Abstract
Peptidomics refers to the comprehensive profiling of endogenous peptides obtained from biological sources. The formation of endogenous peptides is dependent on not only endogenous factors but also exogenous factors such as microbial proteases or process conditions, including fermentation. This study analyzed the probiotic strains of Lactobacillus rhamnosus LOCK900 (LOCK), Bifidobacterium animalis ssp. lactis BB-12 (BB12), and potential probiotic Lactobacillus acidophilus Bauer Ł0938 (BAUER) to assess their ability of fermentation and peptide production in dry-cured pork loin. The peptides obtained after in vitro digestion were characterized by liquid chromatography–tandem mass spectrometry. Based on the sequences identified, the degree of similarity or differences between the peptides was determined and presented graphically on the factor plane. The charts showed that the meat products aged for 180 and 270 days were the most diverse when BB12 or BAUER were used as starter cultures. Myosin and keratin were identified as the most likely precursors of bioactive peptides in products obtained using this strain of lactic acid bacteria (LAB). The knowledge acquired from this study may contribute to the design of functional meat products as the results revealed not only the peptidogenic potential of the LAB strains indicated on their beneficial effect on the bioactivity of peptides.
1. Introduction
Dry-cured meat products are among the most popular meat products. The production of these products requires a considerable period of time during which their characteristics are shaped. In this long duration, several physicochemical changes occur in the meat tissue, which contribute to the high quality of meat products. During aging, intensive proteolysis takes place in meat, resulting in the formation of a large numberof peptides and free amino acids responsible for the final taste and aroma of cured meats. Moreover, due to their unique biological activity, protein-derived compounds or peptides released during aging exhibit health-promoting effects. It has been shown that raw pork or pork products are a good source of bioactive peptides thatare produced after digestion under controlled hydrolysis conditions by various proteases [1,2], including those typical of the human gastrointestinal tract [3,4,5]. The literature reports indicate that peptides present in dry-cured meat products are characterized by antioxidant, angiotensin-converting enzyme(ACE)-inhibitory, and dipeptidyl peptidase IV (DPP-IV)-inhibitory effects. Peptide production from meat is associated with the activity of endogenous muscle enzymes found in meat products [6,7,8], as well as intense protein degradation [9,10]. The raw material—meat—itself has a great influence on the performance of peptides, which can be related to the differences in the type of muscles of the carcass and the methods used for rearing. Recently, Mora et al. [11] indicated that peptide extracts obtained from Teruel (Spanish product), Parma ham (Italian product), and dry-cured ham from Belgium had varied peptide sequences depending on the type of ham [11]. The authors attributed these variations to the different processing methods used, and above all, different aging times. Similarly, Wang et al. [12] evaluated the proteome of Xuanwei and Jinhua ham after 2 years of aging and found differences in peptides (especially in those with a molecular weight of <2500 Da) in these products. The authors indicated that the peptide profile could be influenced by the technological steps applied during production, which is characteristic of a given region. It has also been reported that the dry-cured time of meat has an impact on the peptides obtained after simulated in vitro gastrointestinal digestion [13,14].
Dry-cured products, based on both whole muscles and minced form (e.g., sausages), are produced using lactic acid bacteria (LAB) that can carry out fermentation processes. Due to their ability to produce lactic acid, these bacteria rapidly decrease the pH of the product, and, thus, increase its safety by inactivating pathogens and inhibiting undesirable changes caused by harmful microorganisms. Moreover, acidification of the environment creates appropriate biochemical conditions that shape the sensory properties of products during aging [15]. In addition to their role in improving the physicochemical, rheological, and organoleptic properties of the products, LAB strains may also contribute to the release of bioactive products through proteolysis during their fermentation and aging, which can be attributed to their proteolytic properties. Thus, the use of LAB strains with proteolytic potential has been recognized as a new strategy for the production of functional foods rich in bioactive peptides [16,17,18,19]. For instance, the effect of starter culture (Pediococcus acidilactici MS200 and Staphylococcus vitulus RS34) and protease addition (Erg222) on the bioactive effects of peptides obtained from dry fermented sausages was recently investigated [20]. The results showed that the ACE-inhibitory and antioxidant activity of the protease batches were increased after 63 days of aging, indicating their excellent stability after simulated in vitro gastrointestinal digestion. Among the LAB strains, some have been assigned the “probiotic” status. Probiotics are defined as nonpathogenic, living microorganisms that are resistant to the action of digestive enzymes, and when consumed in adequate amounts (106–108 cfu/g or 108–1010 cfu/day), these organisms have a positive effect on the physiology of the gastrointestinal tract [21]. Probiotic bacterial strains promote microbial balance in the intestines by increasing the effects of beneficial bacteria and/or neutralizing the effects of harmful microorganisms. Thus, these strains are used, for example, to prevent diarrhea and constipation and treat food allergies. Regardless of the biological activity associated with the probiotic properties of bacterial strains, their proteolytic properties can be utilized to release peptides, including the biologically active ones.
In this work, a peptidomic approach was used to identify and quantify the peptides generated during the dry-curing of pork loin and to find sequences exhibiting potential bioactivities. According to the adopted hypothesis, the hydrolytic release of peptides is regulated not only by the prolonged aging period but also by the presence of LAB, such as Lactobacillus rhamnosus LOCK900 (LOCK), Lactobacillus acidophilus Bauer Ł0938 (BAUER), and Bifidobacterium animalis ssp. lactis BB-12. These strains have been well described in the literature, and their use has not been linked with any negative effects in meat products [22,23,24,25,26,27]. It was assumed that the amount of potentially bioactive peptides released during the gastrointestinal digestion of meat is strongly determined by the degree of protein degradation occurring during the aging of meat products. Therefore, the study attempted to assess the trends among the identified peptides after simulated digestion and absorption in the gastrointestinal tract. For this purpose, dry-cured loin extracts were analyzed at different processing times (28, 90, 180, 270, and 360 days) after pepsin and pancreatin hydrolysis by liquid chromatography–tandem mass spectrometry (LC–MS/MS). Based on the differences identified from the relative quantification of the peptides, differences between the variants digested with different LAB strains were determined at different processing times.
2. Materials and Methods
2.1. Preparation of Dry-Cured Meat Products
Dry-cured meat cuts from Polish White Large breed were processed following the traditional procedure for different times (28, 90, 180, 270, and 360 days). Meat was excised at 24 h postmortem from half carcasses chilled at 4 °C coming from pigs reared under the same conditions. At 48 h postmortem, all loins (m. longissimus thoracis) (n = 8) were cured with a curing mixture (20 g of NaCl, 9.7 g of curing salt, and 0.3 g of NaNO3/kg loin) by surface massage. All the cured batches were maintained at 4 °C for 24 h to allow the diffusion of curing salt. After curing, loins were portioned in parts of about 1000 g, which were randomly divided into four experimental groups, with three samples per group.
