
Effect of PEMF on Muscle Oxygenation during Cycling: A Single-Blind Controlled Pilot Study


 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Design
2.2. Experimental Procedures
2.3. Data Analysis
2.4. Statistical Analysis
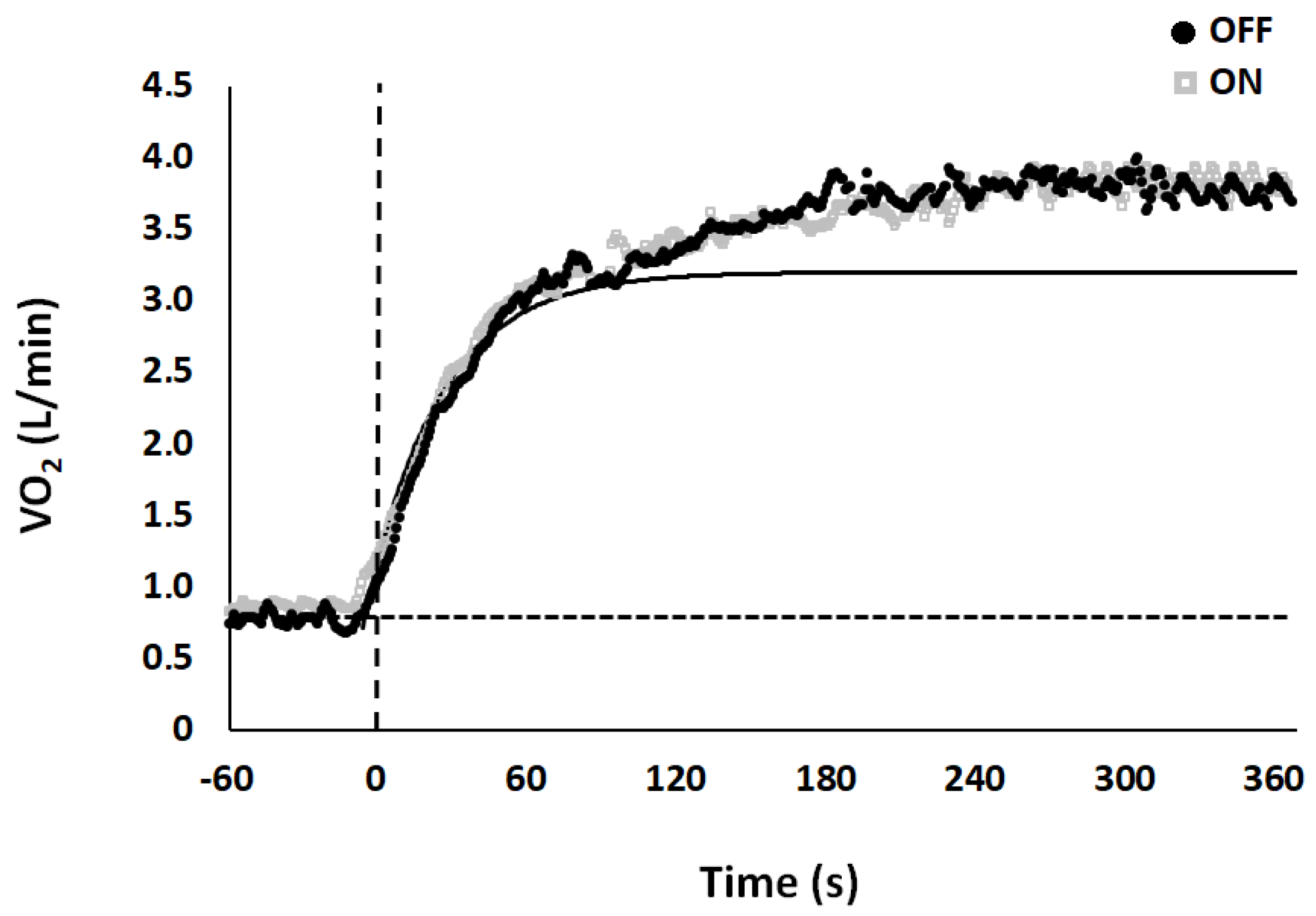
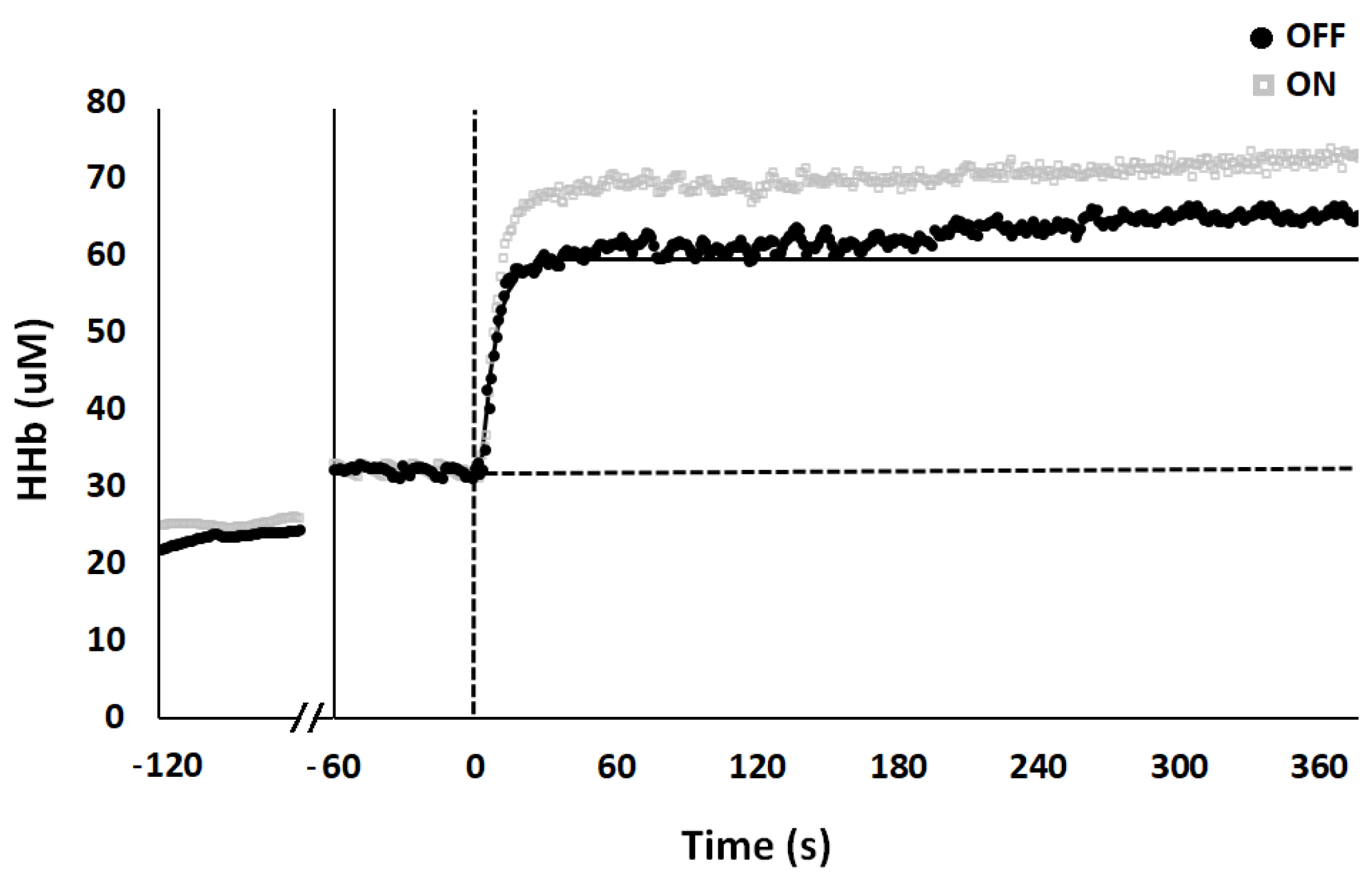
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pesce, M.; Patruno, A.; Speranza, L.; Reale, M. Extremely low frequency electromagnetic field and wound healing: Implication of cytokines as biological mediators. Eur. Cytokine Netw. 2013. [Google Scholar] [CrossRef]
- Kubat, N.J.; Moffett, J.; Fray, L.M. Effect of pulsed electromagnetic field treatment on programmed resolution of inflammation pathway markers in human cells in culture. J. Inflamm. Res. 2015. [Google Scholar] [CrossRef]
- Jeon, H.S.; Kang, S.Y.; Park, J.H.; Lee, H.S. Effects of pulsed electromagnetic field therapy on delayed-onset muscle soreness in biceps brachii. Phys. Ther. Sport 2015. [Google Scholar] [CrossRef]
- Smith, T.L.; Wong-Gibbons, D.; Maultsby, J. Microcirculatory effects of pulsed electromagnetic fields. J. Orthop. Res. 2004. [Google Scholar] [CrossRef]
- Roland, D.; Ferder, M.; Kothuru, R.; Faierman, T.; Strauch, B. Effects of pulsed magnetic energy on a microsurgically transferred vessel. Plast. Reconstr. Surg. 2000, 105, 1371–1374. [Google Scholar] [CrossRef] [PubMed]
- Muehsam, D.; Lalezari, P.; Lekhraj, R.; Abruzzo, P.; Bolotta, A.; Marini, M.; Bersani, F.; Aicardi, G.; Pilla, A.; Casper, D. Non-Thermal Radio Frequency and Static Magnetic Fields Increase Rate of Hemoglobin Deoxygenation in a Cell-Free Preparation. PLoS ONE 2013. [Google Scholar] [CrossRef]
- Pall, M.L. Electromagnetic fields act via activation of voltage-gated calcium channels to produce beneficial or adverse effects. J. Cell. Mol. Med. 2013, 17, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Milkiewicz, M.; Brown, M.D.; Egginton, S.; Hudlicka, O. Association between shear stress, angiogenesis, and VEGF in skeletal muscles in vivo. Microcirculation 2001. [Google Scholar] [CrossRef]
