
Probiotic Molecules That Inhibit Inflammatory Diseases

Abstract
:Featured Application
Abstract
1. Introduction Mod
2. Cell Envelope Molecules
3. Secreted Molecules
4. EPSBs as Probiotic
- A.
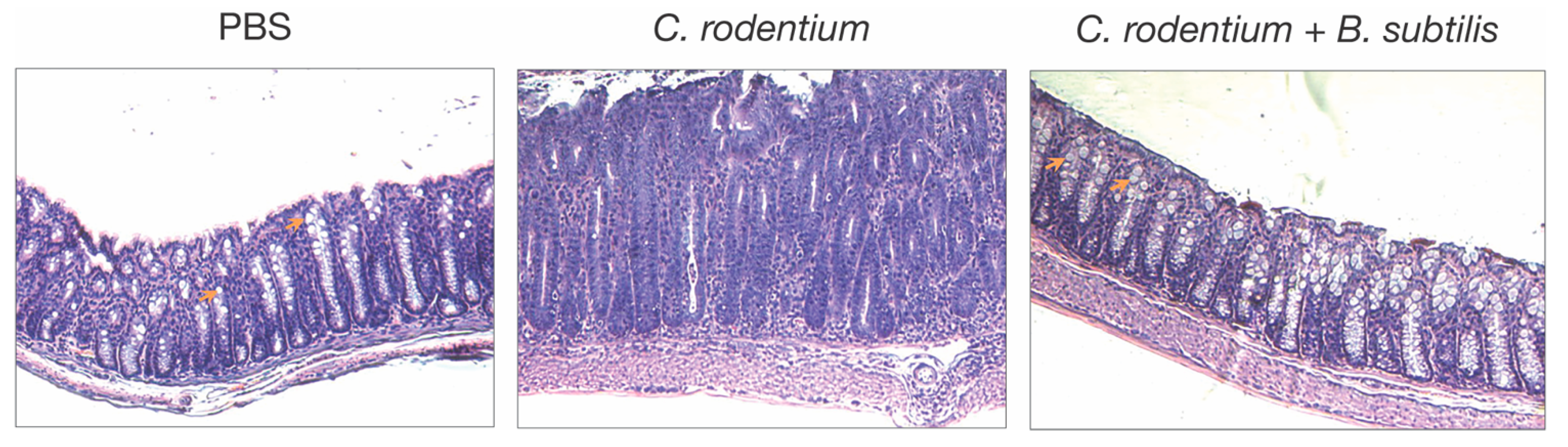
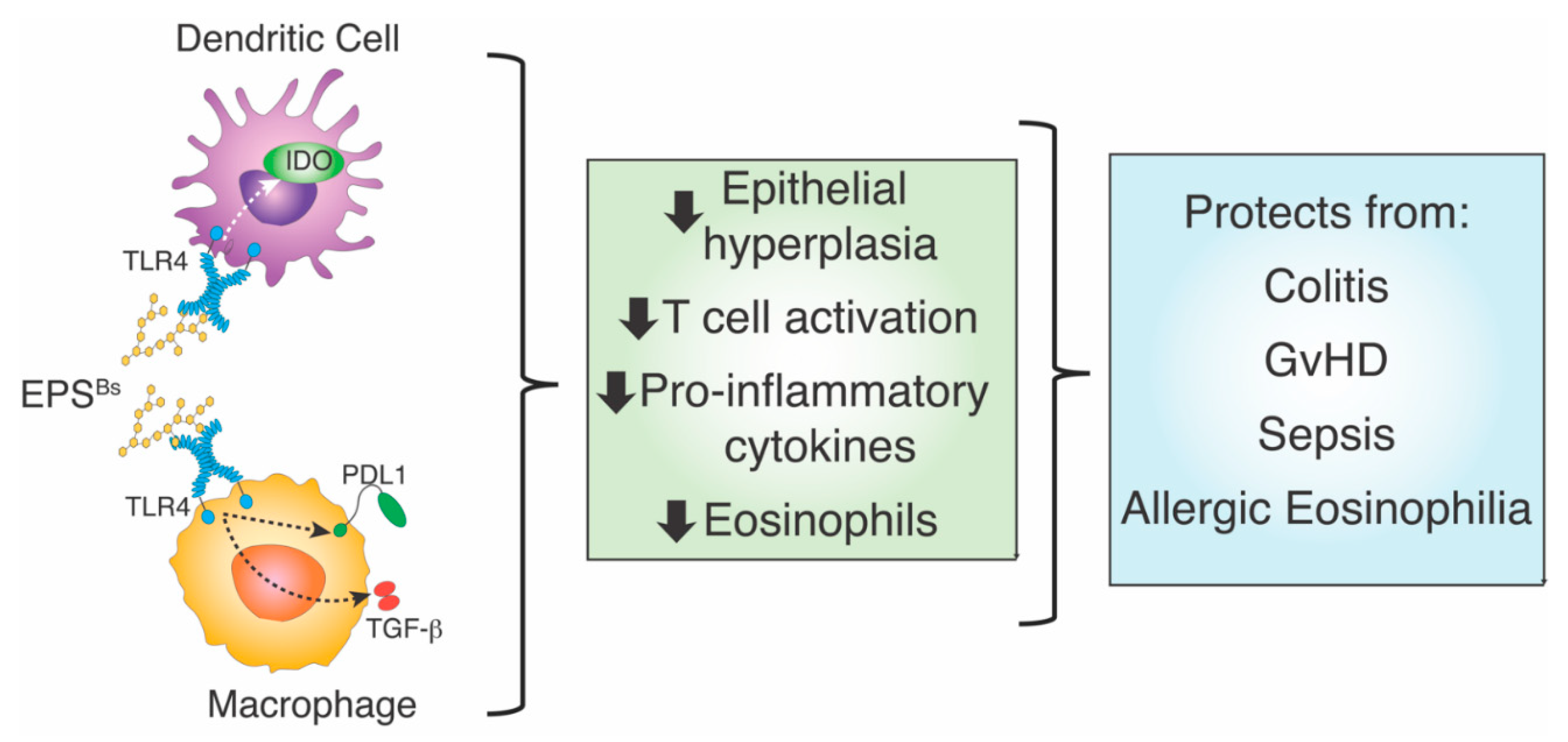
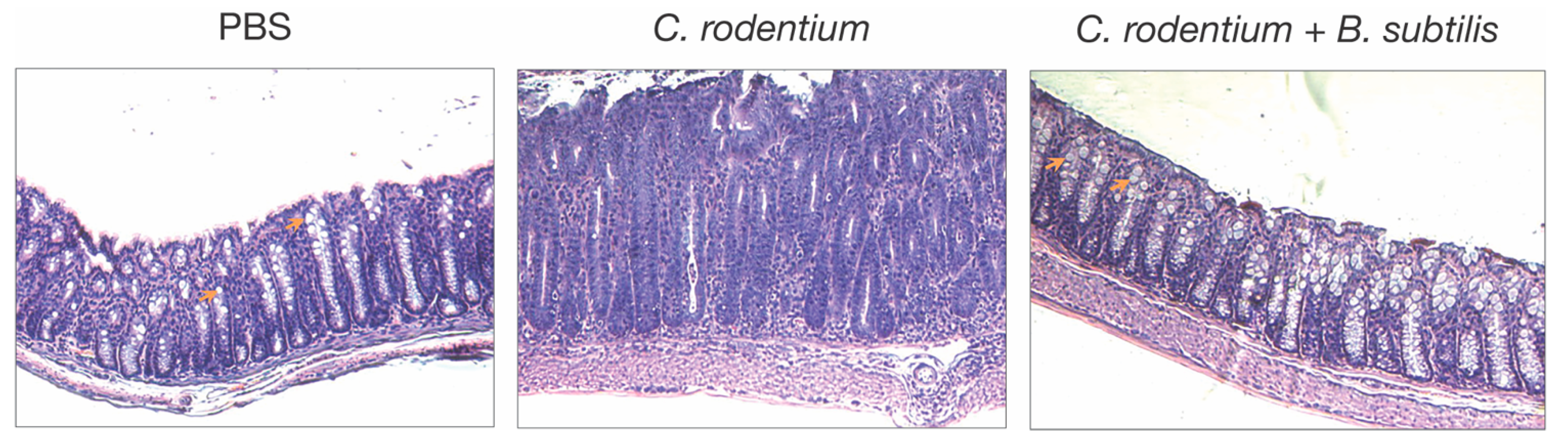
- C. rodentium-induced colitis. Oral administration of a single dose of B. subtilis spores was first shown to reduce colitis in mice after infection with the enteric pathogen, Citrobacter rodentium [9]. In this model, B. subtilis does not function by reducing the colonization of C. rodentium, but instead alters the inflammatory disease process, as indicated by reduced epithelial hyperplasia, diarrhea, and goblet cell loss (Figure 1). Analysis of B. subtilis mutants revealed that a mutation in epsH, which regulates biofilm synthesis [68], did not protect from disease caused by C. rodentium, suggesting that biofilm-associated carbohydrate exopolysaccharide (EPSBs) was required for protection. EPSBs was isolated and purified by treatment with DNase, RNase, proteinase K, and gel filtration [10,12], and indeed, it protected from disease. In fact, a single intraperitoneal injection of EPSBs (2.5 mg/kg) administered one day prior to or as much as 3 days after infection with C. rodentium, was sufficient to reduce epithelial hyperplasia, diarrhea, and goblet cell loss. Protection by EPSBs is mediated by anti-inflammatory macrophages, sometimes designated as M2 macrophages. Intraperitoneal administration of EPSBs results in the accumulation of macrophages with M2 macrophage markers, IL4Ra, CD206, arginase, and PD-L1 in the peritoneum, and adoptive transfer of these cells to untreated mice protects them from colitis after infection with C. rodentium [10,12]. These findings demonstrate the anti-inflammatory potential of EPSBs, and of the anti-inflammatory macrophages induced by EPSBs.
