C.; Schiaparelli, S. Distributional records of Antarctic fungi based on strains preserved in the Culture Collection of Fungi from Extreme Environments (CCFEE) Mycological Section associated with the Italian National Antarctic Museum (MNA). MycoKeys 2015, 10, 57. [Google Scholar] [CrossRef]
- Gostinčar, C.; Turk, M.; Zajc, J.; Gunde-Cimerman, N. Fifty Aureobasidium pullulans genomes reveal a recombining polyextremotolerant generalist. Environ. Microbiol. 2019, 21, 3638–3652. [Google Scholar] [CrossRef] [Green Version]
- Isola, D.; Selbmann, L.; de Hoog, G.S.; Fenice, M.; Onofri, S.; Prenafeta-Boldú, F.X.; Zucconi, L. Isolation and screening of black fungi as degraders of volatile aromatic hydrocarbons. Mycopathologia 2013, 175, 369–379. [Google Scholar] [CrossRef]
- Prenafeta-Boldú, F.X.; Armjio-Medina, C.; Isola, D. Black fungi in the built environment—The good, the bad, and the ugly. In Viruses, Bacteria, and Fungi in the Built Environment. Designing Healthy Indoor Environments; Pacheco-Torgal, F., Ivanov, V., Falkinham, J.O., Eds.; Woodhead Publishing: Duxford, UK, 2021; pp. 65–99. [Google Scholar]
- Gostinčar, C.; Stajich, J.E.; Zupancic, J.; Zalar, P.; Gunde-Cimerman, N. Genomic evidence for intraspecific hybridization in a clonal and extremely halotolerant yeast. BMC Genom. 2018, 19, 364. [Google Scholar] [CrossRef]
- Gostinčar, C.; Zajc, J.; Lenassi, M.; Plemenitas, A.; de Hoog, S.; Al Hatmi, A.M.S.; Gunde-Cimerman, N. Fungi between extremotolerance and opportunistic pathogenicity on humans. Fungal Divers. 2018, 93, 195–213. [Google Scholar] [CrossRef] [Green Version]
- Gostinčar, C.; Zalar, P.; Gunde-Cimerman, N. No need for speed: Slow development of fungi in extreme environments. Fungal Biol. Rev. 2022, 39, 1–14. [Google Scholar] [CrossRef]
- Kogej, T.; Ramos, J.; Plemenitas, A.; Gunde-Cimerman, N. The halophilic fungus Hortaea werneckii and the halotolerant fungus Aureobasidium pullulans maintain low intracellular cation concentrations in hypersaline environments. Appl. Environ. Microbiol. 2005, 71, 6600–6605. [Google Scholar] [CrossRef] [Green Version]
- Butinar, L.; Sonjak, S.; Zalar, P.; Plemenitaš, A.; Gunde-Cimerman, N. Melanized halophilic fungi are eukaryotic members of microbial communities in hypersaline waters of solar salterns. Botanica Marina 2005, 48, 73–79. [Google Scholar] [CrossRef]
- Egidi, E.; De Hoog, G.S.; Isola, D.; Onofri, S.; Quaedvlieg, W.; De Vries, M.; Verkley, G.J.M.; Stielow, J.B.; Zucconi, L.; Selbmann, L. Phylogeny and taxonomy of meristematic rock-inhabiting black fungi in the Dothideomycetes based on multi-locus phylogenies. Fungal Divers. 2014, 65, 127–165. [Google Scholar] [CrossRef]
- Crous, P.W.; Braun, U.; Schubert, K.; Groenewald, J.Z. Delimiting Cladosporium from morphologically similar genera. Stud. Mycol. 2007, 58, 33–56. [Google Scholar] [CrossRef]
- Piñar, G.; Dalnodar, D.; Voitl, C.; Reschreiter, H.; Sterflinger, K. Biodeterioration risk threatens the 3100 year old staircase of Hallstatt (Austria): Possible involvement of halophilic microorganisms. PLoS ONE 2016, 11, e0148279. [Google Scholar] [CrossRef]