- Rikk, J.; Finn, K.J.; Liziczai, I.; Radák, Z.; Bori, Z.; Ihász, F. Influence of pulsing electromagnetic field therapy on resting blood pressure in aging adults. Electromagn. Biol. Med. 2013, 32, 165–172. [Google Scholar] [CrossRef]
- Mayrovitz, H.; Wounds, P.L.-U. Effects of pulsed electromagnetic fields on skin microvascular blood perfusion. Wounds 1992, 4, 197–202. [Google Scholar]
- Mayrovitz, H.N.; Larsen, P.B. A Preliminary Study to Evaluate the Effect of Pulsed Radio Frequency Field Treatment on Lower Extremity Peri-Ulcer Skin Microcirculation of Diabetic Patients. Wounds 1995, 7, 90–93. [Google Scholar]
- Kwan, R.L.C.; Wong, W.C.; Yip, S.L.; Chan, K.L.; Zheng, Y.P.; Cheing, G.L.Y. Pulsed electromagnetic field therapy promotes healing and microcirculation of chronic diabetic foot ulcers. Adv. Ski. Wound Care 2015. [Google Scholar] [CrossRef]
- Burnley, M.; Jones, A.M. Oxygen uptake kinetics as a determinant of sports performance. Eur. J. Sport Sci. 2007, 7, 63–79. [Google Scholar] [CrossRef]
- Koppo, K.; Bouckaert, J.; Jones, A.M. Effects of Training Status and Exercise Intensity on Phase II VO 2 Kinetics. Med. Sci. Sports Exerc. 2004. [Google Scholar] [CrossRef] [PubMed]
- Hirai, D.M.; Copp, S.W.; Holdsworth, C.T.; Ferguson, S.K.; McCullough, D.J.; Behnke, B.J.; Musch, T.I.; Poole, D.C. Skeletal muscle microvascular oxygenation dynamics in heart failure: Exercise training and nitric oxide-mediated function. Am. J. Physiol. Hear. Circ. Physiol. 2014, 306. [Google Scholar] [CrossRef] [PubMed]
- Grassi, B.; Quaresima, V. Near-infrared spectroscopy and skeletal muscle oxidative function in vivo in health and disease: A review from an exercise physiology perspective. J. Biomed. Opt. 2016, 21, 091313. [Google Scholar] [CrossRef] [PubMed]
- Piras, A.; Campa, F.; Toselli, S.; Di Michele, R.; Raffi, M. Physiological responses to partial-body cryotherapy performed during a concurrent strength and endurance session. Appl. Physiol. Nutr. Metab. 2019, 44, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Campa, F.; Piras, A.; Raffi, M.; Trofè, A.; Perazzolo, M.; Mascherini, G.; Toselli, S. The effects of dehydration on metabolic and neuromuscular functionality during cycling. Int. J. Environ. Res. Public Health 2020, 17, 1161. [Google Scholar] [CrossRef] [PubMed]
