
Protective Effect of Ectoin on UVA/H2O2-Induced Oxidative Damage in Human Skin Fibroblast Cells

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
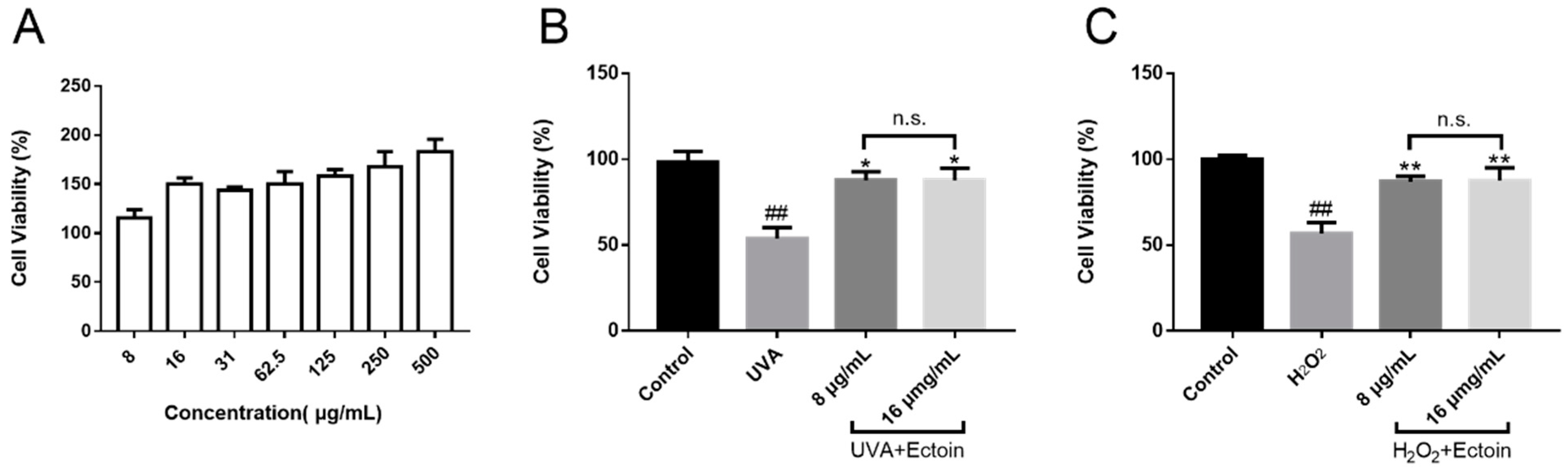
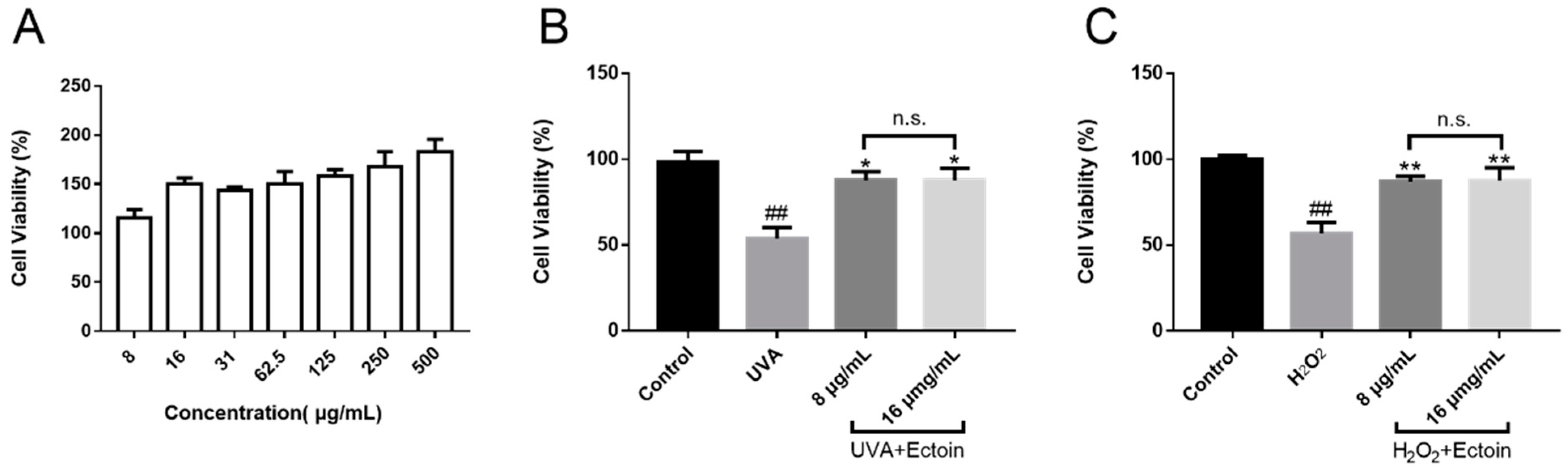
2.3. Cell Viability
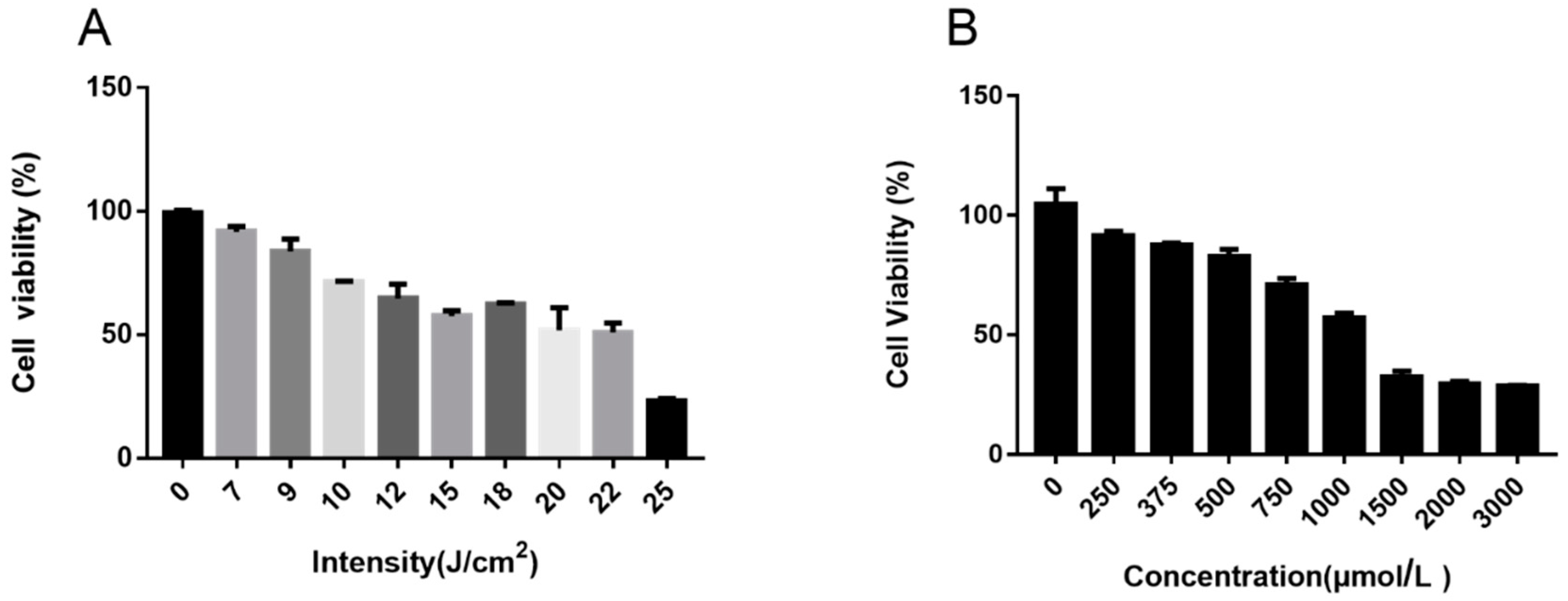
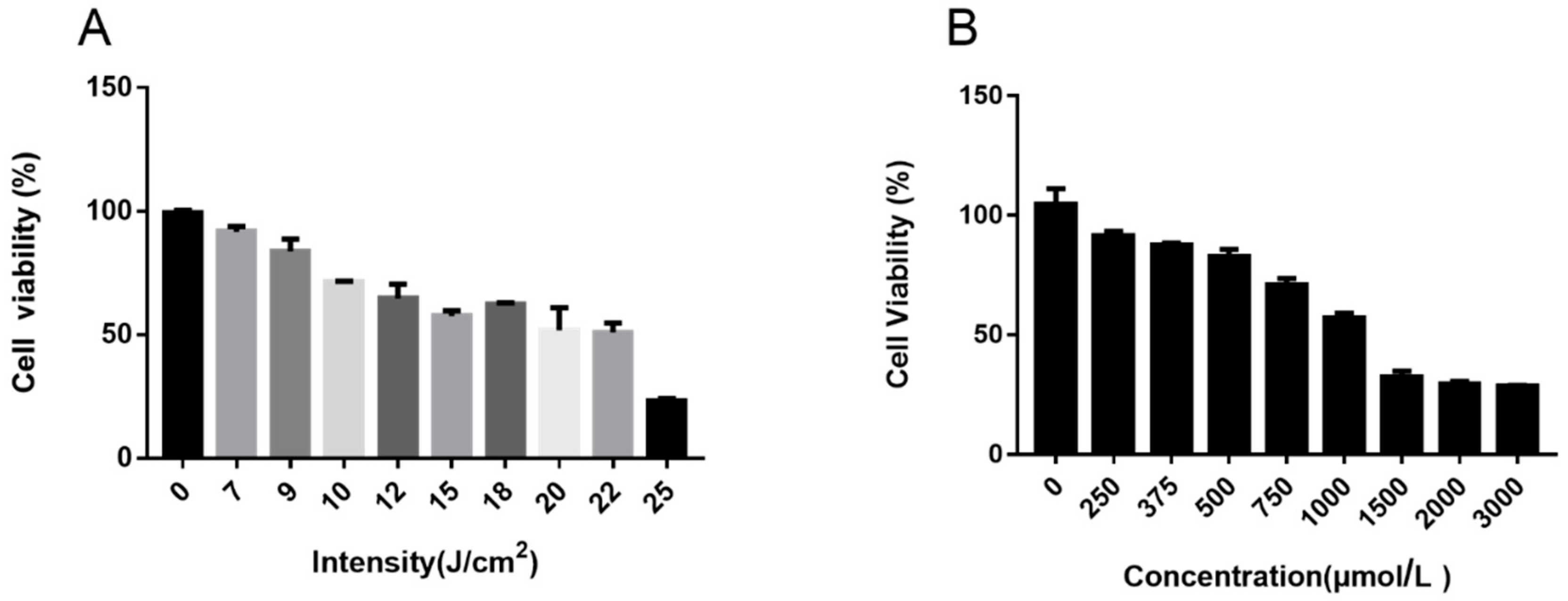
2.4. UVA and H2O2-Induced Model Establishment
2.5. Protective Effects of Ectoin on UVA and H2O2-Induced Model
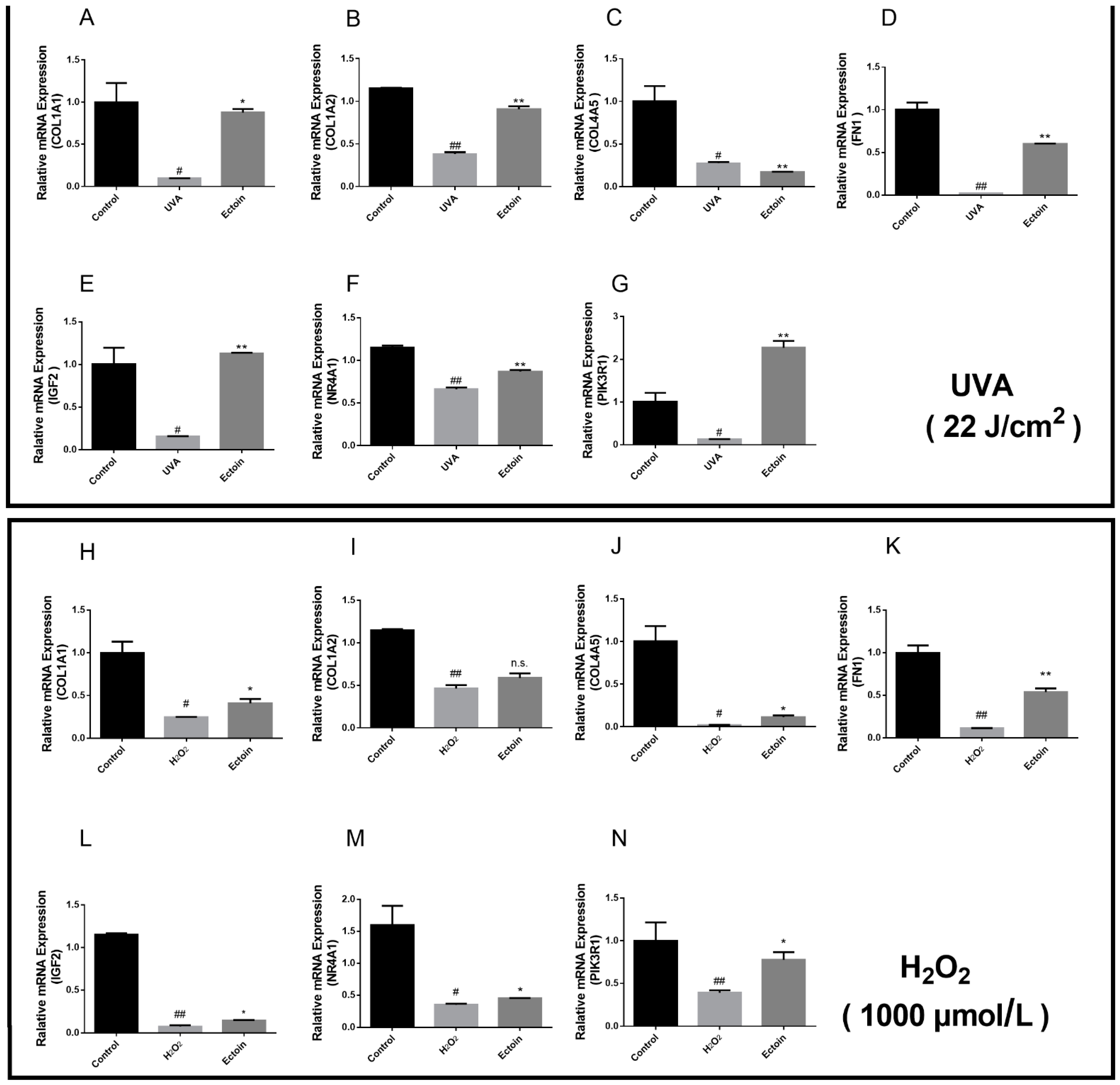
2.6. RT-qPCR
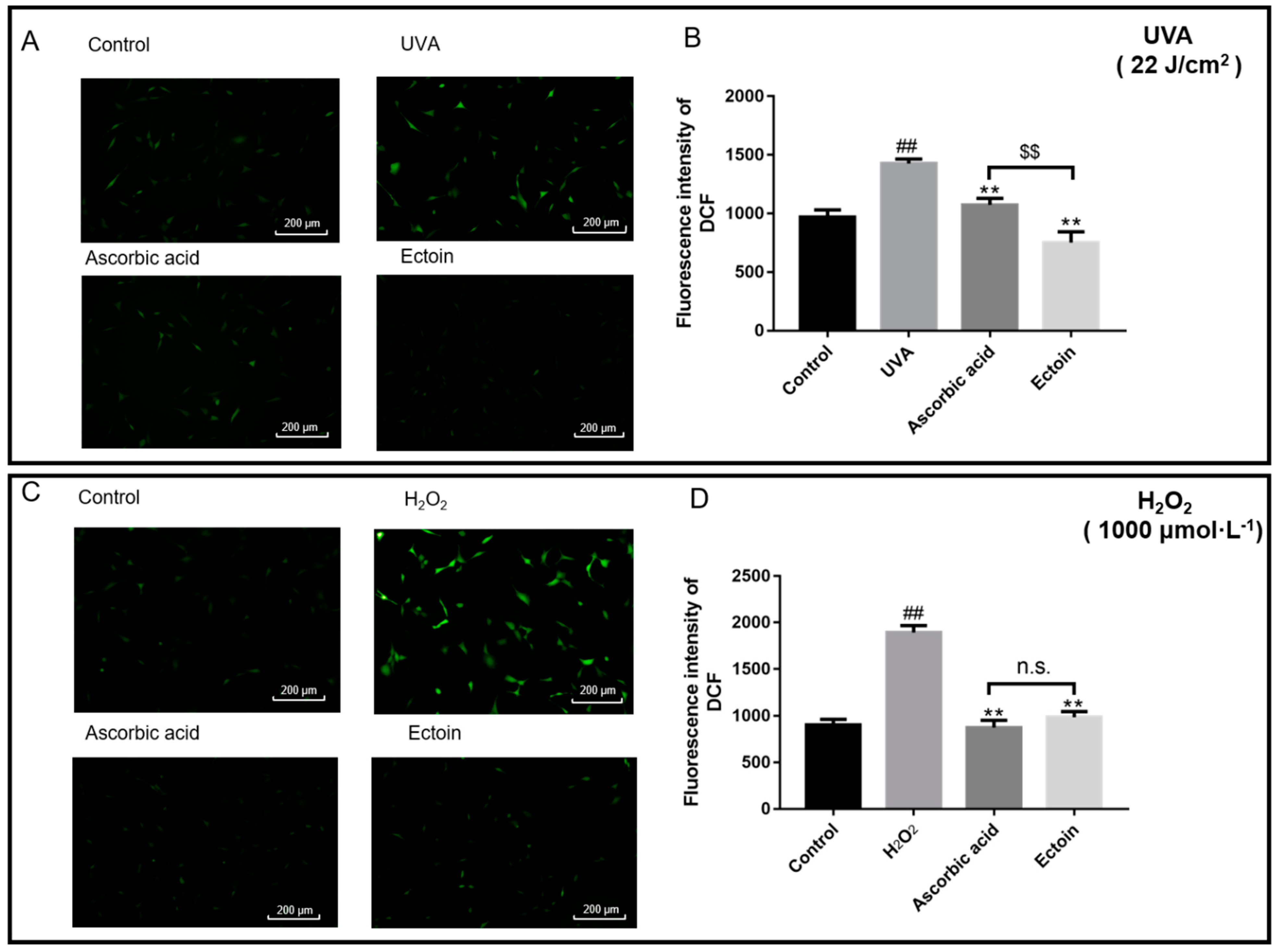
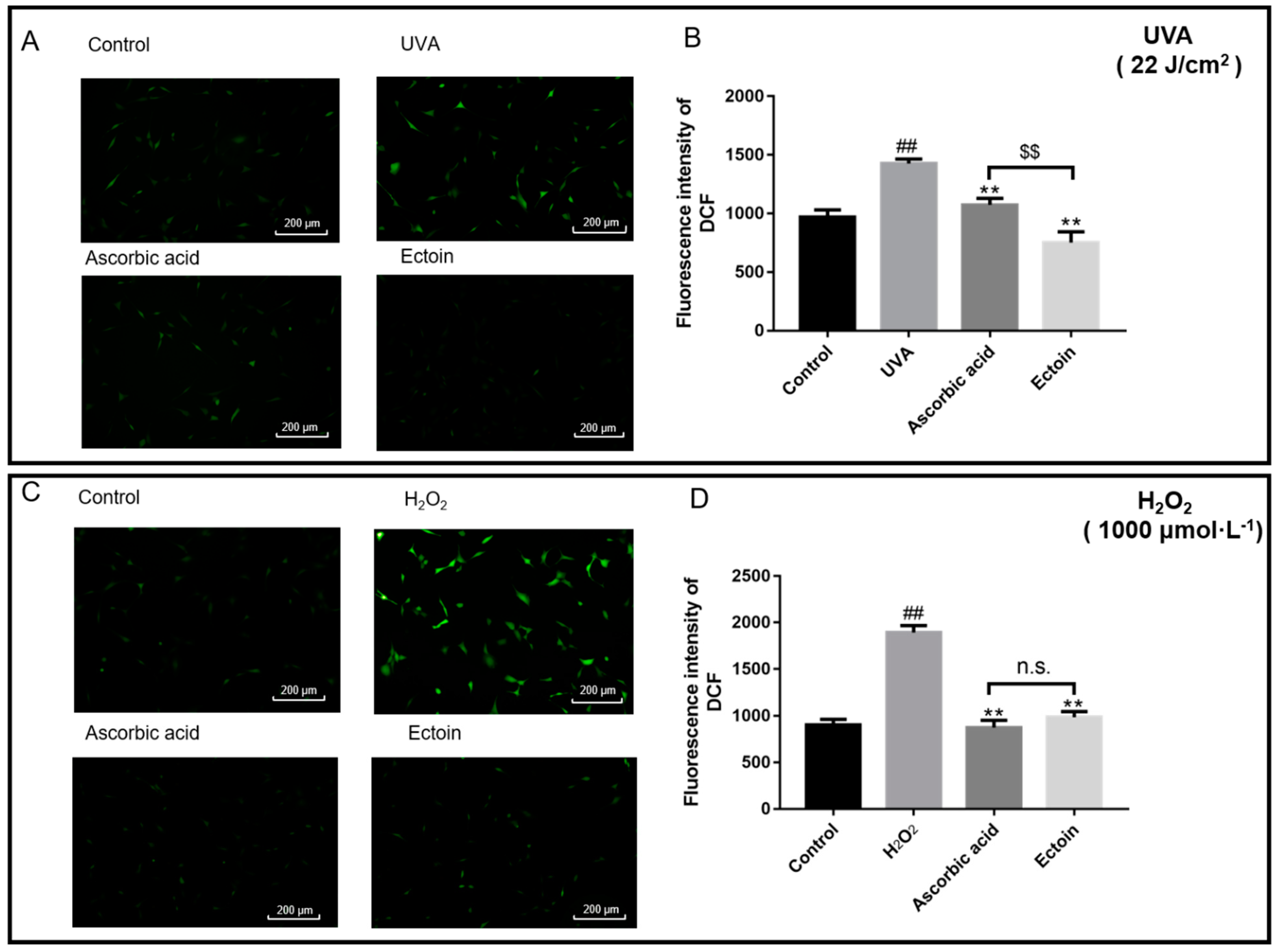
2.7. Determination of Intracellular Reactive Oxygen Species (ROS)
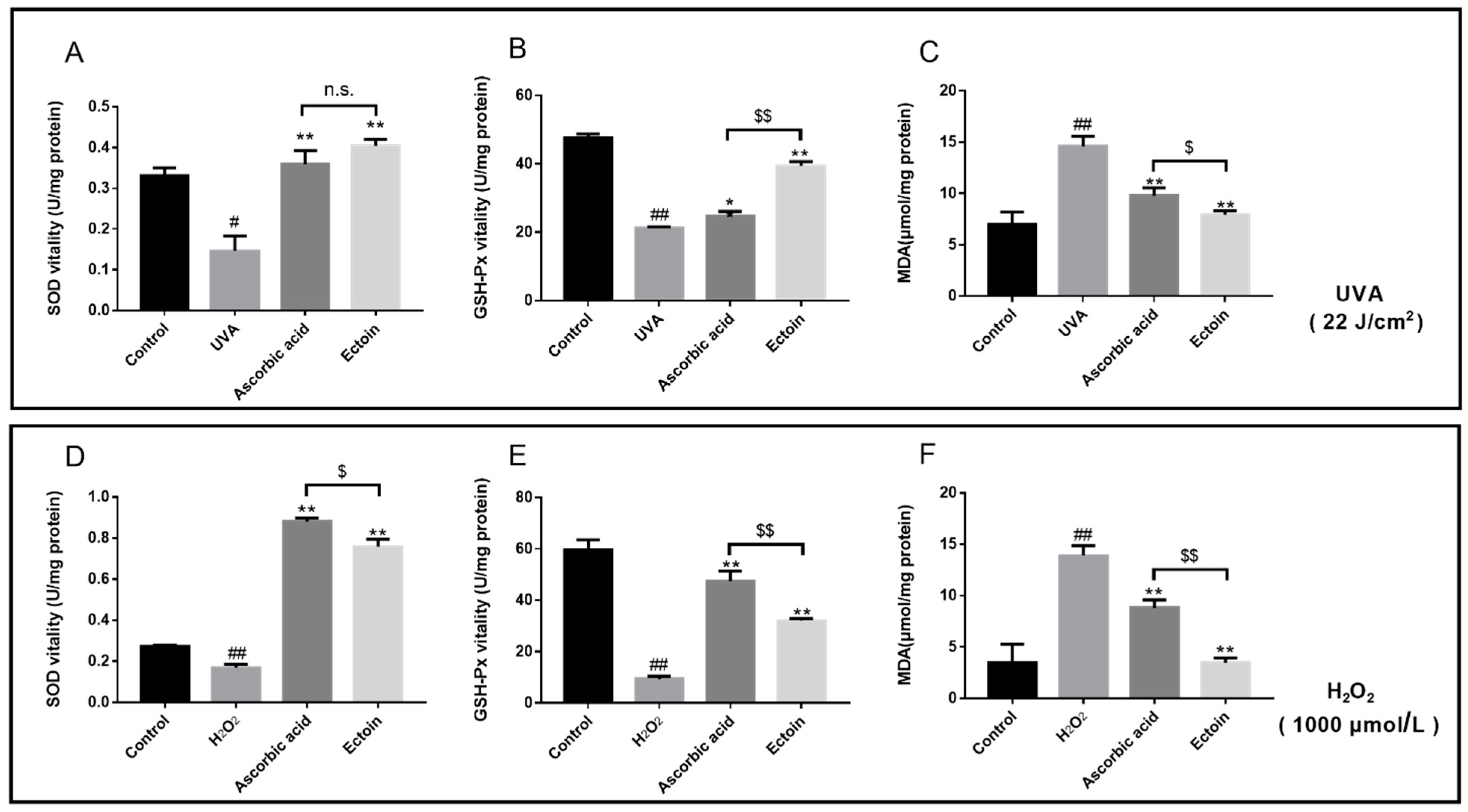
2.8. Determination of Intracellular GSH-Px, MDA, and SOD
2.9. Statistics
3. Results
3.1. Cell Viability of Ectoin and Oxidative Stress Damaged Cells
3.2. Antioxidant Enzymes and ROS Levels of Oxidative Stress Damaged Cells
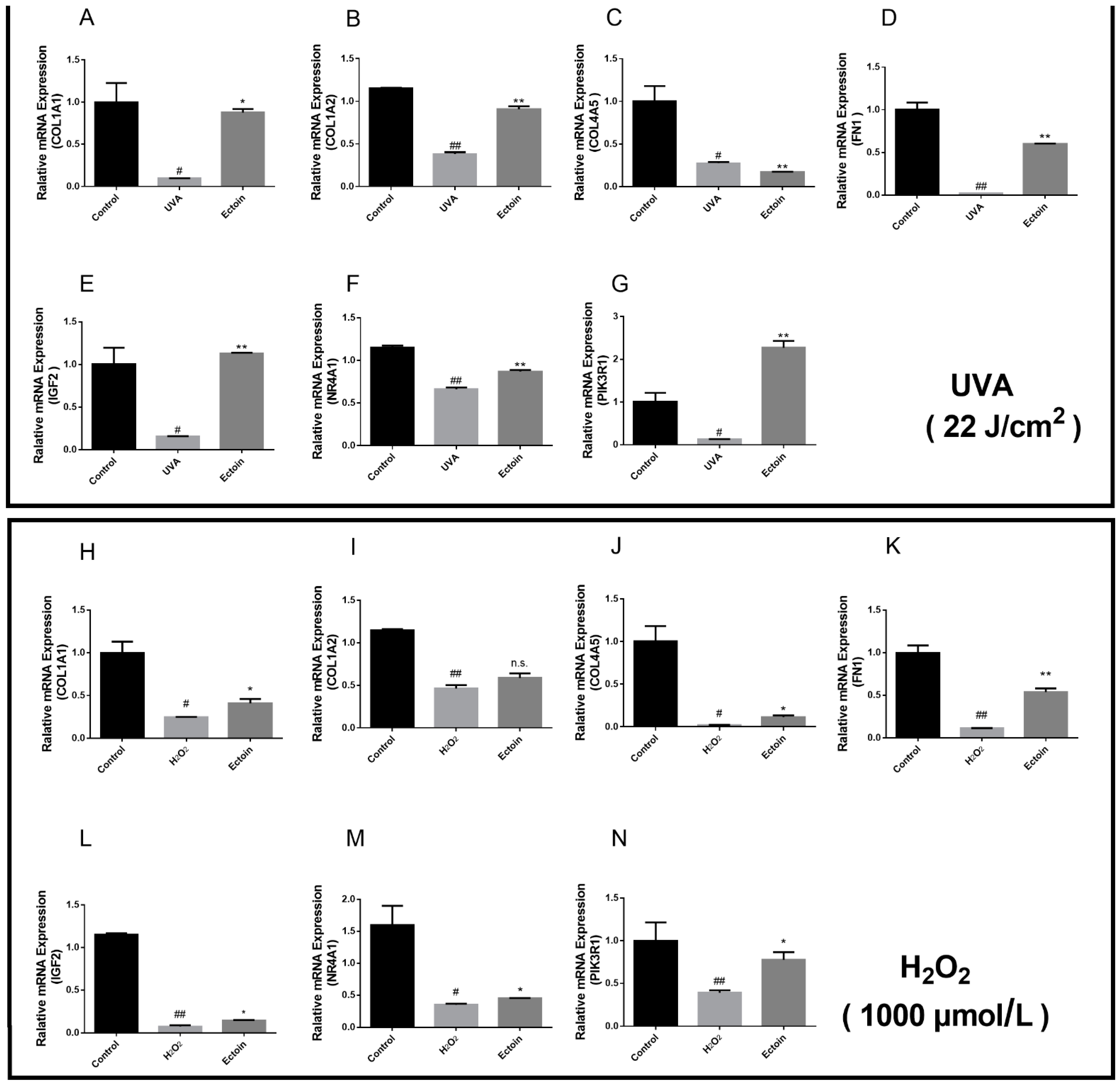
3.3. The Effects of Ectoin on the Expression of Related Genes in the PI3K/AKT Signaling Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vistejnova, L.; Safrankova, B.; Nesporova, K.; Slavkovsky, R.; Hermannova, M.; Hosek, P.; Velebny, V.; Kubala, L. Low Molecular Weight Hyaluronan Mediated CD44 Dependent Induction of IL-6 and Chemokines in Human Dermal Fibroblasts Potentiates Innate Immune Response. Cytokine 2014, 70, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.D.; Watt, F.M. Fibroblast heterogeneity: Implications for human disease. J. Clin. Investig. 2018, 128, 26–35. [Google Scholar] [CrossRef]
- Driskell, R.R.; Watt, F.M. Understanding fibroblast heterogeneity in the skin. Trends Cell Biol. 2015, 25, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, L. Fibroblasts and skin aging. Chin. J. Aesthetic Med. 2005, 14, 243–245. [Google Scholar]