- Gnavi, G.; Garzoli, L.; Poli, A.; Prigione, V.; Burgaud, G.; Varese, G.C. The culturable mycobiota of Flabellia petiolata: First survey of marine fungi associated to a Mediterranean green alga. PLoS ONE 2017, 12, e0175941. [Google Scholar] [CrossRef]
- Owczarek-Kościelniak, M.; Krzewicka, B.; Piątek, J.; Kołodziejczyk, Ł.M.; Kapusta, P. Is there a link between the biological colonization of the gravestone and its deterioration? Int. Biodeterior. Biodegrad. 2020, 148, 104879. [Google Scholar] [CrossRef]
- Casti, M.; Meloni, P.; Pia, G.; Palomba, M. Differential damage in the semi-confined Munazio Ireneo cubicle in Cagliari (Sardinia): A correlation between damage and microclimate. Environ. Earth Sci. 2017, 76, 529. [Google Scholar] [CrossRef]
- Murru, A.; Freire-Lista, D.M.; Fort, R.; Varas-Muriel, M.J.; Meloni, P. Evaluation of post-thermal shock effects in Carrara marble and Santa Caterina di Pittinuri limestone. Constr. Build. Mater. 2018, 186, 1200–1211. [Google Scholar] [CrossRef]
- Silva, B.; Rivas, T.; Prieto, B. Relation between type of soluble salt and decay forms in granitic coastal churches in Galicia (NW Spain). In Proceedings of the Origin, Mechanisms and Effects of Salts on Degradation of Monuments in Marine and Continental Environments, Bari, Italy, 25–27 March 1996; Zezza, Ed.; pp. 181–190. [Google Scholar]
- Stryszewska, T. The change in selected properties of ceramic materials obtained from ceramic brick treated by the sulphate and chloride ions. Constr. Build. Mater. 2014, 66, 268–274. [Google Scholar] [CrossRef]
- Vázquez-Nion, D.; Troiano, F.; Sanmartín, P.; Valagussa, C.; Cappitelli, F.; Prieto, B. Secondary bioreceptivity of granite: Effect of salt weathering on subaerial biofilm growth. Mater. Struct. 2018, 51, 158. [Google Scholar] [CrossRef]
- Zanardini, E.; May, E.; Purdy, K.J.; Murrell, J.C. Nutrient cycling potential within microbial communities on culturally important stoneworks. Environ. Microbiol. Rep. 2019, 11, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Saiz-Jimenez, C.; Laiz, L. Occurrence of halotolerant/halophilic bacterial communities in deteriorated monuments. Int. Biodeterior. Biodegrad. 2000, 46, 319–326. [Google Scholar] [CrossRef]
- Piñar, G.; Ripka, K.; Weber, J.; Sterflinger, K. The micro-biota of a sub-surface monument the medival chapel of St. Virgil (Vienna, Austria). Int. Biodeterior. Biodegrad. 2009, 63, 851–859. [Google Scholar] [CrossRef]
- Adamiak, J.; Otlewska, A.; Gutarowska, B. Halophilic microbial communities in deteriorated buildings. World J. Microbiol. Biotechnol. 2015, 31, 1489–1499. [Google Scholar] [CrossRef]
- Adamiak, J.; Bonifay, V.; Otlewska, A.; Sunner, J.A.; Beech, I.B.; Stryszewska, T.; Kańka, S.; Oracz, J.; Żyżelewicz, D.; Gutarowska, B. Untargeted metabolomics approach in halophiles: Understanding the biodeterioration process of building materials. Front. Microbiol. 2017, 8, 2448. [Google Scholar] [CrossRef]
- Piñar, G.; Ettenauer, J.; Sterflinger, K. La vie en rose: A review of the rosy discoloration of subsurface monuments. In The Conservation of Subterranean Cultural Heritage; Saiz-Jimenez, C., Ed.; CRC Press/Balkema: Leiden, The Netherlands, 2014; pp. 113–124. [Google Scholar]