One of the groups (C) was not treated further, while the three other groups were each inoculated with 0.2% (v/w) L. rhamnosus LOCK900 (LOCK; strain deposit number: CP00548), L. acidophilus Bauer Ł0938 (BAUER), and B. animalis ssp. lactis BB-12 (BB12; strain deposit number: DSM15954) to achieve 106–107 cfu/g meat. Bacteria strains were transferred onto the meat surface. Subsequently, the meat portions were hung in a laboratory aging chamber at 16 ± 1 °C under a relative humidity of 75 ± 5% for 21 days. Then, the whole loin pieces were vacuum-packed and aged at 4 ± 1 °C for 360 days. Two independent experimental trials were conducted on the prepared loin samples.
2.2. Extraction and Digestion Meat Protein
Extraction and gastrointestinal digestion of muscle protein were performed as described earlier [27]. The water-soluble fraction (WSF) of meat proteins was extracted by homogenizing for 5 min (T25 Basic ULTRA-TURRAX; IKA, Staufen, Germany) 10 g of meat and distilled water (1:10 w/v) on ice. To prepare the salt-soluble fraction (SSF), the pellet resulting from the WSF extraction was re-suspended in 0.6 M NaCl in 0.1 M phosphate buffer (pH 6.2) in a ratio of 1: 6 and homogenized for 1 min on ice. The resulting homogenate was deaerated prior to extraction for 18 h at 4 °C. After the centrifugation step at 10,000× g, 4 °C for 10 min, the supernatant was filtered through Whatman Filter Paper No. 1. WSF and SSF were subjected to in vitro hydrolysis using pepsin and pancreatin. As a gastric hydrolysis phase, the protein fractions were adjusted to a pH of 2.0 with 1 M HCl. A 6 M HCl (pH 2.0) pepsin solution was added to protein fractions at a ratio of 1:100, enzyme to substrate. The hydrolysis process was performed for 2 h at 37 °C in darkness and with continuous stirring, after which the solution was neutralized to pH 7.0 with 1 M NaOH to enable enzyme inactivation. Next, pancreatin was added at a ratio of 1:50, enzyme to substrate, for simulated intestinal hydrolysis (intestinal hydrolysis steps; at 37 °C for 3 h in darkness with continuous stirring). Incubation was carried out for 3 h at 37 °C in the dark with continuous stirring. The enzyme was inactivated through heating at 95 °C for 10 min. Then, hydrolysates were dialyzed by using membrane tubes (7 kDa molecular weight cut-off, Spectra/Por®) against phosphate-buffered saline (PBS; pH 7.4; 1:4, v/v) for 1 h at 37 °C without light as a simulated absorption step. Hydrolysates obtained after simulated gastrointestinal hydrolysis and absorption were concentrated in the evaporator and dissolved in 2 mL of 0.01 M HCl prior to chromatographic analysis.
2.3. Peptide Identification
Peptides were analyzed by LC–MS/MS. Before the analysis, the samples were concentrated and desalted on an RP-C18 precolumn (Waters Corp., Milford, MA, USA). Separation was performed on an RP-C18 nano-Ultra Performance column (Waters, BEH130 C18 column, 75 μm i.d., 250 mm long) of a nanoACQUITY UPLC system (Warsaw, Poland) using a 180 min linear acetonitrile gradient (0–35%) at a flow rate of 250 nL·min−1. The column outlet was directly connected to a mass spectrometer (Orbitrap Velos, Thermo Fisher Scientific Inc., Waltham, MA, USA) for the analysis [27]. The procedure was performed twice for each research variant, and the sequences present in both replications were qualified for the next stage of the research.
2.4. Data Analysis
The raw data files were preprocessed using Mascot Distiller software (version 2.4.2.0, Matrix Science Inc., Boston, MA, USA). The obtained peptide masses and their identified fragmentation pattern were compared with the protein sequence database (UniProt KB) [28] using the Mascot search engine (Mascot Daemon v. 2.4.0, Mascot Server v.2.4.1, Matrix Science, London, UK). The “mammals” option was chosen as the taxonomy constraint parameter. The search parameters applied were as follows: enzyme specificity, none; peptide mass tolerance, 5 ppm; and fragment mass tolerance, 0.01 Da. The protein mass was left unrestricted, and the mass values were assumed as monoisotopic with a maximum of two missed cleavages allowed. Methylthiolation, oxidation, and carbamidomethylation were set as fixed and variable modifications. The peptide sequences from unknown original proteins were excluded. Peptide identification was performed using the Mascot search engine (Matrix Science), with a probability-based algorithm. The expected value threshold was set at 0.05 for the analysis (all peptide identification had <0.05% chance of being a random match). The principal component analysis (PCA) method was used to graphically present the relationship between the peptide sequences. PCA was performed using STATISTICA®13.1. software. The number of ingredients was set to the maximum and the minimum custom was set to 1.0. The standardization of variables was performed by default in the statistical program. The correct use of PCA was confirmed by the Kaiser–Meyer–Olkin test for sampling adequacy and the Bartlett sphericity test. The number of main components was determined as 4 based on the percentages of the explained variance (Kaiser criterion <80% was adopted). Normal distribution analysis was not performed when performing PCA.
3. Results and Discussion
This study used a peptidomic approach to evaluate peptides resulting from the digestion of a fermented meat product with selected probiotic (or potential probiotic) LAB strains during long-term aging. This approach has already been presented by various authors, including Paolella et al. [13] who indicated that dry-cured hams can be grouped according to their aging times (18–24 months) after the gastrointestinal digestion stage in the semiquantitative peptide evaluation. On the other hand, a peptidomic approach was used to analyze meat protein degradation in a low-sodium fermented sausage model using autochthonous starter cultures. It was found that the LAB strain present in the model systems generated a unique profile of small peptides and amino acids that served as a distinctive biochemical parameter to distinguish between fermented meat products [29].
Cleavage of peptide bonds by digestive proteases has been shown to result in the release of peptides of various lengths and free amino acids. In this study, peptides were identified based on the results obtained in the LC–MS/MS Orbitrap analyses. As mentioned earlier, most of the peptides found in the profile of the digested samples had a molecular weight between 200 and 1700 Da, which is typical of most known bioactive peptides, as well as peptides that can easily pass through the intestinal lumen and therefore potentially influence nutritional or systemic bioactive function. The length of the peptide fragments was 2–47 amino acids, which is in line with other studies of the peptide profile obtained by digestion [5,30].
3.1. Peptide Profile of Digesta Depending on LAB Strains and Aging Time
Meat proteolysis is a very dynamic process, in which processing time and the presence of microorganisms contribute to variability. This is especially true in the case of dry-cured meat products, where the meat proteome may have a different profile and thus exhibit different biological activities. To understand the effect of time (28, 90, 180, 270, and 360 days) and selected LAB strains on the peptide profile, a PCA scoring plot was constructed to explain the variations associated with processing times, identify those peptides that mainly differentiate the samples, and determine the degree of similarities and differences between the peptides.