- Yim, J.H.; Jang, M.S.; Mi, Y.M.; Lee, H.; Kim, S.; Lee, N. Constituents from the branches of Sambucus sieboldiana var. pendula with the properties of collagen synthesis activation. J. Appl. Pharm. Sci. 2015, 5, 119–122. [Google Scholar] [CrossRef]
- Gao, W.; Wang, Y.S.; Hwang, E.; Lin, P.; Bae, J.; Seo, S.A.; Yan, Z.; Yi, T.-H. Rubus idaeus. L. (red raspberry) blocks UVB-induced MMP production and promotes type I procollagen synthesis via inhibition of MAPK/AP-1, NF-κβ and stimulation of TGF-β/Smad, Nrf2 in normal human dermal fibroblasts. J. Photochem. Photobiol. B Biol. 2018, 185, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-Y.; Lin, Y.-T.; Kuo, H.-C.; Chiou, W.-F.; Lee, M.-H. Compounds isolated from Eriobotrya deflexa leaves protect against ultraviolet radiation B-induced photoaging in human fibroblasts. J. Photochem. Photobiol. B Biol. 2017, 175, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Marionnet, C.; Pierrard, C.; Golebiewski, C.; Bernerd, F. Diversity of biological effects induced by longwave UVA rays (UVA1) in reconstructed skin. PLoS ONE 2014, 9, e105263. [Google Scholar] [CrossRef]
- Zhong, J.; Li, L. Skin-derived precursors against UVB-induced apoptosis via Bcl-2 and Nrf2 up-regulation. BioMed Res. Int. 2016, 2016, 6894743. [Google Scholar] [CrossRef]
- Fu, H.; You, S.; Zhao, D.; An, Q.; Zhang, J.; Wang, C.; Wang, D.; Li, M. Tremella fuciformis polysaccharides inhibit UVA-induced photodamage of human dermal fibroblast cells by activating up-regulating Nrf2/Keap1 pathways. J. Cosmet. Dermatol. 2021, 20, 4052–4059. [Google Scholar] [CrossRef]
- Terra, V.A.; Souza-Neto, F.P.; Pereira, R.C.; Silva, T.N.X.; Costa, A.C.C.; Luiz, R.C.; Cecchini, R.; Cecchini, A.L. Time-dependent reactive species formation and oxidative stress damage in the skin after UVB irradiation. J. Photochem. Photobiol. B Biol. 2012, 109, 34–41. [Google Scholar] [CrossRef]