- Otlewska, A.; Adamiak, J.; Stryszewska, T.; Kanka, S.; Gutarowska, B. Factors determining biodiversity of halophilic microorganisms on historic masonry buildings. Microbes Environ. 2017, 32, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Sanmartín, P.; DeAraujo, A.; Vasanthakumar, A. Melding the old with the new: Trends in methods used to identify, monitor, and control microorganisms on cultural heritage materials. Microb Ecol. 2018, 76, 64–80. [Google Scholar] [CrossRef]
- Gerrits, R.; Pokharel, R.; Breitenbach, R.; Radnik, J.; Feldmann, I.; Schuessler, J.A.; von Blanckenburg, F.; Gorbushina, A.A.; Schott, J. How the rock-inhabiting fungus K. petricola A95 enhances olivine dissolution through attachment. Geochim. Cosmochim. Acta 2020, 282, 76–97. [Google Scholar] [CrossRef]
- Chertov, O.; Gorbushina, A.; Deventer, B. A model for microcolonial fungi growth on rock surfaces. Ecol. Modell 2004, 177, 415–426. [Google Scholar] [CrossRef]
- Salvadori, O.; Municchia, A.C. The role of fungi and lichens in the biodeterioration of stone monuments. Open Conf. Proc. J. 2016, 7, 39–54. [Google Scholar] [CrossRef]
- He, D.; Wu, F.; Ma, W.; Zhang, Y.; Gu, J.D.; Duan, Y.; Xu, R.; Feng, H.; Wang, W.; Li, S.W. Insights into the bacterial and fungal communities and microbiome that causes a microbe outbreak on ancient wall paintings in the Maijishan Grottoes. Int. Biodeterior. Biodegrad. 2021, 163, 105250. [Google Scholar] [CrossRef]
- Zucconi, L.; Gagliardi, M.; Isola, D.; Onofri, S.; Andaloro, M.C.; Pelosi, C.; Pogliani, P.; Selbmann, L. Biodeterioration agents dwelling in or on the wall paintings of the Holy Saviour’s cave (Vallerano, Italy). Int. Biodeterior. Biodegrad. 2012, 70, 40–46. [Google Scholar] [CrossRef]
- Zucconi, L.; Canini, F.; Isola, D.; Caneva, G. Fungi affecting wall paintings: A meta-analysis of their diversity. Appl. Sci. 2022; under review. [Google Scholar]
- Suphaphimol, N.; Suwannarach, N.; Purahong, W.; Jaikang, C.; Pengpat, K.; Semakul, N.; Yimklan, S.; Jongjitngam, S.; Jindasu, S.; Thiangtham, S.; et al. Identification of microorganisms dwelling on the 19th century Lanna mural paintings from Northern Thailand using culture-dependent and-independent approaches. Biology 2022, 11, 228. [Google Scholar] [CrossRef]
- Selbmann, L.; Isola, D.; Fenice, M.; Zucconi, L.; Sterflinger, K.; Onofri, S. Potential extinction of Antarctic endemic fungal species as a consequence of global warming. Sci. Total Environ. 2012, 438, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Quan, Y.; Muggia, L.; Moreno, L.F.; Wang, M.; Al-Hatmi, A.M.; da Silva Menezes, N.; Shi, D.; Deng, S.; Ahmed, S.; Hyde, K.D.; et al. A re-evaluation of the Chaetothyriales using criteria of comparative biology. Fungal Divers. 2020, 103, 47–85. [Google Scholar] [CrossRef]
- Quan, Y.; van den Ende, B.G.; Shi, D.; Prenafeta-Boldú, F.X.; Liu, Z.; Al-Hatmi, A.M.; Ahmed, S.A.; Verweij, P.E.; Kang, Y.; de Hoog, S. A comparison of isolation methods for black fungi degrading aromatic toxins. Mycopathologia 2019, 184, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Baglioni, P.; Berti, D.; Bonini, M.; Carretti, E.; Dei, L.; Fratini, E.; Giorgi, R. Micelle, microemulsions, and gels for the conservation of cultural heritage. Adv. Colloid Interface Sci. 2014, 205, 361–371. [Google Scholar] [CrossRef]