- B.
- Systemic infection with Staphylococcus aureus. Similar to infection with the enteric pathogen, C. rodentium, EPSBs also moderates disease caused by infection with blood-borne S. aureus [11]. In this case, EPSBs increases survival by reducing weight loss and systemic inflammation, as evidenced by decreased levels of inflammatory cytokines and chemokines in blood and bacterial burden [11]. EPSBs induced hybrid-like M1/M2 macrophages, which not only inhibited T cell activation, characteristic of M2 macrophages but also inhibited S. aureus growth through reactive oxygen species (ROS), characteristic of M1 macrophages [69]. Together, data from infection by C. rodentium and S. aureus show that EPSBs from B. subtilis induces an anti-inflammatory environment with decreased inflammatory cytokines and increased anti-inflammatory macrophages that limit T cell activation, as well as macrophages that restrict the growth of bacteria.
- C.
- Allergic eosinophilia. The association of changes in microbiota to allergic disease is well known, not only because of the hygiene hypothesis [70] but also because of a landmark study by Stein et al., who showed that children who grow up in a farm environment with close proximity to farm animals develop considerably fewer allergies than children that grow up without much interaction with farm animals [71]. This “farm effect” is likely explained by the interaction of children with microbes of the farm animals [72]. Swartzendruber et al. orally administered B. subtilis spores to mice and showed that they prevented the development of allergic eosinophilia in response to intranasal administration of house dust mite (HDM) antigen [8]. The infiltration of eosinophils is due in part to cytokines secreted by T