- Alafiatayo, A.A.; Lai, K.-S.; Ahmad, S.; Mahmood, M.; Shaharuddin, N.A. RNA-Seq analysis revealed genes associated with UV-induced cell necrosis through MAPK/TNF-α pathways in human dermal fibroblast cells as an inducer of premature photoaging. Genomics 2020, 112, 484–493. [Google Scholar] [CrossRef] [PubMed]
- Berneburg, M.; Gattermann, N.; Stege, H.; Grewe, M.; Vogelsang, K.; Ruzicka, T.; Krutmann, J. Chronically ultraviolet-exposed human skin shows a higher mutation frequency of mitochondrial DNA as compared to unexposed skin and the hematopoietic system. Photochem. Photobiol. 1997, 66, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Premi, S.; Brash, D.E. Chemical excitation of electrons: A dark path to melanoma. DNA Repair 2016, 44, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G.; Mahomoodally, M.F.; Aktumsek, A.; Jeko, J.; Cziaky, Z.; Rodrigues, M.J.; Custodio, L.; Polat, R.; Cakilcioglu, U.; Ayna, A.; et al. Chemical Profiling and Biological Evaluation of Nepeta Baytopii Extracts and Essential Oil: An Endemic Plant from Turkey. Plant 2021, 10, 1176. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Zhang, Y.; Fu, H.; Yao, F.; Liu, P.; Mo, Q.; Wang, D.; Zhao, D.; Wang, C.; Li, M. Physicochemical and Anti-UVB-Induced Skin Inflammatory Properties of Lacticaseibacillus Paracasei Subsp. Paracasei SS-01 Strain Exopolysaccharide. Fermentation 2022, 8, 198. [Google Scholar] [CrossRef]
- Fu, H.; Zhang, Y.; An, Q.; Wang, D.; You, S.; Zhao, D.; Zhang, J.; Wang, C.; Li, M. Anti-Photoaging Effect of Rhodiola Rosea Fermented by Lactobacillus Plantarum on UVA-Damaged Fibroblasts. Nutrients 2022, 14, 2324. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Luo, F.-J.; Ma, Y.