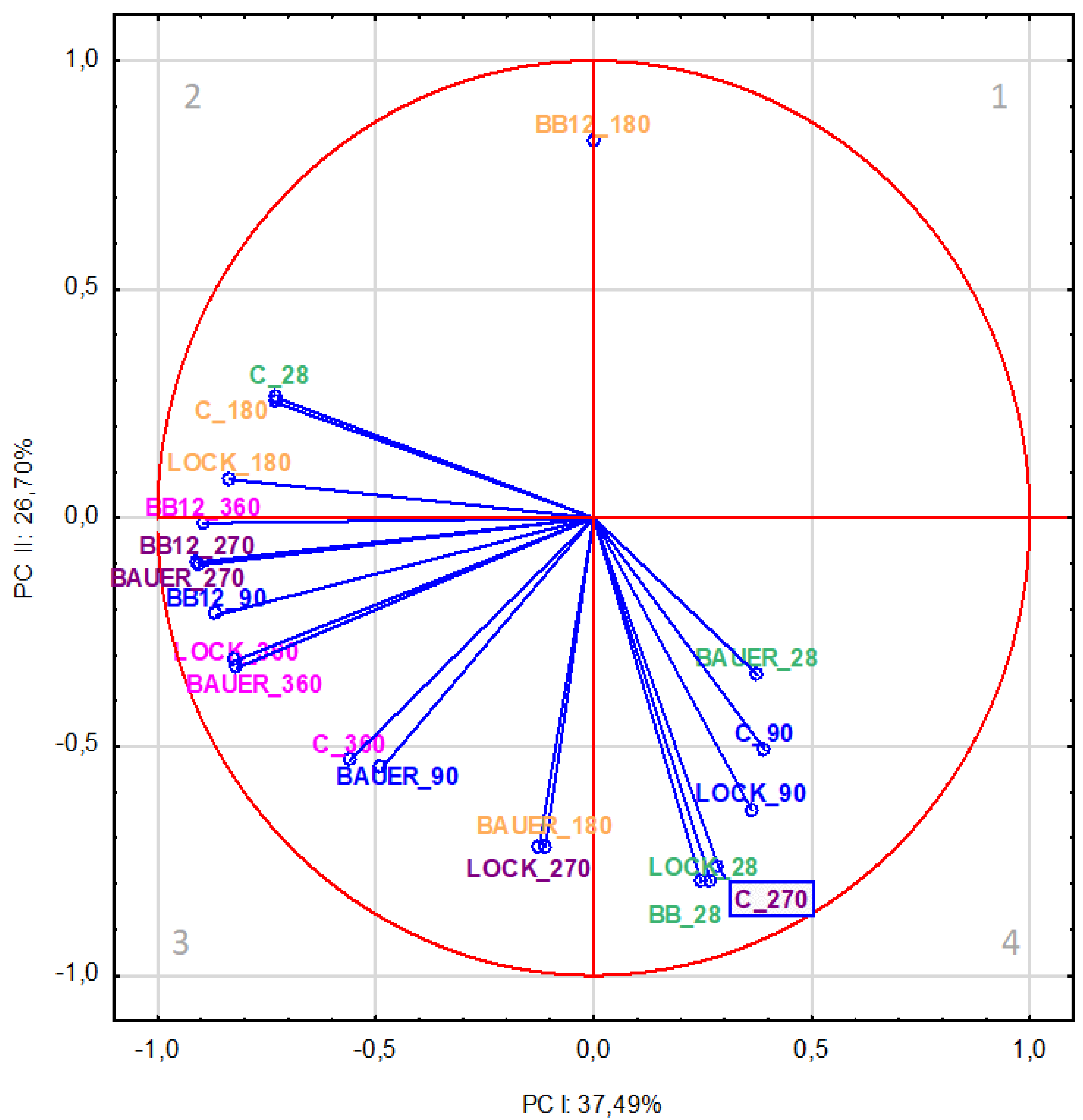
To ensure partial visualization of the data related to the effect of the type of LAB strain used on the formation of peptides in the product at different aging times, the variables were projected on the factor plane (PC-I × PC-II), as shown in Figure 1 and Figure 2. Changes in factors calculated in the factor analysis are presented in Table 1 and Table 2. For the water-soluble fraction (WSF), the results showed a relationship between the research variant (type of LAB strain used) and the aging time of pork loin. The analysis of factor loadings (Table 1) revealed that the first component (PC-I) included nine most strongly correlated variables, accounting for 37.49% of the total variance. It is worth noting that the most diverse were samples aged for 270 days (BB12, BAUER) and 360 days (LOCK, BB12, BAUER), while the C samples were also variable after 28 days and BB12 and BAUER samples after day 90 of the production (Table 1). Indeed, Okoń et al. [31] reported that the dry-cured pork loins with the probiotic LOCK strain had fewer free amino acids and peptides (WSF) compared to the control sample after 2 months of refrigerated storage. During the same period, dry-cured loin samples inoculated with BAUER contained more peptides, while the sample with Bifidobacterium animalis ssp. lactis BB-12 contained the smallest number of peptides, compared to the sample lacking starter cultures [32]. Okoń et al. [33] also proved that BB12 and BAUER caused changes in the quantitative and qualitative profile of amino acids in dry-cured loins after 9 months, thus confirming their participation in the proteolysis of meat protein components. Comparing this finding with previous reports on the effect of probiotic LAB strains in the production of dry-cured loins [17], it can be stated that treatment and storage time can significantly influence the proteolysis index of dry-cured loin (p < 0.01). In that study [17], this index increased gradually during storage, reaching the maximum value in C and LOCK samples after 270 days of storage, and in BAUER and BB12 samples after 360 days.
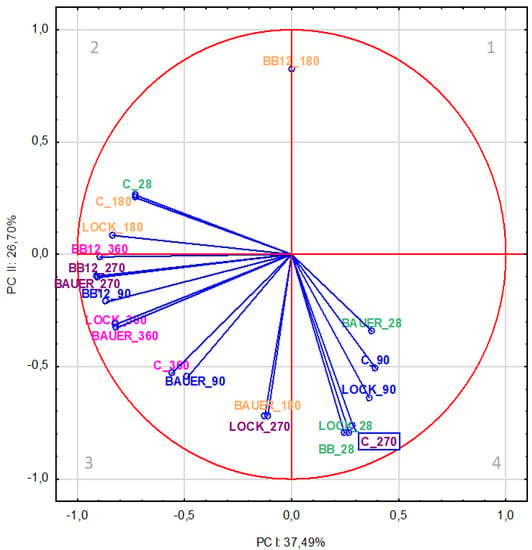
Figure 1.
Projection of variables (time, LAB strain) on the factor plane (PC-I × PC-II) for water-soluble fraction based on the peptides from dry-cured pork loin.