- Gagliano Candela, R.; Maggi, F.; Lazzara, G.; Rosselli, S.; Bruno, M. The essential oil of Thymbra capitata and its application as a biocide on stone and derived surfaces. Plants 2019, 8, 300. [Google Scholar] [CrossRef] [Green Version]
- Fidanza, M.R.; Caneva, G. Natural biocides for the conservation of stone cultural heritage: A review. J. Cultl. Herit. 2019, 38, 271–286. [Google Scholar] [CrossRef]
- Lo Schiavo, S.; De Leo, F.; Urzì, C. Present and future perspectives for biocides and antifouling products for stone-built cultural heritage: Ionic liquids as a challenging alternative. Appl. Sci. 2020, 10, 6568. [Google Scholar] [CrossRef]
- Pinna, D.; Salvadori, B.; Galeotti, M. Monitoring the performance of innovative and traditional biocides mixed with consolidants and water-repellents for the prevention of biological growth on stone. Sci. Total Environ. 2012, 423, 132–141. [Google Scholar] [CrossRef]
- Salvadori, O.; Charola, A.E. Methods to Prevent Biocolonization and Recolonization: An Overview of Current Research for Architectural and Archaeological Heritage; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2011; pp. 37–50. [Google Scholar]
- Urzì, C.; De Leo, F.; Galletta, M.; Salamone, P.; Balzarotti, R. Efficiency of biocide in “in situ” and “in vitro” treatment. Study case of the “Templete de Mudejar”, Guadalupe, Spain. In Proceedings of the 9th International Congress on Deterioration and Conservation of Stone Venice June 19–24; Elsevier Science BV: Amsterdam, The Netherlands, 2000; pp. 531–539. [Google Scholar]
- Delgado Rodrigues, J.; Valero Congil, J.; Wakefield, R.; Brechet, E.; Larrañaga, I. Monitoraggio della biocolonizzazione e valutazione dell’efficacia di un biocida. In Arkos: Scienza e Restauro Dell’architettura; Nardini: Firenze, Italy, 2004; Volume 5, pp. 52–58. [Google Scholar]
- Tesei, D. Black fungi research: Out-of-this-world implications. Encyclopedia 2022, 2, 212–229. [Google Scholar] [CrossRef]
- Onofri, S.; Selbmann, L.; Barreca, D.; Isola, D.; Zucconi, L. Do fungi survive under actual space conditions? Searching for evidence in favor of the lithopansperimia. Plant. Biosyst. 2009, 143, S85–S87. [Google Scholar] [CrossRef]
- Onofri, S.; de la Torre, R.; de Vera, J.P.; Ott, S.; Zucconi, L.; Selbmann, L.; Scalzi, G.; Venkateswaran, K.J.; Rabbow, E.; Sánchez Iñigo, F.J.; et al. Survival of rock-colonizing organisms after 1.5 years in outer space. Astrobiol 2012, 12, 508–516. [Google Scholar] [CrossRef]
- Acero, F.J.; Carbú, M.; El-Akhal, M.R.; Garrido, C.; González-Rodríguez, V.E.; Cantoral, J.M. Development of proteomics-based fungicides: New strategies for environmentally friendly control of fungal plant diseases. Int. J. Mol. Sci. 2011, 12, 795–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, P.; Moore, L.E. Cationic antiseptics: Diversity of action under a common epithet. J. Appl. Microbiol. 2005, 99, 703–715. [Google Scholar] [CrossRef]
- Caneva, G.; Nugari, M.P.; Salvadori, O. Plant. Biology for Cultural Heritage: Biodeterioration and Conservation; Getty Publications: Los Angeles, CA, USA, 2008. [Google Scholar]
- Corbett, J.R.; Wright, K.; Baillie, A.C. The Biochemical Mode of Action of Pesticides, 2nd ed.; Academic Press: London, UK, 1984. [Google Scholar]