-C.; Wang, S.-X.; Huang, J.; Qin, D.-D.; Xue, F.-F.; Liu, B.-Y.; Wu, Q.; Wang, X.-L.; et al. Dual Antioxidant Activity and the Related Mechanisms of a Novel Pentapeptide GLP4 from the Fermented Mycelia of Ganoderma Lingzhi. Food Funct. 2022, 1–17. [Google Scholar] [CrossRef]
- Qiu, X.; Cheng, J.-C.; Chang, H.-M.; Leung, P.C.K. COX2 and PGE2 mediate EGF-induced E-cadherin-independent human ovarian cancer cell invasion. Endocr. Relat. Cancer 2014, 21, 533–543. [Google Scholar] [CrossRef]
- Ahn, I.E.; Ju, J.H.; Lee, S.Y.; Park, J.S.; Oh, H.J.; Kim, H.R.; Lee, S.H.; Park, S.H.; Kim, H.Y.; Cho, M.L. Upregulation of stromal cell-derived factor by IL-17 and IL-18 via a phosphatidylinositol 3-kinase-dependent pathway. Scand. J. Immunol. 2012, 76, 433–439. [Google Scholar] [CrossRef]
- Domin, J. Phosphoinositide 3-kinase signalling pathways. J. Cell Sci. 2001, 114, 1439–1445. [Google Scholar]
- Franck, V.; Wagner, E.F.; Alain, M. Distinct involvement of the Jun-N-terminal kinase and NF-κB pathways in the repression of the human COL1A2 gene by TNF-alpha. Embo Rep. 2002, 3, 1069–1074. [Google Scholar]
- Xu, T.; Huang, M.; Xia, R.; Liu, X.; Sun, M.; Yin, L.; Chen, W.; Han, L.; Zhang, E.; Kong, R.; et al. Decreased expression of the long non-coding RNA FENDRR is associated with poor prognosis in gastric cancer and FENdRR regulates gastric cancer cell metastasis by affecting fibronectin1 expression. J. Hematol. Oncol. 2014, 7, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Chen, L.; Zhuang, Y.; Han, Y.; Tang, W.; Xia, F. Fibronectin 1 inhibits the apoptosis of human trophoblasts by activating the PI3K/Akt signaling pathway. Int. J. Mol. Med. 2020, 46, 1908–1922. [Google Scholar] [CrossRef]
- Wang, F.X. Ecdoin, a natural skin care factor from the Egyptian desert salt lake. China Cosmet. 2020, 5, 88–93. [Google Scholar]