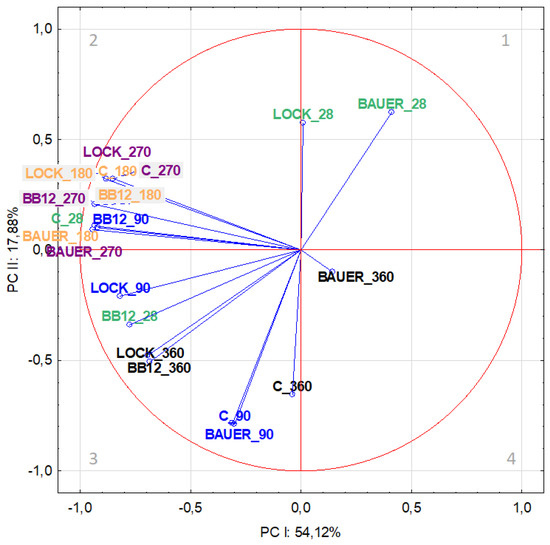
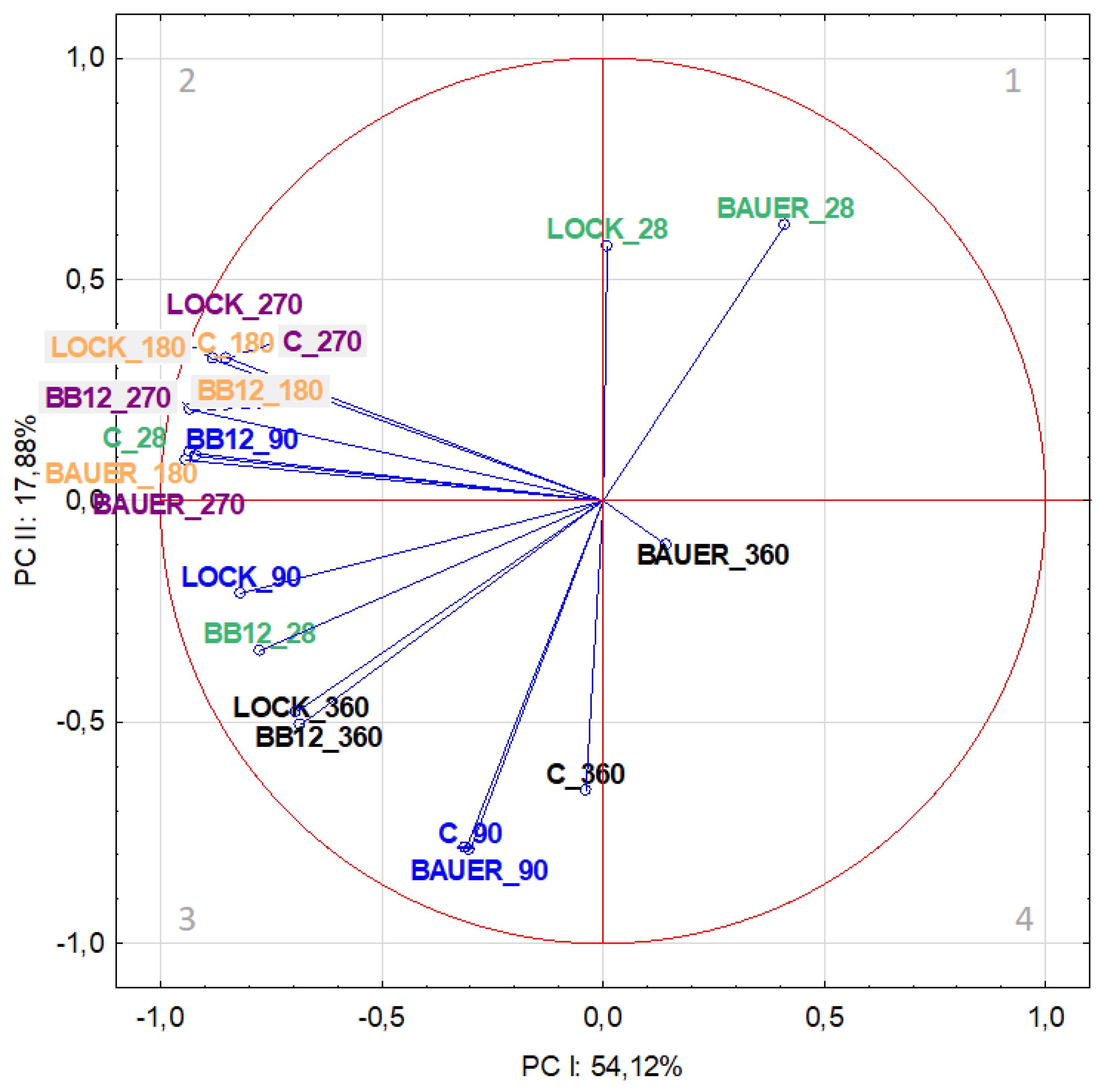
Figure 2.
Projection of variables (time, LAB strain) on the factor plane (PC-I × PC-II) for salt-soluble fraction based on the peptides from dry-cured pork loin.

Table 1.
Correlations between variables and factors in PCA based on the factor loadings for water-soluble fraction (WSF) and salt-soluble fraction (SSF).
On the other hand, as shown in Figure 1, after 28 days of storage, the effect of LAB strains was more pronounced, which was positively correlated with PC-I, although previous studies showed no significant difference (p > 0.05) in the content of primary amines in WSF between control and inoculated samples on day 28 of the in vitro test [17]. This discrepancy may be explained by the fact that in the present study, no additional proteolysis occurred in the inoculated dry-cured loin compared to the control during the initial storage stage in quantification, but qualitative changes were observed in peptides released from meat proteins under the influence of microbial peptidases. This confirms the variability of the proteome during the production of meat products, which was directly reflected by the results observed after the simulated gastrointestinal digestion process in this study. It is also worth noting that the variables related to the results of the trials in the initial production period, i.e., 28 and 90 days, were in quadrant 4, which implies that they were positively correlated with the first component. The second PC (PC-II) accounting for 26.7% of the variance was mainly determined by the action of LAB strains in WSF on day 28 (LOCK and BB12) and after 180 (BB12 and BAUER) and 270 (C and LOCK) days of aging (Table 1).
The distribution of variables on the factor plane for SSF is shown in Figure 2. The analysis of correlations between variables and factors in PCA based on factor loadings showed that in the first component, the most strongly correlated variables were associated with the middle period of the study, i.e., 180 and 270 days of aging, irrespective of the LAB strain (Table 1), and their location in quadrants 2 and 3 on Figure 2 indicates that they were negatively correlated with PC-I. PC-II accounting for 17.78% of the variance was mainly determined by the action of variants C and BAUER on day 90 of the analysis.