cells [73]. Because DCs are also crucial for the activation of T cells and the development of eosinophilia, Swartzendruber et al. hypothesized that EPSBs-treated DCs could mitigate the allergic eosinophilia caused by an allergy to HDM. Intranasal adoptive transfer of EPSBs-treated bone marrow-derived DCs (BMDCs) prevented eosinophilia induced by HDM-pulsed DCs, indicating that EPSBs induces anti-inflammatory DCs, which can prevent an allergic response, as might be predicted by previous studies [70,72].
- D.
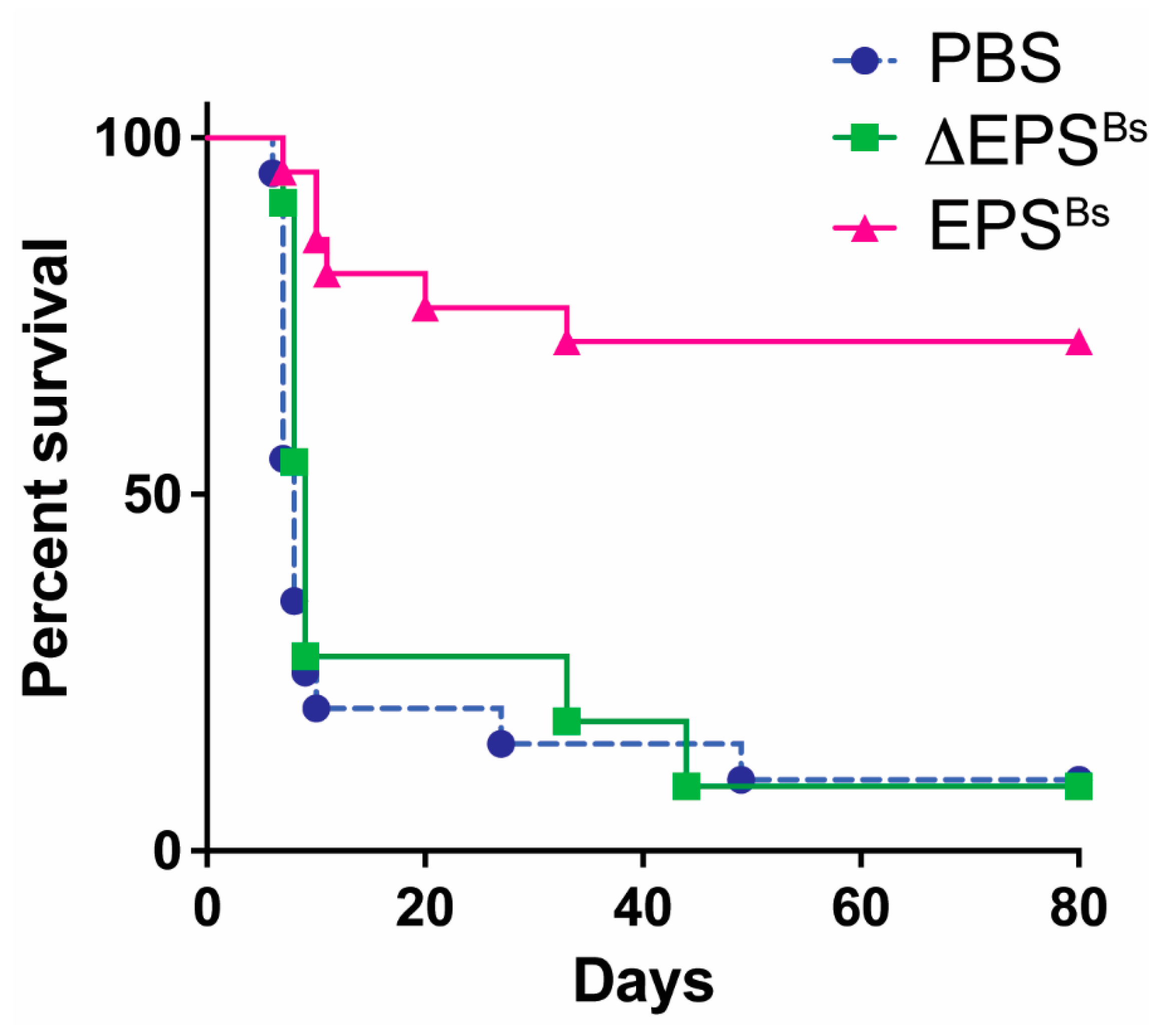
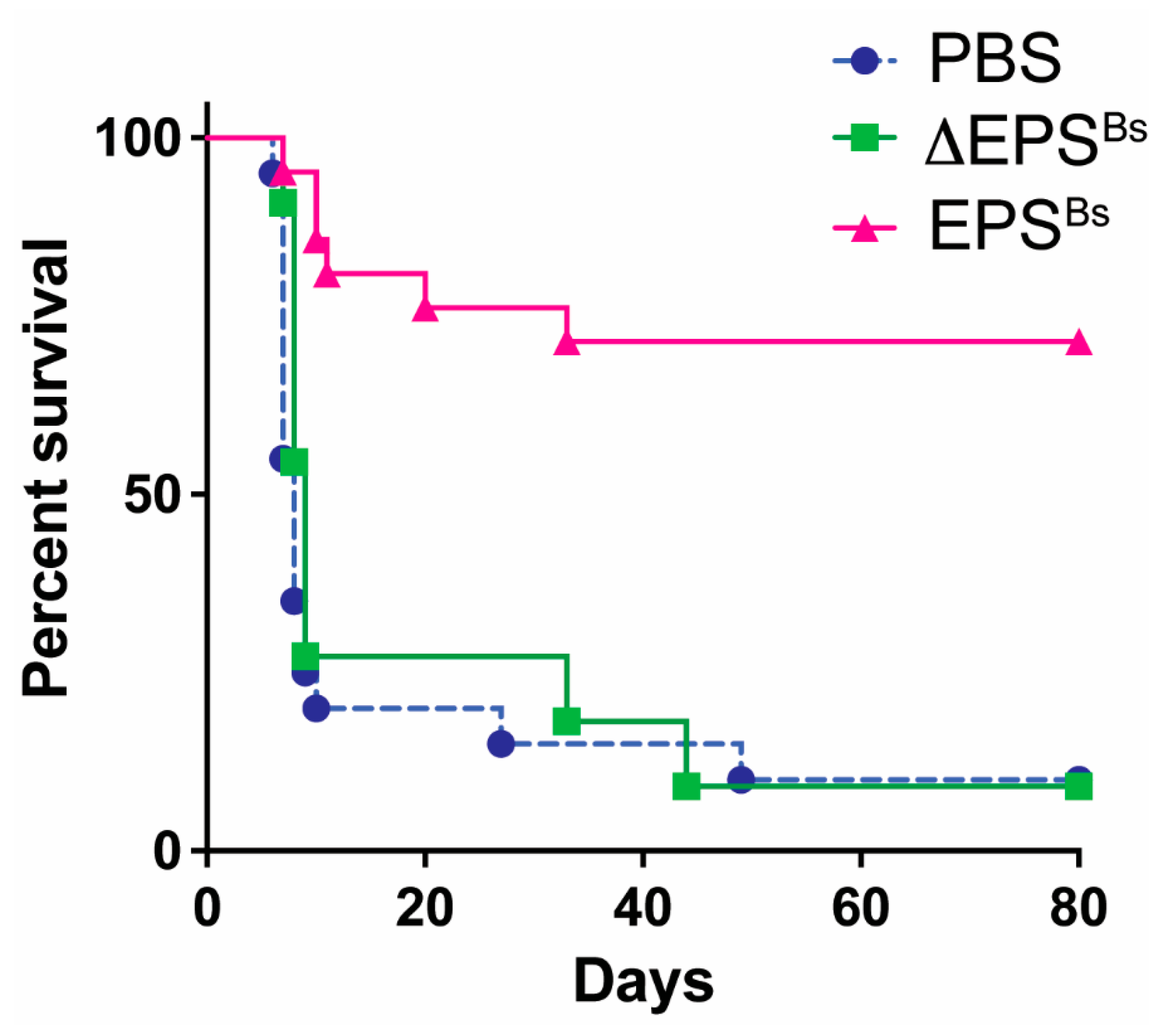
- Graft versus host disease (GvHD). Another T cell-mediated disease attenuated by EPSBs is GvHD, a severe and often lethal complication of hematopoietic stem cell transplantation, which is frequently used to treat leukemia. The devastating effects of GvHD are mediated by alloreactive donor T cells that recognize host antigens as foreign, become activated, and destroy host tissues and organs. Intraperitoneal injection of EPSBs (2.5 mg/kg) administered several times, 7, 5, and 3 days prior to induction of GvHD, increased survival of mice 80 days post GvHD from 10% to 70% (Figure 2). Kalinina et al. assessed inflammation in live mice during GvHD using a caspase-1 reporter mouse to measure inflammasome activation [7]. With this biosensor mouse model, they found that the administration of EPSBs prevented the activation of alloreactive donor T cells, explaining the increased survival of mice. The results showed that EPSBs did not directly affect alloreactive T cells. In mixed lymphocyte reactions (MLR) in vitro, EPSBs-treated BMDCs potently inhibited alloreactive T cells, suggesting that in vivo, EPSBs induces DCs or other innate cells to become inhibitory and prevent the activation of alloreactive T cells, thereby reducing GvHD.