- Buenger, J.; Dsiller, H. Ectoin: An effective natural substance to prevent UVA-induced premature photoaging. Ski. Pharmacol. Physiol. 2004, 17, 232–237. [Google Scholar] [CrossRef]
- Schroeter, M.-A.; Meyer, S.; Hahn, M.B.; Solomun, T.; Sturm, H.; Kunte, H.J. Ectoine protects DNA from damage by ionizing radiation. Sci. Rep. 2017, 7, 15272. [Google Scholar] [CrossRef]
- Botta, C.; Di Giorgio, C.; Sabatier, A.-S.; De Méo, M. Genotoxicity of visible light (400–800 nm) and photoprotEctoin assessment of ectoin, L-ergo-thioneine and mannitol and four sunscreens. J. Photochem. Photobiol. B Biol. 2008, 91, 24–34. [Google Scholar] [CrossRef]
- Heinrich, U.; Garbe, B.; Tronnier, H. In vivo Assessment of Ectoin:A Randomized, Vehicle-Controlled Clinical Trial. Ski. Pharmacol. Physiol. 2007, 20, 211–218. [Google Scholar] [CrossRef]
- Marini, A.; Reinelt, K.; Krutmann, J.; Bilstein, A. Ectoine-containing cream in the treatment of mild to moderate atopic dermatitis: A randomised, comparator-controlled, intra-individual double-blind, multi-center trial. Ski. Pharmacol. Physiol. 2014, 27, 57–65. [Google Scholar] [CrossRef]
- Addor, F.A.S.A. Topical effects of sCa® (Cryptomphalus aspersasecretion) associated with regenerative and antioxidant ingredients on aged skin: Evaluation by confocal and clinical microscopy. Clin. Cosmet. Investig. Dermatol. 2019, 12, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.-R.; Han, A.-R.; Park, S.-G.; Cho, C.-W.; Rhee, Y.-K.; Hong, H.-D. Effect of enzyme-assisted extraction on the physicochemical properties and bioactive potential of lotus leaf polysaccharides. Int. J. Biol. Macromol. 2020, 153, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Hseu, Y.-C.; Lo, H.-W.; Korivi, M.; Tsai, Y.-C.; Tang, M.-J.; Yang, H.-L. Dermato-protective properties of ergothioneine through induction of Nrf2/ARE-mediated antioxidant genes in UVA-irradiated Human keratinocytes. Free Radic. Biol. Med. 2015, 86, 102–117. [Google Scholar] [CrossRef]