- Piras, A.; Cortesi, M.; Campa, F.; Perazzolo, M.; Gatta, G. Recovery time profiling after short-, middle-and long-distance swimming performance. J. Strength Cond. Res. 2019, 33, 1408–1415. [Google Scholar] [CrossRef] [PubMed]
- Piras, A.; Gatta, G. Evaluation of the effectiveness of compression garments on autonomic nervous system recovery after exercise. J. Strength Cond. Res. 2017, 31, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Mancini, D.M.; Bolinger, L.; Li, H.; Kendrick, K.; Chance, B.; Wilson, J.R. Validation of near-infrared spectroscopy in humans. J. Appl. Physiol. 1994. [Google Scholar] [CrossRef]
- Belardinelli, R.; Barstow, T.J.; Porszasz, J.; Wasserman, K. Skeletal muscle oxygenation during constant work rate exercise. Med. Sci. Sport. Exerc. 1995, 27, 512–519. [Google Scholar] [CrossRef]
- Niwayama, M.; Lin, L.; Shao, J.; Kudo, N.; Yamamoto, K. Quantitative measurement of muscle hemoglobin oxygenation using near-infrared spectroscopy with correction for the influence of a subcutaneous fat layer. Rev. Sci. Instrum. 2000, 71, 4571–4575. [Google Scholar] [CrossRef]
- Walsh, S.; Diamond, D. Non-linear curve fitting using microsoft excel solver. Talanta 1995. [Google Scholar] [CrossRef]
- Brown, A.M. A non-linear regression analysis program for describing electrophysiological data with multiple functions using Microsoft Excel. Comput. Methods Programs Biomed. 2006, 82, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Kemmer, G.; Keller, S. Nonlinear least-squares data fitting in Excel spreadsheets. Nat. Protoc. 2010. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.G.; Marshall, S.W.; Batterham, A.M.; Hanin, J. Progressive statistics for studies in sports medicine and exercise science. Med. Sci. Sport. Exerc. 2009, 41, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Grassi, B. Regulation of oxygen consumption at exercise onset: Is it really controversial? Exerc. Sport Sci. Rev. 2001, 29, 134–138. [Google Scholar] [CrossRef]