Considering the distribution of variables in individual areas of the graphical presentation of PCA, it was found that the LOCK batches showed the greatest similarity to the control sample, regardless of time. In terms of WSF, it can be observed, inter alia, in quadrants 2 (C and LOCK after 180 days of aging) and 4 (C and LOCK after 90 days of aging) (Figure 1). However, for SSF, it can be observed, among others, in quadrant 2 (C and LOCK after 180 and 270 days of aging), as well as quadrant 3 (C and LOCK after 90 and 360 days of aging), although, in this case, the distance between the variants was greater (Figure 2). The observed trend can be explained by the minor influence of the LOCK strain on changes in the protein fraction resulting from proteolysis, which is in line with the findings reported by other authors [17,34,35].
3.2. Potentially Bioactive Peptides
In previous studies, meat hydrolysates showed several biological activities that can be attributed to the presence of bioactive peptide sequences encrypted in meat proteins and released first during processing and then during digestion by gastrointestinal proteases [27,36,37]. The antioxidant and antihypertensive or antidiabetic activity of the peptides were analyzed based on the same LAB strains used in the study. Based on the results of those studies, their results were compared and only the test variants that showed the most favorable biological activity during the 360-day processing period are presented in Table 2.
Table 2.
Test variant showing the strongest biological effect—influence of probiotic LAB strains on the biological activity of peptides after digestion with pepsin and pancreatin(<7 kDa).
Literature data indicate that pork loin peptides showed biological activities, i.e., the ability to inhibit ACE and DPP-IV, and an antioxidant effect in in vitro tests (as summarized in Table 2). As the authors of other studies have indicated, among all variants in all analysis periods, the same as in this study, the highest DPP-IV inhibiting activity and the highest ACE-inhibiting activity at the level of 81.05% and 67.07% (WSF), respectively, were recorded for BB12 (although for BB12 and C, p > 0.05) after 180 days [27,37]. Similarly, after 180 days, the strongest anti-radical effect was recorded at the level of 88.96% in WSF for the BAUER batches [36]. Taking into account SSF, the strongest DPP-IV inhibitory protein (BB12 and LOCK at the level of 75.45% and 78.10%, p > 0.05) and the strongest anti-radical effect (BB12 and C at 89.58% and 90.48%, p > 0.05) were