- Li, Q.; Bai, D.; Qin, L.; Shao, M.; Zhang, S.; Yan, C.; Yu, G.; Hao, J. Protective effect of D-tetramannuronic acid tetrasodium salt on UVA-induced photo-aging in HaCaT cells. Biomed. Pharmacother. 2020, 126, 110094. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.-X.; Zhang, Z.-P.; Zhao, J.; Liu, J.-P. Effects of Fibronectin 1 on Cell Proliferation, Senescence and Apoptosis of Human Glioma Cells Through the PI3K/AKT Signaling Pathway. Cell. Physiol. Biochem. 2018, 48, 1382–1396. [Google Scholar] [CrossRef] [PubMed]
- Xian, D.; Xiong, X.; Xu, J.; Xian, L.; Lei, Q.; Song, J.; Zhong, J. Nrf2 Overexpression for the Protective Effect of Skin-Derived Precursors against UV-Induced Damage: Evidence from a Three-Dimensional Skin Model. Oxidative Med. Cell. Longev. 2019, 2019, 7021428. [Google Scholar] [CrossRef]
- Sosa, V.; Moline, T.; Somoza, R.; Paciucci, R.; Kondoh, H.; LLeonart, M.E. Oxidative stress and cancer: An overview. Ageing Res. Rev. 2013, 12, 376–390. [Google Scholar] [CrossRef]
- Sohal, R.S.; Allen, R.G. Oxidative stress as a causal factor in differentiation and aging: A unifying hypothesis. Exp. Gerontol. 1990, 25, 499–522. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Morry, J.; Ngamcherdtrakul, W.; Yantasee, W. Oxidative stress in cancer and fibrosis: Opportunity for therapeutic intervention with antioxidant compounds, enzymes, and nanoparticles. Redox Biol. 2017, 11, 240–253. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.; Kang, S.; Varani, J.; Bata-Csorgo, Z.; Wan, Y.; Datta, S.; Voorhees, J. Mechanisms of photoaging and chronological skin aging. Arch. Dermatol. 