- Juergensen, L.; Busnarda, J.; Caux, P.Y.; Kent, R. Fate, behavior, and aquatic toxicity of the fungicide IPCB in the Canadian environment. Environ. Toxicol. 2000, 15, 201–213. [Google Scholar] [CrossRef]
- Williams, T.M. The mechanism of action of isothiazolone biocides. Power Plant. Chem. 2007, 9, 14–22. [Google Scholar]
- Scheerer, S.; Ortega-Morales, O.; Gaylarde, C. Microbial deterioration of stone monuments—An updated overview. Adv. Appl. Microbiol. 2009, 66, 97–139. [Google Scholar] [CrossRef]
- Gostinčar, C.; Grube, M.; Gunde-Cimerman, N. Evolution of fungal pathogens in domestic environments? Fungal Biol. 2011, 115, 1008–1018. [Google Scholar] [CrossRef]
- Langsrud, S.; Sundheim, G.; Borgmann-Strahsen, R. Intrinsic and acquired resistance to quaternary ammonium compounds in food-related Pseudomonas spp. J. Appl. Microbiol. 2003, 95, 874–882. [Google Scholar] [CrossRef]
- Ruibal, C.; Selbmann, L.; Avci, S.; Martin-Sanchez, P.M.; Gorbushina, A.A. Roof-inhabiting cousins of rock-inhabiting fungi: Novel melanized microcolonial fungal species from photocatalytically reactive subaerial surfaces. Life 2018, 8, 30. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | CCFEE | Funerary Monument |
|---|---|---|
| Aureobasidium pullulans | 5736 | Giuseppina Ara dei conti Ciarella |
| Coniosporium uncinatum | 5737 | Francesca Warzee |
| Exophiala bonariae (T) | 5792 | Zelina Ferrà Gastaldi Millelire |
| Exophiala oligosperma | 6327 | Anna Maria Frau Carta |
| Knufia karalitana | 6001 | Unknown tombstone |
| Knufia marmoricola | 6204 | Anna Maria Frau Carta |
| Knufia mediterranea | 5710 | Avv. Giuseppe Todde |
| Knufia petricola | 5776 | Giuseppina Ara dei conti Ciarella |
| Neodevriesia bulbillosa | 5704 | Ignazio Ruda Roych conte di San Lorenzo |
| Neodevriesia capensis | 6200 | Anna Maria Frau Carta |
| Neodevriesia sardiniae (T) | 6202 | Anna Maria Frau Carta |
| Neophaeotheca triangularis | 5703 | Ignazio Ruda Roych conte di San Lorenzo |
| Saccotheciaceae sp. | 5778 | Zelina Ferrà Gastaldi Millelire |
| Salinomyces thailandicus * | 5723 | Antonio Viganigo |
| Saxophyla tyrrhenica (T) | 5935 | Unknown burial |
| Unknown dothideomycete | 5945 | Rossino Bolla’s spouses |
| Vermiconidia calcicola (T) | 5770 | Giuseppina Ara dei conti Ciarella |
| Verrucocladosporium dirinae | 5707 | Ignazio Ruda Roych conte di San Lorenzo |
| Strain No. | Species | ACID | LIP | AMY | SM | PSAM | CMC |
|---|---|---|---|---|---|---|---|
| 5703 | Neophaeotheca triangularis | − | ++ | + | ++++ | + | ++ |
| 5704 | Neodevriesia bulbillosa | − | +++ | +++ | − | ++++ | +++ |
| 5707 | Verrucocladosporium dirinae | − | ++ | + | ++++ | + | ++ |
| 5710 | Knufia mediterranea. | +++ | +++ | ++++ | +++ | ++ | +++ |
| 5723 | Salinomyces thailandicus | + | + | ++ | − | − | ++++ |
| 5736 | Aureobasidium pullulans | + | + | ++ | ++ | − | ++++ |
| 5737 | Coniosporium uncinatum | − | +++ | + | + | + | − |
| 5770 | Vermiconidia calcicola (T) | − | +++ | + | − | − | − |
| 5776 | Knufia petricola | +++ | +++ | + | ++ | ++ | + |
| 5778 | Saccotheciaceae sp. | + | ++ | ++++ | + | ++ | ++++ |
| 5792 | Exophiala bonariae (T) | − | ++++ | − | − | +++ | − |