- DiMenna, F.J.; Bailey, S.J.; Vanhatalo, A.; Chidnok, W.; Jones, A.M. Elevated baseline VO2 per se does not slow O2 uptake kinetics during work-to-work exercise transitions. J. Appl. Physiol. 2010, 109, 1148–1154. [Google Scholar] [CrossRef][Green Version]
- Murias, J.M.; Spencer, M.D.; Kowalchuk, J.M.; Paterson, D.H. Muscle deoxygenation to VO 2 relationship differs in young subjects with varying τvO 2. Eur. J. Appl. Physiol. 2011. [Google Scholar] [CrossRef]
- Grassi, B.; Gladden, L.B.; Samaja, M.; Stary, C.M.; Hogan, M.C. Faster adjustment of O2 delivery does not affect VO2 on-kinetics in isolated in situ canine muscle. J. Appl. Physiol. 1998, 85, 1394–1403. [Google Scholar] [CrossRef][Green Version]
- Murias, J.M.; Kowalchuk, J.M.; Peterson, D.H. Speeding of Vo2 kinetics with endurance training in old and young men is associated with improved matching of local O2 delivery to muscle O2 utilization. J. Appl. Physiol. 2010. [Google Scholar] [CrossRef] [PubMed]
- Diniz, P.; Soejima, K.; Ito, G. Nitric oxide mediates the effects of pulsed electromagnetic field stimulation on the osteoblast proliferation and differentiation. Nitric Oxide 2002, 7, 18–23. [Google Scholar] [CrossRef]
- Behnke, B.J.; Kindig, C.A.; Musch, T.I.; Koga, S.; Poole, D.C. Dynamics of microvascular oxygen pressure across the rest-exercise transition in rat skeletal muscle. Respir. Physiol. 2001, 126, 53–63. [Google Scholar] [CrossRef]
- Hirai, D.M.; Copp, S.W.; Ferguson, S.K.; Holdsworth, C.T.; McCullough, D.J.; Behnke, B.J.; Musch, T.I.; Poole, D.C. Exercise training and muscle microvascular oxygenation: Functional role of nitric oxide. J. Appl. Physiol. 2012, 113, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Cocksedge, S.P.; Breese, B.C.; Morgan, P.T.; Nogueira, L.; Thompson, C.; Wylie, L.J.; Jones, A.M.; Bailey, S.J. Influence of muscle oxygenation and nitrate-rich beetroot juice supplementation on O2 uptake kinetics and exercise tolerance. Nitric Oxide Biol. Chem. 2020, 99, 25–33. [Google Scholar] [CrossRef]