recorded after 180 days. In turn, the ability to inhibit ACE (BB12 with an inhibitory effect of 71.19%) was the strongest after 270 days. In terms of the strain type, BB12 and BAUER variants were the most promising, because the samples of meat products inoculated with BAUER were characterized by the strongest antiradical (against ABTS) effect [36], while the BB12 variant showed the strongest DPP-IV-inhibitory effect and ACE-inhibitory effect [27,37], reflecting its antidiabetic and antihypertensive potential. Okoń [33] also observed the highest antioxidant activity in peptides isolated from 9-month-old (270-day-old) loins with BB12, which showed the most effective ABTS cationic radical scavenging effect. As shown in Figure 1, BB12 and BAUER were on opposite positions on the chart during 180 and 270 days for WSF but not SSF. This demonstrates the large variations in water-soluble peptide sequences between these variants. Therefore, peptides present in the hydrolysates isolated from BB12 and BAUER variants (after 180 and 270 days) were taken into account for further analysis. The graphical presentation of the results suggested that during this time period, the sequences from SSF were more similar than those from WSF, so WSF was considered in the further stages of the study. First, the sequences shared with the control sample and then with LOCK were removed from the analysis, in order to determine the actual influence of these two strains on the generation of bioactive sequences. A total of 401 peptide sequences were obtained after 180 days, of which only 22 were common to both research variants. On the other hand, in the second research period (270 days), the sequence similarity was found to be much higher, with 140 fragments out of 460 identified peptide sequences common to BB12 and BAUER.
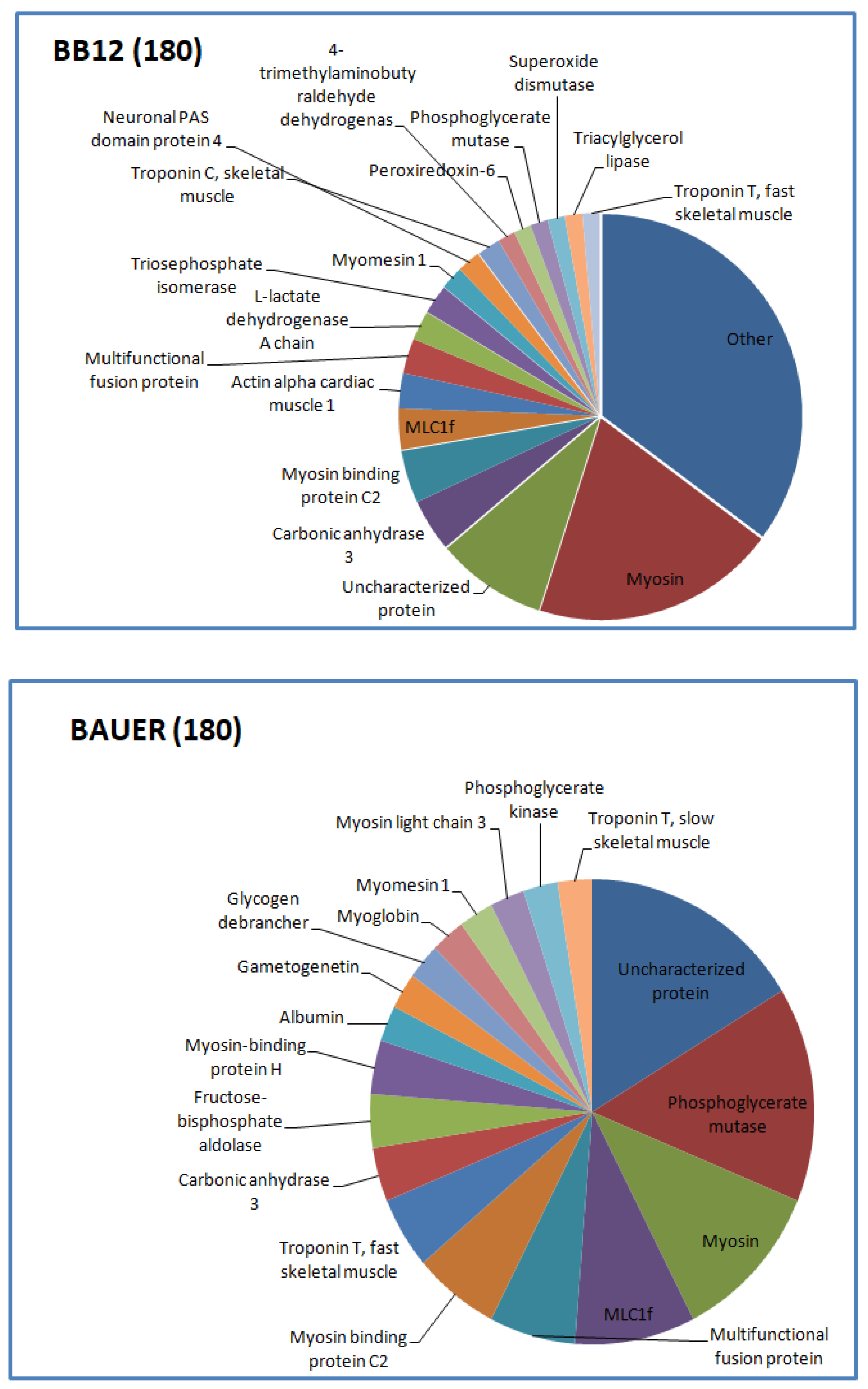
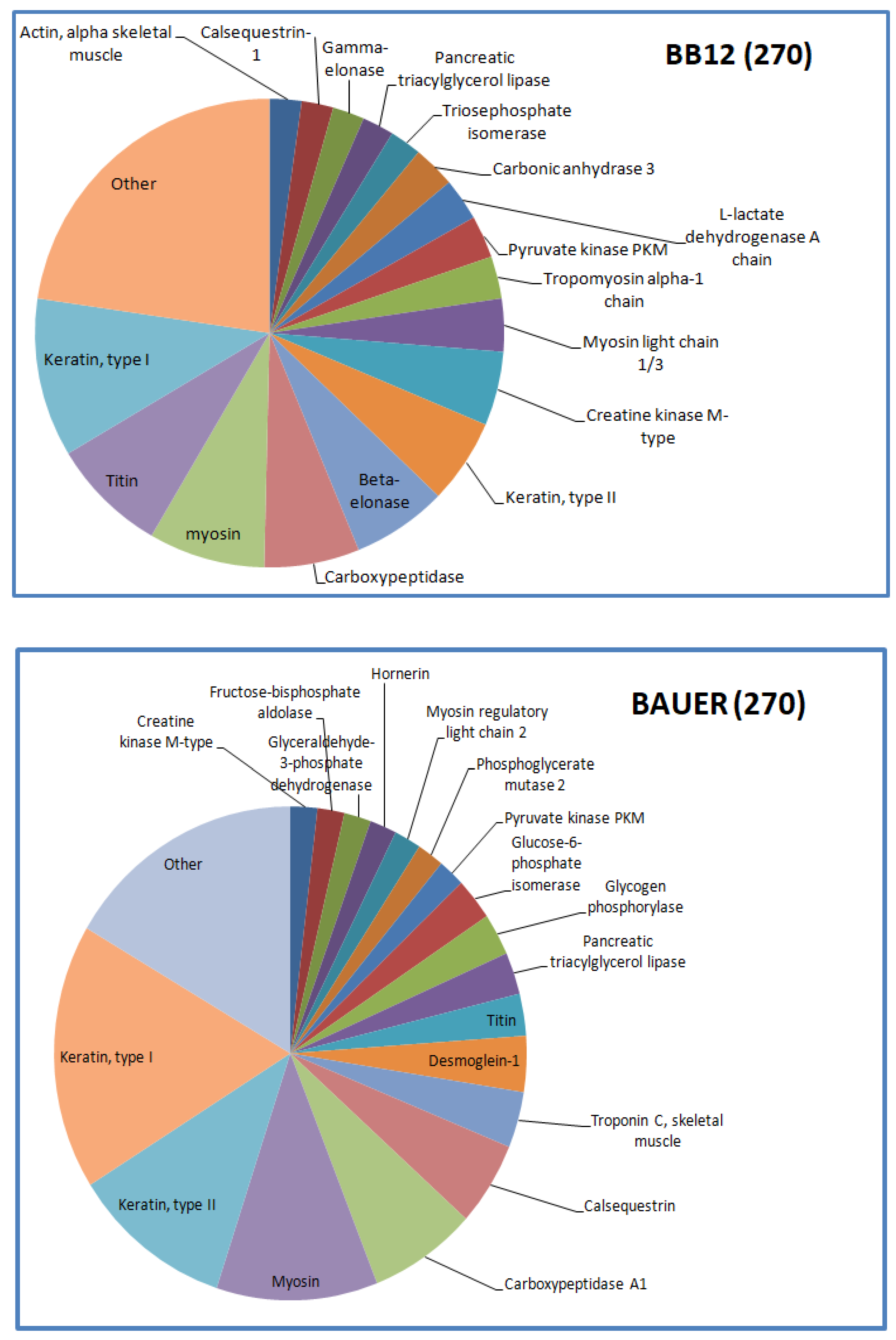
The distribution of the identified peptides in terms of their protein origin is shown in Figure 3 and Figure 4. A different number of protein fragments was identified in the tested variants, indicating no specific influence of the type of strain (BB12 vs. BAUER), but an influence of aging time on the peptide profile in the hydrolysates.
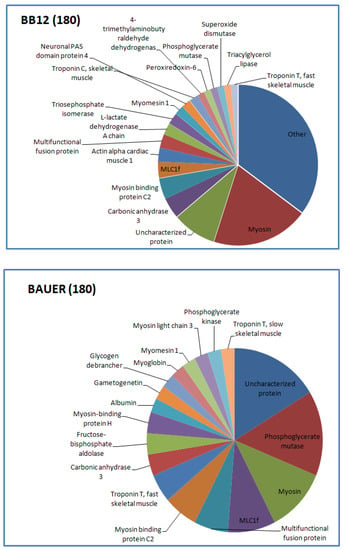
Figure 3.
Distribution of the identified peptides typical for BB12 and BAUER from water-soluble fraction after 180 days.
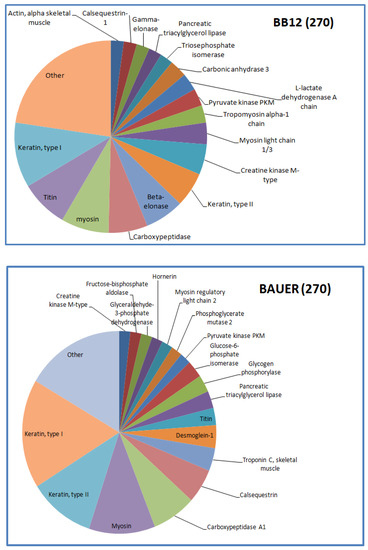
Figure 4.
Distribution of the identified peptides typical for BB12 and BAUER from water-soluble fraction after 270 days.