5. Mechanism by Which EPSBs Inhibits Inflammation
5.1. Cells
5.2. Immune Regulator Molecules
6. Translational Potential of EPSBs
7. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Clarke, T.C.; Black, L.I.; Stussman, B.J.; Barnes, P.M.; Nahin, R.L. Trends in the use of complementary health approaches among adults: United States, 2002–2012. Natl. Health Stat. Rep. 2015, 79, 1–16. [Google Scholar]
- Venugopalan, V.; Shriner, K.A.; Wong-Beringer, A. Regulatory oversight and safety of probiotic use. Emerg. Infect. Dis. 2010, 16, 1661–1665. [Google Scholar] [CrossRef] [PubMed]
- Suez, J.; Zmora, N.; Segal, E.; Elinav, E. The pros, cons, and many unknowns of probiotics. Nat. Med. 2019, 25, 716–729. [Google Scholar] [CrossRef]
- Ford, A.C.; Quigley, E.M.; Lacy, B.E.; Lembo, A.J.; Saito, Y.A.; Schiller, L.R.; Soffer, E.E.; Spiegel, B.M.; Moayyedi, P. Efficacy of prebiotics, probiotics, and synbiotics in irritable bowel syndrome and chronic idiopathic constipation: Systematic review and meta-analysis. Am. J. Gastroenterol. 2014, 109, 1547–1561. [Google Scholar] [CrossRef]
- Li, L.; Han, Z.; Niu, X.; Zhang, G.; Jia, Y.; Zhang, S.; He, C. Probiotic Supplementation for Prevention of Atopic Dermatitis in Infants and Children: A Systematic Review and Meta-analysis. Am. J. Clin. Dermatol. 2019, 20, 367–377. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, J.G.; Chain, F.; Martín, R.; Bermúdez-Humarán, L.G.; Courau, S.; Langella, P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Fact. 2017, 16, 79. [Google Scholar] [CrossRef] [Green Version]
- Kalinina, O.; Talley, S.; Zamora-Pineda, J.; Paik, W.; Campbell, E.M.; Knight, K.L. Amelioration of Graft-versus-Host Disease by Exopolysaccharide from a Commensal Bacterium. J. Immunol. 2021, 206, 2101–2108. [Google Scholar] [CrossRef]
- Swartzendruber, J.A.; Incrocci, R.W.; Wolf, S.A.; Jung, A.; Knight, K.L. Bacillus subtilis exopolysaccharide prevents allergic eosinophilia. Allergy 2019, 74, 819–821. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.E.; Knight, K.L. Bacillus subtilis-mediated protection from Citrobacter rodentium-associated enteric disease requires espH and functional flagella. Infect. Immun. 2012, 80, 710–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paynich, M.L.; Jones-Burrage, S.E.; Knight, K.L. Exopolysaccharide from Bacillus subtilis Induces Anti-Inflammatory M2 Macrophages That Prevent T Cell-Mediated Disease. J. Immunol. 2017, 198, 2689–2698. [Google Scholar] [CrossRef] [Green Version]
- Paik, W.; Alonzo, F., 3rd; Knight, K.L. Probiotic Exopolysaccharide Protects against Systemic Staphylococcus aureus Infection, Inducing Dual-Functioning Macrophages That Restrict Bacterial Growth and Limit Inflammation. Infect. Immun. 2019, 87, e00791-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, S.E.; Paynich, M.L.; Kearns, D.B.; Knight, K.L. Protection from intestinal inflammation by bacterial exopolysaccharides. J. Immunol. 2014, 192, 4813–4820. [Google Scholar] [CrossRef] [Green Version]
- Paik, W.; Alonzo, F., 3rd; Knight, K.L. Suppression of Staphylococcus aureus Superantigen-Independent Interferon Gamma Response by a Probiotic Polysaccharide. Infect. Immun 2020, 88, e00661-19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Fang, L.; Wu, H.M.; Ding, P.S.; Xu, K.; Liu, R.Y. Mer receptor tyrosine kinase negatively regulates lipoteichoic acid-induced inflammatory response via PI3K/Akt and SOCS3. Mol. Immunol. 2016, 76, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; McLoughlin, R.M.; Cobb, B.A.; Charrel-Dennis, M.; Zaleski, K.J.; Golenbock, D.; Tzianabos, A.O.; Kasper, D.L. A bacterial carbohydrate links innate and adaptive responses through Toll-like receptor 2. J. Exp. Med. 2006, 203, 2853–2863. [Google Scholar] [CrossRef] [Green Version]
- Ochoa-Repáraz, J.; Mielcarz, D.W.; Ditrio, L.E.; Burroughs, A.R.; Begum-Haque, S.; Dasgupta, S.; Kasper, D.L.; Kasper, L.H. Central nervous system demyelinating disease protection by the human commensal Bacteroides fragilis depends on polysaccharide A expression. J. Immunol. 2010, 185, 4101–4108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazmanian, S.K.; Round, J.L.; Kasper, D.L. A microbial symbiosis factor prevents intestinal inflammatory disease. Nature 2008, 453, 620–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Round, J.L.; Lee, S.M.; Li, J.; Tran, G.; Jabri, B.; Chatila, T.A.; Mazmanian, S.K. The Toll-like receptor 2 pathway establishes colonization by a commensal of the human microbiota. Science 2011, 332, 974–977. [Google Scholar] [CrossRef] [Green Version]
- Ochoa-Repáraz, J.; Mielcarz, D.W.; Wang, Y.; Begum-Haque, S.; Dasgupta, S.; Kasper, D.L.; Kasper, L.H. A polysaccharide from the human commensal Bacteroides fragilis protects against CNS demyelinating disease. Mucosal Immunol. 2010, 3, 487–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefan, K.L.; Kim, M.V.; Iwasaki, A.; Kasper, D.L. Commensal Microbiota Modulation of Natural Resistance to Virus Infection. Cell 2020, 183, 1312–1324.e10. [Google Scholar] [CrossRef]
- An, D.; Oh, S.F.; Olszak, T.; Neves, J.F.; Avci, F.Y.; Erturk-Hasdemir, D.; Lu, X.; Zeissig, S.; Blumberg, R.S.; Kasper, D.L. Sphingolipids from a symbiotic microbe regulate homeostasis of host intestinal natural killer T cells. Cell 2014, 156, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Wieland Brown, L.C.; Penaranda, C.; Kashyap, P.C.; Williams, B.B.; Clardy, J.; Kronenberg, M.; Sonnenburg, J.L.; Comstock, L.E.; Bluestone, J.A.; Fischbach, M.A. Production of α-galactosylceramide by a prominent member of the human gut microbiota. PLoS Biol. 2013, 11, e1001610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, R.; Zuo, F.; Ma, H.; Chen, S. Exopolysaccharide-Producing Bifidobacterium adolescentis Strains with Similar Adhesion Property Induce Differential Regulation of Inflammatory Immune Response in Treg/Th17 Axis of DSS-Colitis Mice. Nutrients 2019, 11, 782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanning, S.; Hall, L.J.; Cronin, M.; Zomer, A.; MacSharry, J.; Goulding, D.; Motherway, M.O.; Shanahan, F.; Nally, K.; Dougan, G.; et al. Bifidobacterial surface-exopolysaccharide facilitates commensal-host interaction through immune modulation and pathogen protection. Proc. Natl. Acad. Sci. USA 2012, 109, 2108–2113. [Google Scholar] [CrossRef] [Green Version]