2002, 138, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.A.; Wang, Z.H.; Zhang, R.; Piao, M.J.; Kim, K.C.; Kang, S.S.; Kim, Y.W.; Lee, J.; Park, D.; Hyun, J.W. Myricetin Protects Cells against Oxidative Stress-Induced Apoptosisvia Regulation of PI3K/AKT and MAPK Signaling Pathway. Int. J. Mol. Sci. 2010, 11, 4348–4360. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.S.; Ko, Y.; Sklar, G.E. Identification and evaluation of pharmacists’ commonly used drug information sources. Ann. Pharmacother. 2009, 43, 347–352. [Google Scholar] [CrossRef]
- Han, L.; Yang, Y.; Yue, X.; Huang, K.; Liu, X.; Pu, P.; Jiang, H.; Yan, W.; Jiang, T.; Kang, C. Inactivation of PI3K/AKT signaling inhibits glioma cell growth through modulation of β-catenin-mediated transcription. Brain Res. 2010, 1366, 9–17. [Google Scholar] [CrossRef]
- Datta, S.R.; Brunet, A.; Greenberg, M.E. Cellular survival: A play in three AKTs. Genes Dev. 1999, 13, 2905–2927. [Google Scholar] [CrossRef]
- Xie, Y.; Shi, X.; Sheng, K.; Han, G.; Li, W.; Zhao, Q.; Jiang, B.; Feng, J.; Li, J.; Gu, Y. PI3K/AKT signaling transduction pathway, erythropoiesis and glycolysis in hypoxia (Review). Mol. Med. Rep. 2019, 19, 783–791. [Google Scholar] [CrossRef]
- Yang, S.S.; Zhou, L.; Nephrology, D.O. Resveratrol Inhibits Oxidative Stress-mediated Apoptosis in Renal Tubular Epithelial Cells by Activating PI3K/AKT Pathway. Mod. Chin. Med. 2019, 21, 913–919. [Google Scholar]
- Zhang, X.; Xue, H.; Zhou, P.; Liu, L.; Yu, J.; Dai, P.; Qu, M. Angelica polysaccharide alleviates oxidative response damage in HaCaT cells through up-regulation of miR-126. Exp. Mol. Pathol. 2019, 110, 104281. [Google Scholar] [CrossRef]