- Wilkerson, D.P.; Campbell, I.T.; Jones, A.M. Influence of nitric oxide synthase inhibition on pulmonary O2 uptake kinetics during supra-maximal exercise in humans. J. Physiol. 2004, 561, 623–635. [Google Scholar] [CrossRef]
- Sakurai, T.; Satake, A.; Sumi, S.; Inoue, K.; Miyakoshi, J. An Extremely Low Frequency Magnetic Field Attenuates Insulin Secretion from the Insulinoma Cell Line, RIN-m. Bioelectromagnetics 2004. [Google Scholar] [CrossRef]
- Ross, C.L.; Siriwardane, M.; Almeida-Porada, G.; Porada, C.D.; Brink, P.; Christ, G.J.; Harrison, B.S. The effect of low-frequency electromagnetic field on human bone marrow stem/progenitor cell differentiation. Stem Cell Res. 2015, 15, 96–108. [Google Scholar] [CrossRef]
- Breese, B.C.; Mcnarry, M.A.; Marwood, S.; Blackwell, J.R.; Bailey, S.J.; Jones, A.M. Beetroot juice supplementation speeds O2 uptake kinetics and improves exercise tolerance during severe-intensity exercise initiated from an elevated metabolic rate. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305. [Google Scholar] [CrossRef]
- Ferguson, S.K.; Hirai, D.M.; Copp, S.W.; Holdsworth, C.T.; Allen, J.D.; Jones, A.M.; Musch, T.I.; Poole, D.C. Effects of nitrate supplementation via beetroot juice on contracting rat skeletal muscle microvascular oxygen pressure dynamics. Respir. Physiol. Neurobiol. 2013, 187, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Krustrup, P.; Söderlund, K.; Mohr, M.; Bangsbo, J.; Arch, P.; Krustrup, P.; Söderlund, K.; Mohr, M. The slow com ponent of oxygen uptake during intense sub-maximal exercise in man is associated with additional fi bre recruitment. Pflügers Arch. 2004, 447, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Robergs, R.A.; Ghiasvand, F.; Parker, D. Biochemistry of exercise-induced metabolic acidosis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R502–R516. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Base (L/min) | Ss (L/min) | Ap (L/min) | TDp (sec) | τp (sec) | As (L/min) | TDs (sec) | τs (sec) | MRTp (sec) | MRTs (sec) | Sc (L/min) | Sc (%) | Gain (ml/min/kg) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OFF | 0.93 ± 0.00 | 3.45 ± 0.10 | 2.52 ± 0.10 | 19.15 ± 1.50 | 19.55 ± 1.50 | 0.54 ± 0.00 | 107.26 ± 5.90 | 93.78 ± 15.80 | 38.70 ± 1.80 | 201.04 ± 15.60 | 0.24 ± 0.00 | 6.24 ± 0.90 | 8.88 ± 0.20 |