- Hickey, A.; Stamou, P.; Udayan, S.; Ramón-Vázquez, A.; Esteban-Torres, M.; Bottacini, F.; Woznicki, J.A.; Hughes, O.; Melgar, S.; Ventura, M.; et al. Bifidobacterium breve Exopolysaccharide Blocks Dendritic Cell Maturation and Activation of CD4+ T Cells. Front. Microbiol. 2021, 12, 653587. [Google Scholar] [CrossRef]
- Hughes, K.R.; Harnisch, L.C.; Alcon-Giner, C.; Mitra, S.; Wright, C.J.; Ketskemety, J.; van Sinderen, D.; Watson, A.J.; Hall, L.J. Bifidobacterium breve reduces apoptotic epithelial cell shedding in an exopolysaccharide and MyD88-dependent manner. Open Biol. 2017, 7, 160155. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Gan, M.; Yu, X.; Wu, X.; Xu, F. Study on the regulatory effects and mechanisms of action of bifidobacterial exopolysaccharides on anaphylaxes in mice. Int. J. Biol. Macromol. 2020, 165, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, D.; Emonet, C.; Foata, F.; Affolter, M.; Delley, M.; Fisseha, M.; Blum-Sperisen, S.; Kochhar, S.; Arigoni, F. A serpin from the gut bacterium Bifidobacterium longum inhibits eukaryotic elastase-like serine proteases. J. Biol. Chem. 2006, 281, 17246–17252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarville, J.L.; Dong, J.; Caminero, A.; Bermudez-Brito, M.; Jury, J.; Murray, J.A.; Duboux, S.; Steinmann, M.; Delley, M.; Tangyu, M.; et al. A Commensal Bifidobacterium longum Strain Prevents Gluten-Related Immunopathology in Mice through Expression of a Serine Protease Inhibitor. Appl. Environ. Microbiol. 2017, 83, e01323-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiavi, E.; Gleinser, M.; Molloy, E.; Groeger, D.; Frei, R.; Ferstl, R.; Rodriguez-Perez, N.; Ziegler, M.; Grant, R.; Moriarty, T.F.; et al. The Surface-Associated Exopolysaccharide of Bifidobacterium longum 35624 Plays an Essential Role in Dampening Host Proinflammatory Responses and Repressing Local TH17 Responses. Appl. Environ. Microbiol. 2016, 82, 7185–7196. [Google Scholar] [CrossRef] [Green Version]
- Schiavi, E.; Plattner, S.; Rodriguez-Perez, N.; Barcik, W.; Frei, R.; Ferstl, R.; Kurnik-Lucka, M.; Groeger, D.; Grant, R.; Roper, J.; et al. Exopolysaccharide from Bifidobacterium longum subsp. longum 35624™ modulates murine allergic airway responses. Benef. Microbes 2018, 9, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Yang, B.; Zhao, J.; Zhao, J.; Stanton, C.; Ross, R.P.; Zhang, H.; Chen, W. A ropy exopolysaccharide producing strain Bifidobacterium longum subsp. longum YS108R alleviates DSS-induced colitis by maintenance of the mucosal barrier and gut microbiota modulation. Food Funct. 2019, 10, 1595–1608. [Google Scholar] [CrossRef] [PubMed]
- O’Connell Motherway, M.; Zomer, A.; Leahy, S.C.; Reunanen, J.; Bottacini, F.; Claesson, M.J.; O’Brien, F.; Flynn, K.; Casey, P.G.; Munoz, J.A.; et al. Functional genome analysis of Bifidobacterium breve UCC2003 reveals type IVb tight adherence (Tad) pili as an essential and conserved host-colonization factor. Proc. Natl. Acad. Sci. USA 2011, 108, 11217–11222. [Google Scholar] [CrossRef] [Green Version]
- Vargas García, C.E.; Petrova, M.; Claes, I.J.; De Boeck, I.; Verhoeven, T.L.; Dilissen, E.; von Ossowski, I.; Palva, A.; Bullens, D.M.; Vanderleyden, J.; et al. Piliation of Lactobacillus rhamnosus GG promotes adhesion, phagocytosis, and cytokine modulation in macrophages. Appl. Environ. Microbiol. 2015, 81, 2050–2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turroni, F.; Serafini, F.; Foroni, E.; Duranti, S.; O’Connell Motherway, M.; Taverniti, V.; Mangifesta, M.; Milani, C.; Viappiani, A.; Roversi, T.; et al. Role of sortase-dependent pili of Bifidobacterium bifidum PRL2010 in modulating bacterium-host interactions. Proc. Natl. Acad. Sci. USA 2013, 110, 11151–11156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlee, M.; Wehkamp, J.; Altenhoefer, A.; Oelschlaeger, T.A.; Stange, E.F.; Fellermann, K. Induction of human beta-defensin 2 by the probiotic Escherichia coli Nissle 1917 is mediated through flagellin. Infect. Immun. 2007, 75, 2399–2407. [Google Scholar] [CrossRef] [Green Version]
- Konstantinov, S.R.; Smidt, H.; de Vos, W.M.; Bruijns, S.C.; Singh, S.K.; Valence, F.; Molle, D.; Lortal, S.; Altermann, E.; Klaenhammer, T.R.; et al. S layer protein A of Lactobacillus acidophilus NCFM regulates immature dendritic cell and T cell functions. Proc. Natl. Acad. Sci. USA 2008, 105, 19474–19479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Schillde, M.A.; Hörmannsperger, G.; Weiher, M.; Alpert, C.A.; Hahne, H.; Bäuerl, C.; van Huynegem, K.; Steidler, L.; Hrncir, T.; Pérez-Martínez, G.; et al. Lactocepin secreted by Lactobacillus exerts anti-inflammatory effects by selectively degrading proinflammatory chemokines. Cell Host Microbe 2012, 11, 387–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seth, A.; Yan, F.; Polk, D.B.; Rao, R.K. Probiotics ameliorate the hydrogen peroxide-induced epithelial barrier disruption by a PKC- and MAP kinase-dependent mechanism. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1060–G1069. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Cao, H.; Cover, T.L.; Washington, M.K.; Shi, Y.; Liu, L.; Chaturvedi, R.; Peek, R.M., Jr.; Wilson, K.T.; Polk, D.B. Colon-specific delivery of a probiotic-derived soluble protein ameliorates intestinal inflammation in mice through an EGFR-dependent mechanism. J. Clin. Investig. 2011, 121, 2242–2253. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Cao, H.; Cover, T.L.; Whitehead, R.; Washington, M.K.; Polk, D.B. Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterology 2007, 132, 562–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasuda, E.; Serata, M.; Sako, T. Suppressive effect on activation of macrophages by Lactobacillus casei strain Shirota genes determining the synthesis of cell wall-associated polysaccharides. Appl. Environ. Microbiol. 2008, 74, 4746–4755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, S.; Hara, T.; Hori, T.; Mitsuyama, K.; Nagaoka, M.; Tomiyasu, N.; Suzuki, A.; Sata, M. Probiotic Lactobacillus-induced improvement in murine chronic inflammatory bowel disease is associated with the down-regulation of pro-inflammatory cytokines in lamina propria mononuclear cells. Clin. Exp. Immunol. 2005, 140, 417–426. [Google Scholar] [CrossRef]