Myosin-derived peptides were predominant in both BB12 and BAUER variants after 180 days. In addition, at this time, phosphoglycerate mutase protein chains were degraded and a large number of different peptides was observed in the BAUER variant. Phosphoglycerate mutase is an enzyme involved in glycolysis, and its shorter fragments (peptides) can be found in the meat matrix. In a previous study, peptides from this enzyme remained stable during a long storage period in pork loins; however, these products were not inoculed with LAB [17]. The participation of the BAUER strain observed in this study may indicate their role in shaping the pool of various peptides, first in the meat matrix and then during hydrolysis. Keratin is a structural protein of animal origin, and present as cytoskeletal and microfibrillar keratin in two types: type I (acidic; 40–55 kDa) and type II (neutral to basic; 56–70 kDa) [28]. It is commonly found in meat processing wastes, such as chicken feathers or pig hair. Keratin peptides formed after the hydrolysis of these waste products exhibit valuable biological properties, including antioxidant, ACE-inhibitory, and DPP-IV-inhibitory activity [38,39,40,41]. In the present study, a significant share of keratin-derived peptides (BAUER—type I, BB12—types I and II) was observed in the hydrolysates after 270 days of aging. Recently, López-Pedrouso et al. [42] discovered a strong antioxidant peptide FLEQQNQVLQTK derived from keratin in liver porcine hydrolysates.
4. Conclusions
The results of the study emphasize that proteolysis occurring during aging has a strong impact on the peptide profile of the digested sample, and thus the nutritional value and potential biological activity of peptides encrypted in meat proteins. In particular, the amount of potentially bioactive peptides released during meat digestion can vary depending on the probiotic LAB strains used for production and the aging time of dry-cured products.
Author Contributions
Conceptualization, P.K. and J.S.; methodology, P.K.; formal analysis, J.S.; investigation, P.K.; data curation, P.K.; writing—original draft preparation, P.K.; writing—review and editing, J.S.; visualization, P.K. and J.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Arihara, K.; Yokoyama, I.; Ohata, M. Bioactivities generated from meat proteins by enzymatic hydrolysis and the Maillard reaction. Meat Sci. 2021, 180, 108561. [Google Scholar] [CrossRef] [PubMed]
- Choe, J.; Seol, K.H.; Son, D.I.; Lee, H.J.; Lee, M.; Jo, C. Identification of angiotensin I-converting enzyme inhibitory peptides from enzymatic hydrolysates of pork loin. Int. J. Food Prop. 2019, 22, 1112–1121. [Google Scholar] [CrossRef] [Green Version]
- Wen, S.; Zhou, G.; Song, S.; Xu, X.; Voglmeir, J.; Liu, L.; Li, C. Discrimination of in vitro and in vivo digestion products of meat proteins from pork, beef, chicken, and fish. Proteomics 2015, 15, 3688–3698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escudero, E.; Aristoy, M.C.; Nishimura, H.; Arihara, K.; Toldrá, F. Antihypertensive effect and antioxidant activity of peptide fractions extracted from Spanish dry-cured ham. Meat Sci. 2012, 91, 306–311. [Google Scholar] [CrossRef]
- Escudero, E.; Sentandreu, M.A.; Arihara, K.; Toldra, F. Angiotensin I-converting enzyme inhibitory peptides generated from in vitro gastrointestinal digestion of pork meat. J. Agric. Food Chem. 2010, 58, 2895–2901. [Google Scholar] [CrossRef]
- Kaur, L.; Hui, S.X.; Morton, J.D.; Kaur, R.; Chian, F.M.; Boland, M. Endogenous proteolytic systems and meat tenderness: Influence of post-mortem storage and processing. Food Sci. Anim. Resour. 2021, 41, 589. [Google Scholar] [CrossRef]
- Bhat, Z.F.; Morton, J.D.; Mason, S.L.; Bekhit, A.E.D.A. Role of calpain system in meat tenderness: A review. Food Sci. Hum. Wellness 2018, 7, 196–204. [Google Scholar] [CrossRef]
- Hu, S.; Zhou, G.; Xu, X.; Zhang, W.; Li, C. Contribution of cathepsin B and L to endogenous proteolysis in the course of modern Jinhua ham processing. Food Control 2022, 135, 108584. [Google Scholar] [CrossRef]
- Di Luccia, A.; Picariello, G.; Cacace, G.; Scaloni, A.; Faccia, M.; Liuzzi, V.; Musso, S.S. Proteomic analysis of water soluble and myofibrillar protein changes occurring in dry-cured hams. Meat Sci. 2005, 69, 479–491. [Google Scholar] [CrossRef]
- López, C.M.; Bru, E.; Vignolo, G.M.; Fadda, S.G. Identification of small peptides arising from hydrolysis of meat proteins in dry fermented sausages. Meat Sci. 2015, 104, 20–29. [Google Scholar] [CrossRef]
- Mora, L.; Escudero, E.; Toldrá, F. Characterization of the peptide profile in Spanish Teruel, Italian Parma and Belgian dry-cured hams and its potential bioactivity. Food Res. Int. 2016, 89, 638–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Guo, M.; Wang, Q.; Dong, J.; Lu, S.; Lyu, B.; Ma, X. Antioxidant activities of peptides derived from mutton ham, Xuanwei ham and Jinhua ham. Food Res. Int. 2021, 142, 110195. [Google Scholar] [CrossRef] [PubMed]
- Paolella, S.; Falavigna, C.; Faccini, A.; Virgili, R.; Sforza, S.; Dall’Asta, C.; Galaverna, G. Effect of dry-cured ham maturation time on simulated gastrointestinal digestion: Characterization of the released peptide fraction. Food Res. Int. 2015, 67, 136–144. [Google Scholar] [CrossRef]
- Martini, S.; Conte, A.; Tagliazucchi, D. Comparative peptidomic profile and bioactivities of cooked beef, pork, chicken and turkey meat after in vitro gastro-intestinal digestion. J. Proteom. 2019, 208, 103500. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, R.; Munekata, P.E.; Agregan, R.; Lorenzo, J.M. Effect of commercial starter cultures on free amino acid, biogenic amine and free fatty acid contents in dry-cured foal sausage. LWT-Food Sci. Technol. 2016, 71, 47–53. [Google Scholar] [CrossRef]
- Pogorzelska-Nowicka, E.; Atanasov, A.G.; Horbańczuk, J.; Wierzbicka, A. Bioactive compounds in functional meat products. Molecules 2018, 23, 307. [Google Scholar] [CrossRef] [Green Version]
- Kęska, P.; Stadnik, J.; Wójciak, K.M.; Neffe-Skocińska, K. Physico-chemical and proteolytic changes during cold storage of dry-cured pork loins with probiotic strains of LAB. Int. J. Food Sci. Technol. 2020, 55, 1069–1079. [Google Scholar] [CrossRef]