| 5935 | Saxophyla tyrrhenica (T) | − | + | − | − | − | + |
| 5945 | Unknown dothideomycete | − | ++ | + | − | − | + |
| 6001 | Knufia karalitana | ++ | ++ | ++ | − | − | ++ |
| 6200 | Neodevriesia capensis | +++ | ++++ | ++ | + | + | ++ |
| 6202 | Neodevriesia sardiniae (T) | ++ | ++ | + | + | − | + |
| 6204 | Knufia marmoricola | + | ++ | − | − | − | ++ |
| 6327 | Exophiala oligosperma | − | ++ | − | − | − | − |
| CCFEE Strain No. | Species | Biocides | |||
|---|---|---|---|---|---|
| PREV 3% | BZC 3% | LICH 1% | BioR.5% | ||
| 5703 | Neoph. triangularis | 22.8 ± 0.86 | 29.5 ± 0.52 | 45.3 ± 0.62 | 90 ± 0 |
| 5704 | N. bulbillosa | 57.2 ±0.4 | 59.3 ± 0.61 | 76.12 ± 0.52 | 90 ± 0 |
| 5707 | V. dirinae | 21.7 ± 1.1 | 20.6 ± 0.7 | 39.5 ± 1.01 | 74.5 ± 0.5 |
| 5710 | K. mediterranea | 54.3 ±0.79 | 41.5 ± 0.33 | 45.3 ± 0.75 | 90 ± 0 |
| 5723 | Sal. thailandicus | 76.7 ±0.6 | 49.3 ± 0.5 | 63.86 ± 0.81 | 90 ± 0 |
| 5736 | A. pullulans | 26.3 ±1.1 | 13.6 ± 0.91 | 38.82 ± 0.63 | 46.52 ± 0.6 |
| 5737 | C. uncinatum | 56.7 ±0.8 | 30.1 ± 0.55 | 61.875 ± 0.9 | 76.85 ± 1.02 |
| 5770 | V. calcicola | 58.8 ±1.03 | 63.5 ± 0.76 | 59.61 ± 0.62 | 90 ± 0 |
| 5776 | K. petricola | 57.8 ±0.92 | 35.2 ± 1.03 | 40.32 ± 0.3 | 79.23 ± 0.5 |
| 5778 | Saccotheciaceae sp. | 49 ± 0.7 | 30.4 ± 0.98 | 39.82 ± 0.71 | 62.03 ± 1.03 |
| 5792 | E. bonariae | 26.5 ±1.1 | 26.5 ± 0.98 | 17.53 ± 1.03 | 16.06 ± 0.2 |
| 5935 | S. tyrrhenica | 56.5 ± 0.7 | 39.28 ± 0.25 | 68.89 ± 1.05 | 90 ± 0 |
| 5945 | Unknown dothideomycete | 32.3 ±0.84 | 28.8 ± 0.8 | 46.53 ± 0.63 | 90 ± 0 |
| 6001 | K. karalitana | 64.13 ± 0.71 | 29.68 ± 0.33 | 90 ± 0 | 90 ± 0 |
| 6200 | N. capensis | 47.6 ± 0.2 | 68.7 ± 1.1 | 67.58 ± 0.62 | 90 ± 0 |
| 6202 | N. sardiniae | 56.26 ± 0.08 | 40.3 ± 0.74 | 76.74 ± 0.72 | 84.1 ± 0.1 |
| 6204 | K. marmoricola | 50.6 ± 0.3 | 36.8 ± 0.42 | 53.7 ± 0.94 | 90 ± 0 |
| 6327 | E. oligosperma | 29.49 ± 0.68 | 14.97 ± 0.43 | 29.49 ± 0.68 | 66.78 ± 0.8 |

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isola, D.; Bartoli, F.; Meloni, P.; Caneva, G.; Zucconi, L. Black Fungi and Stone Heritage Conservation: Ecological and Metabolic Assays for Evaluating Colonization Potential and Responses to Traditional Biocides. Appl. Sci. 2022, 12, 2038. https://doi.org/10.3390/app12042038
Isola D, Bartoli F, Meloni P, Caneva G, Zucconi L. Black Fungi and Stone Heritage Conservation: Ecological and Metabolic Assays for Evaluating Colonization Potential and Responses to Traditional Biocides. Applied Sciences. 2022; 12(4):2038. https://doi.org/10.3390/app12042038
Chicago/Turabian StyleIsola, Daniela, Flavia Bartoli, Paola Meloni, Giulia Caneva, and Laura Zucconi. 2022. "Black Fungi and Stone Heritage Conservation: Ecological and Metabolic Assays for Evaluating Colonization Potential and Responses to Traditional Biocides" Applied Sciences 12, no. 4: 2038. https://doi.org/10.3390/app12042038
APA StyleIsola, D., Bartoli, F., Meloni, P., Caneva, G., & Zucconi, L. (2022). Black Fungi and Stone Heritage Conservation: Ecological and Metabolic Assays for Evaluating Colonization Potential and Responses to Traditional Biocides. Applied Sciences, 12(4), 2038. https://doi.org/10.3390/app12042038