- Liu, Y.; Zheng, S.; Cui, J.; Guo, T.; Zhang, J.; Li, B. Alleviative Effects of Exopolysaccharide Produced by Lactobacillus helveticus KLDS1.8701 on Dextran Sulfate Sodium-Induced Colitis in Mice. Microorganisms 2021, 9, 2086. [Google Scholar] [CrossRef] [PubMed]
- Vinderola, G.; Perdigón, G.; Duarte, J.; Farnworth, E.; Matar, C. Effects of the oral administration of the exopolysaccharide produced by Lactobacillus kefiranofaciens on the gut mucosal immunity. Cytokine 2006, 36, 254–260. [Google Scholar] [CrossRef]
- Al-Hassi, H.O.; Mann, E.R.; Sanchez, B.; English, N.R.; Peake, S.T.; Landy, J.; Man, R.; Urdaci, M.; Hart, A.L.; Fernandez-Salazar, L.; et al. Altered human gut dendritic cell properties in ulcerative colitis are reversed by Lactobacillus plantarum extracellular encrypted peptide STp. Mol. Nutr. Food Res. 2014, 58, 1132–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Lee, Y.D.; Kim, M.; Kim, H.; Chung, D.K. Combination treatment with lipoteichoic acids isolated from Lactobacillus plantarum and Staphylococcus aureus alleviates atopic dermatitis via upregulation of CD55 and CD59. Immunol. Lett. 2019, 214, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.E.; Kim, H.; Chung, D.K. Lipoteichoic Acid Isolated from Lactobacillus plantarum Maintains Inflammatory Homeostasis through Regulation of Th1- and Th2-Induced Cytokines. J. Microbiol. Biotechnol. 2019, 29, 151–159. [Google Scholar] [CrossRef]
- Jeon, B.; Kim, H.R.; Kim, H.; Chung, D.K. In vitro and in vivo downregulation of C3 by lipoteichoic acid isolated from Lactobacillus plantarum K8 suppressed cytokine-mediated complement system activation. FEMS Microbiol. Lett. 2016, 363, fnw140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, M.; Lee, J.; Park, S.; Kwon, O.H.; Seo, J.; Roh, S. Exopolysaccharide Isolated from Lactobacillus plantarum L-14 Has Anti-Inflammatory Effects via the Toll-Like Receptor 4 Pathway in LPS-Induced RAW 264.7 Cells. Int. J. Mol. Sci. 2020, 21, 9283. [Google Scholar] [CrossRef]
- Murofushi, Y.; Villena, J.; Morie, K.; Kanmani, P.; Tohno, M.; Shimazu, T.; Aso, H.; Suda, Y.; Hashiguchi, K.; Saito, T.; et al. The toll-like receptor family protein RP105/MD1 complex is involved in the immunoregulatory effect of exopolysaccharides from Lactobacillus plantarum N14. Mol. Immunol. 2015, 64, 63–75. [Google Scholar] [CrossRef]
- Lebeer, S.; Claes, I.; Tytgat, H.L.; Verhoeven, T.L.; Marien, E.; von Ossowski, I.; Reunanen, J.; Palva, A.; Vos, W.M.; Keersmaecker, S.C.; et al. Functional analysis of Lactobacillus rhamnosus GG pili in relation to adhesion and immunomodulatory interactions with intestinal epithelial cells. Appl. Environ. Microbiol. 2012, 78, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claes, I.J.; Lebeer, S.; Shen, C.; Verhoeven, T.L.; Dilissen, E.; De Hertogh, G.; Bullens, D.M.; Ceuppens, J.L.; Van Assche, G.; Vermeire, S.; et al. Impact of lipoteichoic acid modification on the performance of the probiotic Lactobacillus rhamnosus GG in experimental colitis. Clin. Exp. Immunol. 2010, 162, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Harb, H.; van Tol, E.A.; Heine, H.; Braaksma, M.; Gross, G.; Overkamp, K.; Hennen, M.; Alrifai, M.; Conrad, M.L.; Renz, H.; et al. Neonatal supplementation of processed supernatant from Lactobacillus rhamnosus GG improves allergic airway inflammation in mice later in life. Clin. Exp. Allergy 2013, 43, 353–364. [Google Scholar] [CrossRef] [PubMed]
- You, G.E.; Jung, B.J.; Kim, H.R.; Kim, H.G.; Kim, T.R.; Chung, D.K. Lactobacillus sakei lipoteichoic acid inhibits MMP-1 induced by UVA in normal dermal fibroblasts of human. J. Microbiol. Biotechnol. 2013, 23, 1357–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macho Fernandez, E.; Valenti, V.; Rockel, C.; Hermann, C.; Pot, B.; Boneca, I.G.; Grangette, C. Anti-inflammatory capacity of selected lactobacilli in experimental colitis is driven by NOD2-mediated recognition of a specific peptidoglycan-derived muropeptide. Gut 2011, 60, 1050–1059. [Google Scholar] [CrossRef]
- Kaji, R.; Kiyoshima-Shibata, J.; Nagaoka, M.; Nanno, M.; Shida, K. Bacterial teichoic acids reverse predominant IL-12 production induced by certain lactobacillus strains into predominant IL-10 production via TLR2-dependent ERK activation in macrophages. J. Immunol. 2010, 184, 3505–3513. [Google Scholar] [CrossRef] [Green Version]
- Dinić, M.; Pecikoza, U.; Djokić, J.; Stepanović-Petrović, R.; Milenković, M.; Stevanović, M.; Filipović, N.; Begović, J.; Golić, N.; Lukić, J. Exopolysaccharide Produced by Probiotic Strain Lactobacillus paraplantarum BGCG11 Reduces Inflammatory Hyperalgesia in Rats. Front. Pharmacol. 2018, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Qi, W.; Hong, T.; Xiong, T.; Gong, D.; Xie, M.; Nie, S. Exopolysaccharides from Lactobacillus plantarum NCU116 Regulate Intestinal Barrier Function via STAT3 Signaling Pathway. J. Agric. Food Chem. 2018, 66, 9719–9727. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Q.; Gao, N.; Wang, Z.; Li, F.; Li, J.; Shan, A. Exopolysaccharides produced by Lactobacillus rhamnosus GG alleviate hydrogen peroxide-induced intestinal oxidative damage and apoptosis through the Keap1/Nrf2 and Bax/Bcl-2 pathways in vitro. Food Funct. 2021, 12, 9632–9641. [Google Scholar] [CrossRef]
- Riehl, T.E.; Alvarado, D.; Ee, X.; Zuckerman, A.; Foster, L.; Kapoor, V.; Thotala, D.; Ciorba, M.A.; Stenson, W.F. Lactobacillus rhamnosus GG protects the intestinal epithelium from radiation injury through release of lipoteichoic acid, macrophage activation and the migration of mesenchymal stem cells. Gut 2019, 68, 1003–1013. [Google Scholar] [CrossRef]
- Nowak, B.; Śróttek, M.; Ciszek-Lenda, M.; Skałkowska, A.; Gamian, A.; Górska, S.; Marcinkiewicz, J. Exopolysaccharide from Lactobacillus rhamnosus KL37 Inhibits T Cell-dependent Immune Response in Mice. Arch. Immunol. Ther. Exp. 2020, 68, 17. [Google Scholar] [CrossRef] [PubMed]