- Toldrá, F.; Gallego, M.; Reig, M.; Aristoy, M.C.; Mora, L. Bioactive peptides generated in the processing of dry-cured ham. Food Chem. 2020, 321, 126689. [Google Scholar] [CrossRef]
- Xing, L.; Li, G.; Toldrá, F.; Zhang, W. The physiological activity of bioactive peptides obtained from meat and meat by-products. Adv. Food Nutr. Res. 2021, 97, 147–185. [Google Scholar]
- Fernández, M.; Benito, M.J.; Martín, A.; Casquete, R.; Córdoba, J.J.; Córdoba, M.G. Influence of starter culture and a protease on the generation of ACE-inhibitory and antioxidant bioactive nitrogen compounds in Iberian dry-fermented sausage “salchichón”. Heliyon 2016, 2, e00093. [Google Scholar] [CrossRef] [Green Version]
- Champagne, C.P.; Ross, R.P.; Saarela, M.; Hansen, K.F.; Charalampopoulos, D. Recommendations for the viability assessment of probiotics as concentrated cultures and in food matrices. Int. J. Food Microbiol. 2011, 149, 185–193. [Google Scholar] [CrossRef]
- Libera, J.; Karwowska, M.; Stasiak, D.M.; Dolatowski, Z.J. Microbiological and physicochemical properties of dry-cured neck inoculated with probiotic of Bifidobacterium animalis ssp. lactis BB-12. Int. J. Food Sci. Technol. 2015, 50, 1560–1566. [Google Scholar] [CrossRef]
- Sionek, B.; Kołożyn-Krajewska, D.; Pasternok, I. Przeżywalność bakterii o właściwościach probiotycznych w kiełbasach surowo dojrzewających w czasie chłodniczego przechowywania. Żywność Nauka Technol. Jakość 2014, 21, 103–113. [Google Scholar]
- Stadnik, J.; Dolatowski, Z.J. Biogenic amines content during extended ageing of dry-cured pork loins inoculated with probiotics. Meat Sci. 2012, 91, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Neffe-Skocińska, K.; Okoń, A.; Kołożyn-Krajewska, D.; Dolatowski, Z. Amino acid profile and sensory characteristics of dry fermented pork loins produced with a mixture of probiotic starter cultures. J. Sci. Food Agric. 2017, 97, 2953–2960. [Google Scholar] [CrossRef]
- Wójciak, K.M.; Libera, J.; Stasiak, D.M.; Kołożyn-Krajewska, D. Technological aspect of Lactobacillus acidophilus Bauer, Bifidobacterium animalis BB-12 and Lactobacillus rhamnosus LOCK900 use in dry-fermented pork neck and sausage. J. Food Process. Preserv. 2017, 41, e12965. [Google Scholar] [CrossRef]
- Kęska, P.; Stadnik, J. Dipeptidyl Peptidase IV Inhibitory Peptides Generated in Dry-Cured Pork Loin during Aging and Gastrointestinal Digestion. Nutrients 2022, 14, 770. [Google Scholar] [CrossRef]
- Uniprot. Available online: https://www.uniprot.org/help/uniprotkb (accessed on 30 October 2021).
- De Almeida, M.A.; Saldaña, E.; da Silva Pinto, J.S.; Palacios, J.; Contreras-Castillo, C.J.; Sentandreu, M.A.; Fadda, S.G. A peptidomic approach of meat protein degradation in a low-sodium fermented sausage model using autochthonous starter cultures. Food Res. Int. 2018, 109, 368–379. [Google Scholar] [CrossRef]
- Sayd, T.; Dufour, C.; Chambon, C.; Buffière, C.; Remond, D.; Sante-Lhoutellier, V. Combined in vivo and in silico approaches for predicting the release of bioactive peptides from meat digestion. Food Chem. 2018, 249, 111–118. [Google Scholar] [CrossRef]
- Okoń, A.; Dolatowski, Z.J. Proteoliza białek w wędlinach surowo dojrzewających z udziałem szczepu probiotycznego Lactobacillus casei ŁOCK 0900. Żywność Nauka Technol. Jakość 2012, 19, 138–151. [Google Scholar]
- Okoń, A.; Dolatowski, Z.J. Effect of probiotic bacteria on free amino acid profile and sensory traits of raw-ripening pork sirloin during storage. Żywność Nauka Technol. Jakość 2014, 3, 92–107. [Google Scholar] [CrossRef]
- Okoń, A.; Stadnik, J.; Dolatowski, Z.J. Effect of Lactobacillus acidophilus Bauer and Bifidobacterium animalis ssp. Lactis BB12 on proteolytic changes in dry-cured loins. Food Sci. Biotechnol. 2017, 26, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Stadnik, J.; Dolatowski, Z.J. Changes in selected parameters related to proteolysis during ageing of dry-cured pork loins inoculated with probiotics. Food Chem. 2013, 139, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Stadnik, J.; Stasiak, D.M.; Dolatowski, Z.J. Proteolysis in dry-aged loins manufactured with sonicated pork and inoculated with Lactobacillus casei ŁOCK 0900 probiotic strain. Int. J. Food Sci. Technol. 2014, 49, 2578–2584. [Google Scholar] [CrossRef]
- Kęska, P.; Stadnik, J. Stability of antiradical activity of protein extracts and hydrolysates from dry-cured pork loins with probiotic strains of LAB. Nutrients 2018, 10, 521. [Google Scholar] [CrossRef] [Green Version]
- Kęska, P.; Stadnik, J. Ageing-time dependent changes of angiotensin I-converting enzyme-inhibiting activity of protein hydrolysates obtained from dry-cured pork loins inoculated with probiotic lactic acid bacteria. Int. J. Pept. Res. Ther. 2019, 25, 1173–1185. [Google Scholar] [CrossRef] [Green Version]
- Fontoura, R.; Daroit, D.J.; Corrêa, A.P.F.; Moresco, K.S.; Santi, L.; Beys-da-Silva, W.O.; Brandelli, A. Characterization of a novel antioxidant peptide from feather keratin hydrolysates. New Biotechnol. 2019, 49, 71–76. [Google Scholar] [CrossRef]
- Sinkiewicz, I.; Staroszczyk, H.; Śliwińska, A. Solubilization of keratins and functional properties of their isolates and hydrolysates. J. Food Biochem. 2018, 42, e12494. [Google Scholar] [CrossRef]
- Taraszkiewicz, A.; Sinkiewicz, I.; Sommer, A.; Dąbrowska, M.; Staroszczyk, H. Prediction of Bioactive Peptides from Chicken Feather and Pig Hair Keratins Using in Silico Analysis Based on Fragmentomic Approach. Curr. Pharm. Des. 2022, 28, 841–851. [Google Scholar] [CrossRef]
- Alahyaribeik, S.; Nazarpour, M. Effects of Bioactive Peptides Derived from Feather Keratin on Small Intestinal Function, Meat Quality, And Performance of Broiler Chicks; Research Square: Durham, UK, 2022. [Google Scholar] [CrossRef]
- López-Pedrouso, M.; Borrajo, P.; Amarowicz, R.; Lorenzo, J.M.; Franco, D. Peptidomic analysis of antioxidant peptides from porcine liver hydrolysates using SWATH-MS. J. Proteom. 2021, 232, 104037. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).