| ON | 0.94 ± 0.00 | 3.46 ± 0.10 | 2.52 ± 0.10 | 18.44 ± 2.20 | 17.94 ± 1.60 | 0.51 ± 0.00 | 104.50 ± 10.10 | 90.21 ± 12.67 | 36.38 ± 2.60 | 194.71 ± 15.10 | 0.25 ± 0.00 | 6.26 ± 0.80 | 8.88 ± 0.20 |
| Base (µM) | Ss (µM) | Ap (µM) | TDp (sec) | τp (sec) | As (µM) | TDs (sec) | τs (sec) | MRTp (sec) | MRTs (sec) | Sc (µM) | Sc (%) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OFF | 33.34 ± 4.20 | 60.70 ± 4.10 | 27.36 ± 2.30 | 7.80 ± 1.10 | 8.10 ± 1.20 | 8.95 ± 1.80 | 97.56 ± 13.30 | 111.44 ± 17.10 | 15.90 ± 1.60 | 209.00 ± 17.10 | 1.99 ± 0.40 | 2.77 ± 0.40 |
| ON | 34.34 ± 4.70 | 67.58 ± 5.00 * | 33.40 ± 3.30 * | 4.90 ± 0.80 * | 5.66 ± 0.70 * | 8.43 ± 2.20 | 75.57 ± 11.70 * | 137.85 ± 19.40 | 10.56 ± 1.10 * | 213.42 ± 25.90 | 2.56 ± 0.50 | 3.43 ± 0.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trofè, A.; Raffi, M.; Muehsam, D.; Meoni, A.; Campa, F.; Toselli, S.; Piras, A. Effect of PEMF on Muscle Oxygenation during Cycling: A Single-Blind Controlled Pilot Study. Appl. Sci. 2021, 11, 3624. https://doi.org/10.3390/app11083624
Trofè A, Raffi M, Muehsam D, Meoni A, Campa F, Toselli S, Piras A. Effect of PEMF on Muscle Oxygenation during Cycling: A Single-Blind Controlled Pilot Study. Applied Sciences. 2021; 11(8):3624. https://doi.org/10.3390/app11083624
Chicago/Turabian StyleTrofè, Aurelio, Milena Raffi, David Muehsam, Andrea Meoni, Francesco Campa, Stefania Toselli, and Alessandro Piras. 2021. "Effect of PEMF on Muscle Oxygenation during Cycling: A Single-Blind Controlled Pilot Study" Applied Sciences 11, no. 8: 3624. https://doi.org/10.3390/app11083624
APA StyleTrofè, A., Raffi, M., Muehsam, D., Meoni, A., Campa, F., Toselli, S., & Piras, A. (2021). Effect of PEMF on Muscle Oxygenation during Cycling: A Single-Blind Controlled Pilot Study. Applied Sciences, 11(8), 3624. https://doi.org/10.3390/app11083624