- Le Maréchal, C.; Peton, V.; Plé, C.; Vroland, C.; Jardin, J.; Briard-Bion, V.; Durant, G.; Chuat, V.; Loux, V.; Foligné, B.; et al. Surface proteins of Propionibacterium freudenreichii are involved in its anti-inflammatory properties. J. Proteom. 2015, 113, 447–461. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Liu, C.H.; Tzianabos, A.O.; Kasper, D.L. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, S.; Hara, T.; Nagaoka, M.; Mike, A.; Mitsuyama, K.; Sako, T.; Yamamoto, M.; Kado, S.; Takada, T. A component of polysaccharide peptidoglycan complex on Lactobacillus induced an improvement of murine model of inflammatory bowel disease and colitis-associated cancer. Immunology 2009, 128, e170–e180. [Google Scholar] [CrossRef] [PubMed]
- Laiño, J.; Villena, J.; Kanmani, P.; Kitazawa, H. Immunoregulatory Effects Triggered by Lactic Acid Bacteria Exopolysaccharides: New Insights into Molecular Interactions with Host Cells. Microorganisms 2016, 4, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Divanovic, S.; Trompette, A.; Atabani, S.F.; Madan, R.; Golenbock, D.T.; Visintin, A.; Finberg, R.W.; Tarakhovsky, A.; Vogel, S.N.; Belkaid, Y.; et al. Inhibition of TLR-4/MD-2 signaling by RP105/MD-1. J. Endotoxin Res. 2005, 11, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Kearns, D.B.; Chu, F.; Branda, S.S.; Kolter, R.; Losick, R. A master regulator for biofilm formation by Bacillus subtilis. Mol. Microbiol. 2005, 55, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Flannagan, R.S.; Heit, B.; Heinrichs, D.E. Antimicrobial Mechanisms of Macrophages and the Immune Evasion Strategies of Staphylococcus aureus. Pathogens 2015, 4, 826–868. [Google Scholar] [CrossRef] [Green Version]
- Strachan, D.P. Hay fever, hygiene, and household size. BMJ 1989, 299, 1259–1260. [Google Scholar] [CrossRef] [Green Version]
- Stein, M.M.; Hrusch, C.L.; Gozdz, J.; Igartua, C.; Pivniouk, V.; Murray, S.E.; Ledford, J.G.; Marques Dos Santos, M.; Anderson, R.L.; Metwali, N.; et al. Innate Immunity and Asthma Risk in Amish and Hutterite Farm Children. N. Engl. J. Med. 2016, 375, 411–421. [Google Scholar] [CrossRef] [Green Version]
- Ober, C.; Sperling, A.I.; von Mutius, E.; Vercelli, D. Immune development and environment: Lessons from Amish and Hutterite children. Curr. Opin. Immunol. 2017, 48, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Okudaira, H.; Nogami, M.; Matsuzaki, G.; Dohi, M.; Suko, M.; Kasuya, S.; Takatsu, K. T-cell-dependent accumulation of eosinophils in the lung and its inhibition by monoclonal anti-interleukin-5. Int. Arch. Allergy Appl. Immunol. 1991, 94, 171–173. [Google Scholar] [CrossRef]
- Biswas, S.K.; Lopez-Collazo, E. Endotoxin tolerance: New mechanisms, molecules and clinical significance. Trends Immunol. 2009, 30, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Seeley, J.J.; Ghosh, S. Molecular mechanisms of innate memory and tolerance to LPS. J. Leukoc. Biol. 2017, 101, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Horvatinovich, J.M.; Grogan, E.W.; Norris, M.; Steinkasserer, A.; Lemos, H.; Mellor, A.L.; Tcherepanova, I.Y.; Nicolette, C.A.; DeBenedette, M.A. Soluble CD83 Inhibits T Cell Activation by Binding to the TLR4/MD-2 Complex on CD14+ Monocytes. J. Immunol. 2017, 198, 2286–2301. [Google Scholar] [CrossRef] [Green Version]
- Gringhuis, S.I.; den Dunnen, J.; Litjens, M.; van Het Hof, B.; van Kooyk, Y.; Geijtenbeek, T.B. C-type lectin DC-SIGN modulates Toll-like receptor signaling via Raf-1 kinase-dependent acetylation of transcription factor NF-kappaB. Immunity 2007, 26, 605–616. [Google Scholar] [CrossRef] [Green Version]
- Yao, X.; Dong, G.; Zhu, Y.; Yan, F.; Zhang, H.; Ma, Q.; Fu, X.; Li, X.; Zhang, Q.; Zhang, J.; et al. Leukadherin-1-Mediated Activation of CD11b Inhibits LPS-Induced Pro-inflammatory Response in Macrophages and Protects Mice Against Endotoxic Shock by Blocking LPS-TLR4 Interaction. Front. Immunol. 2019, 10, 215. [Google Scholar] [CrossRef]
- Li, Y.; Komai-Koma, M.; Gilchrist, D.S.; Hsu, D.K.; Liu, F.T.; Springall, T.; Xu, D. Galectin-3 is a negative regulator of lipopolysaccharide-mediated inflammation. J. Immunol. 2008, 181, 2781–2789. [Google Scholar] [CrossRef] [Green Version]
- Terness, P.; Bauer, T.M.; Röse, L.; Dufter, C.; Watzlik, A.; Simon, H.; Opelz, G. Inhibition of allogeneic T cell proliferation by indoleamine 2,3-dioxygenase-expressing dendritic cells: Mediation of suppression by tryptophan metabolites. J. Exp. Med. 2002, 196, 447–457. [Google Scholar] [CrossRef] [Green Version]
- Bressler, B.; Marshall, J.K.; Bernstein, C.N.; Bitton, A.; Jones, J.; Leontiadis, G.I.; Panaccione, R.; Steinhart, A.H.; Tse, F.; Feagan, B. Clinical practice guidelines for the medical management of nonhospitalized ulcerative colitis: The Toronto consensus. Gastroenterology 2015, 148, 1035–1058.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Organism | Molecule | Results | Reference |
|---|---|---|---|
| Bacillus subtilis | Lipoteichoic acid (LTA) EPSBs | LTA activates a TLR2-dependent inflammatory response and concomitantly induces activation of MerTK signaling to counteract the inflammation in vitro. EPSBs reduces Citrobacter rodentium infection and generates peritoneal anti-inflammatory macrophages. EPSBs prevents allergic eosinophilia. EPSBs ameliorates GvHD and can generate tolerogenic DCs in vitro. EPSBs protects against systemic infection of Staphylococcus aureus. | [14] [9] [10,12] [8] [7] [11,13] |
| Bacteroides fragilis | Polysaccharide A (PSA) Glycosphingolipids | PSA is protective and therapeutic in murine models of colitis and multiple sclerosis via the induction of IL-10 secreting Tregs. PSA activates colonic DCs and produces IFN-β that enhances resistance to viral infection in murine models. This protection is dependent on TLR4. Glycosphingolipids decrease the number of invariant natural killer T cells in the colonic lamina propria leading to improved outcomes in a murine colitis model. | [15,16,17,18,19] [20] [21,22] |
| Bifidobacterium adolescentis | EPSBa | EPSBa induces IL-10 production, protects from colitis by activation of DCs and macrophages, and increases the Treg/Th17 cell ratio in mice. | [23] |
| Bifidobacterium breve UCC2003 | EPSBb | EPSBb reduces Citrobacter rodentium infection in mice. EPSBb prevents the maturation of DCs and activation of antigen-specific CD4+ T cells. EPSBb reduces the rate of small epithelial cell shedding in a mouse model of pathological cell shedding. | [24] [25] [26] |
| Bifidobacterium breve WBBR04 | EPSBb | EPSBb enhances the intestinal barrier integrity to prevent allergen infiltration and food allergy in mice. | [27] |