- Wu, J.; Ibtisham, F.; Niu, Y.F.; Wang, Z.; Li, G.H.; Zhao, Y.; Nawab, A.; Xiao, M.; An, L. Curcumin inhibits heat-induced oxidative stress by activating the MAPK-Nrf2/ARE signaling pathway in chicken fibroblasts cells. J. Therm. Biol. 2019, 79, 112–119. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Direction | Primer Sequences (5′-3′) |
|---|---|---|
| GAPDH | F | TCAGACACCATGGGGAAGGT |
| R | TCCCGTTCTCAGCCATGTAG | |
| COL4A5 | F | CAAGGTCTACCAGGTCCAGAA |
| R | TCATTCCATTGAGACCCGGC | |
| FN1 | F | CCCAATTGAGTGCTTCATGCC |
| R | CCTCCAGAGCAAAGGGCTTA | |
| IGF2 | F | TCCTGTGAAAGAGACTTCCAG |
| R | GTCTCACTGGGGCGGTAAG | |
| COL1A1 | F | GAGGGCCAAGACGAAGACATC |
| R | CAGATCACGTCATCGCACAAC | |
| COL1A2 | F | GTTGCTGCTTGCAGTAACCTT |
| R | AGGGCCAAGTCCAACTCCTT | |
| PIK3R1 | F | ACCACTACCGGAATGAATCTCT |
| R | GGGATGTGCGGGTATATTCTTC | |
| NR4A5 | F | ATGCCCTGTATCCAAGCCC |
| R | GTGTAGCCGTCCATGAAGGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, W.; An, Q.; Zhang, J.; Shi, X.; Wang, C.; Li, M.; Zhao, D. Protective Effect of Ectoin on UVA/H2O2-Induced Oxidative Damage in Human Skin Fibroblast Cells. Appl. Sci. 2022, 12, 8531. https://doi.org/10.3390/app12178531
Cheng W, An Q, Zhang J, Shi X, Wang C, Li M, Zhao D. Protective Effect of Ectoin on UVA/H2O2-Induced Oxidative Damage in Human Skin Fibroblast Cells. Applied Sciences. 2022; 12(17):8531. https://doi.org/10.3390/app12178531
Chicago/Turabian StyleCheng, Wenjing, Quan An, Jiachan Zhang, Xiuqin Shi, Changtao Wang, Meng Li, and Dan Zhao. 2022. "Protective Effect of Ectoin on UVA/H2O2-Induced Oxidative Damage in Human Skin Fibroblast Cells" Applied Sciences 12, no. 17: 8531. https://doi.org/10.3390/app12178531
APA StyleCheng, W., An, Q., Zhang, J., Shi, X., Wang, C., Li, M., & Zhao, D. (2022). Protective Effect of Ectoin on UVA/H2O2-Induced Oxidative Damage in Human Skin Fibroblast Cells. Applied Sciences, 12(17), 8531. https://doi.org/10.3390/app12178531