| Bifidobacterium longum | Serpin | Serpin inhibits eukaryotic elastase-like serine proteases, which are dysregulated in inflammatory disorders. | [28,29] |
| Bifidobacterium Longum 35624 | EPSBl | EPSBl dampens pro-inflammatory cytokines and reduces inflammatory symptoms in a T cell transfer colitis model. EPSBl stimulates the release of IL-10 in a TLR2-dependent manner and reduces eosinophil recruitment in the lungs in a respiratory inflammation mouse model. | [30] [31] |
| Bifidobacterium Longum YS108R | EPSBl | EPSBl reduces the pro-inflammatory cytokines IL-6 and IL-17A, alleviating inflammation in a colitis murine model. | [32] |
| Bifidobacterium sp. | Fimbriae | Fimbriae facilitate gut colonization and stimulation of macrophage cytokine production, TNF-α, IL-6, and Il-10. | [33,34,35] |
| Escherichia coli Nissle 1917 | Flagellin | Flagellin induces the release of β-defensin-2 in epithelial cells through NF-κB- and AP-1-dependent pathways in vitro. | [36] |
| Lactobacillus acidophilus NCFM | Surface layer protein A (SlpA) | SlpA binds the lectin-receptor DC-SIGN and increases IL-10 and reduces IL-12p70 production from DCs. | [37] |
| Lactobacillus casei | Lactoceptin | Lactoceptin can selectively hydrolyze pro-inflammatory chemokine IP-10 leading to reduced lymphocyte recruitment in an ileitis murine model. | [38] |
| Lactobacillus casei BL23 | p40 and p75 | Secreted proteins p40 and p75 stimulate Akt activation, display anti-apoptotic activity, and prevent epithelial barrier damage in colitis murine models. | [39,40,41] |
| Lactobacillus casei Shirota | High molecular components of cell wall Polysaccharide peptidoglycan complex | High molecular weight cell wall components of Lactobacillus casei Shirota decrease LPS-induced IL-6 production in macrophages. Polysaccharide peptidoglycan complex improves ileitis and inhibits IL6/STAT3 signaling in a murine colitis model. | [42] [43] |
| Lactobacillus helveticus KLD1.8701 | EPSLh | EPSLh reduces intestinal inflammation and improves mucosal barrier function in a murine colitis model. | [44] |
| Lactobacillus kefiranofaciens | EPSLk | EPSLk increases the number of IgA+ cells in the small and large intestines and increases the levels of IL-4 and IL-12 in the intestinal fluid and serum. | [45] |
| Lactobacillus plantarum | Serine-threonine peptide (STp) | STp changes the phenotype of DC from ulcerative colitis patients by reducing TLR expression, increasing activation markers, and restoring stimulatory capacity. | [46] |
| Lactobacillus plantarum | LTA | LTA from L. plantarum and Staphylococcus aureus alleviates atopic dermatitis by regulating the complement regulatory proteins CD55 and CD59 and reducing activation of the complement system. LTA inhibits the release of TNF-α and IL-10 from stimulated THP-1 cells by dephosphorylating c-Jun N-terminal kinase (JNK) and p38, respectively. | [47] [48] |
| Lactobacillus plantarum K8 | LTA | LTA suppresses inflammatory cytokine-mediated complement activation through the inhibition of C3 synthesis. | [49] |
| Lactobacillus plantarum L-14 | EPSLp | EPSLp suppresses the pro-inflammatory cytokine mediators, COX-2, IL-6, TNF-α, and IL-1β induced by LPS. | [50] |
| Lactobacillus plantarum N-14 | EPSLp | EPSLp activates RP105/MD1 on intestinal epithelial cells to reduce inflammatory pathways. | [51] |
| Lactobacillus rhamnosus GG | Pili | Pili helps the adhesion of Lactobacillus rhamnosus GG to epithelial and the release of anti-inflammatory IL-10, IL-8, and IL-6 from epithelial cells. | [34,52] |
| Lactobacillus rhamnosus GG | Lipoteichoic acid (LTA) Supernatants | LTA improves colitis in a murine model. Administration of culture supernatants reduces eosinophil numbers, goblet cells, and lung inflammation in murine allergy model. | [53,54] |
| Lactobacillus sakei | LTA | LTA inhibits the secretion of TNF-α from UVA-exposed derma fibroblasts. | [55] |
| Lactobacillus salivarius Ls33 | Peptidoglycan | Peptidoglycan protects mice from chemically induced colitis in a NOD2-IL-10-dependent manner. | [56] |
| Lactobacillus Sp | Teichonic acids | Teichonic acid induces IL-10 in a TLR2-dependent manner in macrophages. | [57] |
| Lactobacillus paraplantarum BGCG11 | EPSLp | EPSLp reduces pro-inflammatory cytokines in a hyperalgesia rat model that results in anti-hyperalgesic and anti-edematous outcomes. | [58] |
| Lactobacillus plantarum NCU116 | EPSLp | EPSLp regulates the tight junction proteins occluding and ZO-1 by activating STAT3. | [59] |
| Lactobacillus rhamnosus GG | EPSLr LTA | EPSLr reduces hydrogen peroxide-induced intestinal oxidative damage and apoptosis by Keap1/Nrf2 and Bax/Bcl-2 pathways in vitro. LTA protects intestinal epithelial cells from radiation injury through the activation of pericryptal macrophages. These macrophages release CXCL12 that binds to CXCR4 on COX-2 expressing mesenchymal stem cells and stimulate the release of PGE, which protects epithelial stem cells from radiation. | [60] [61] |
| Lactobacillus rhamnosus KL37 | EPSLr | EPSLr inhibits T cell-dependent immune response reducing the arthritogenic antibodies in an arthritis murine model. | [62] |
| Propionibacterium freudenreichii | Guanidine surface protein extract | Treatment of human peripheral blood with guanidine surface protein extract releases IL-10 and IL-6, while having no effect on IL-12, TNF-α, and IFNγ. | [63] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zamora-Pineda, J.; Kalinina, O.; Osborne, B.A.; Knight, K.L. Probiotic Molecules That Inhibit Inflammatory Diseases. Appl. Sci. 2022, 12, 1147. https://doi.org/10.3390/app12031147
Zamora-Pineda J, Kalinina O, Osborne BA, Knight KL. Probiotic Molecules That Inhibit Inflammatory Diseases. Applied Sciences. 2022; 12(3):1147. https://doi.org/10.3390/app12031147
Chicago/Turabian StyleZamora-Pineda, Jesus, Olga Kalinina, Barbara A. Osborne, and Katherine L. Knight. 2022. "Probiotic Molecules That Inhibit Inflammatory Diseases" Applied Sciences 12, no. 3: 1147. https://doi.org/10.3390/app12031147
APA StyleZamora-Pineda, J., Kalinina, O., Osborne, B. A., & Knight, K. L. (2022). Probiotic Molecules That Inhibit Inflammatory Diseases. Applied Sciences, 12(3), 1147. https://doi.org/10.3390/app12031147