
Time Course of Sleep Inertia Dissipation in Memory Tasks

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Design and Procedure
2.3. Tasks
2.4. Data Analysis
3. Results
3.1. PSG Data
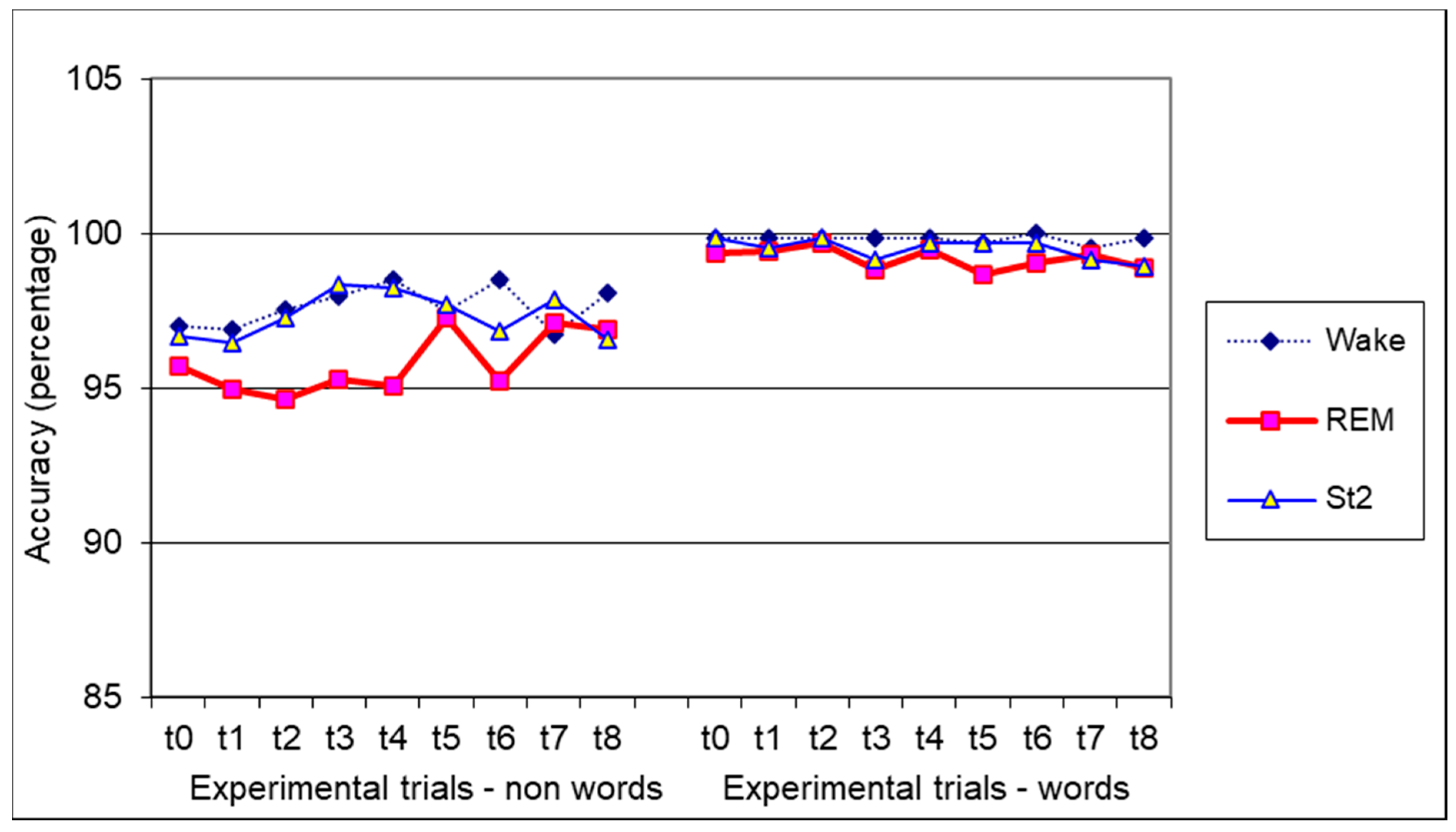
3.2. Lexical Decision Task
3.3. Episodic Learning Task
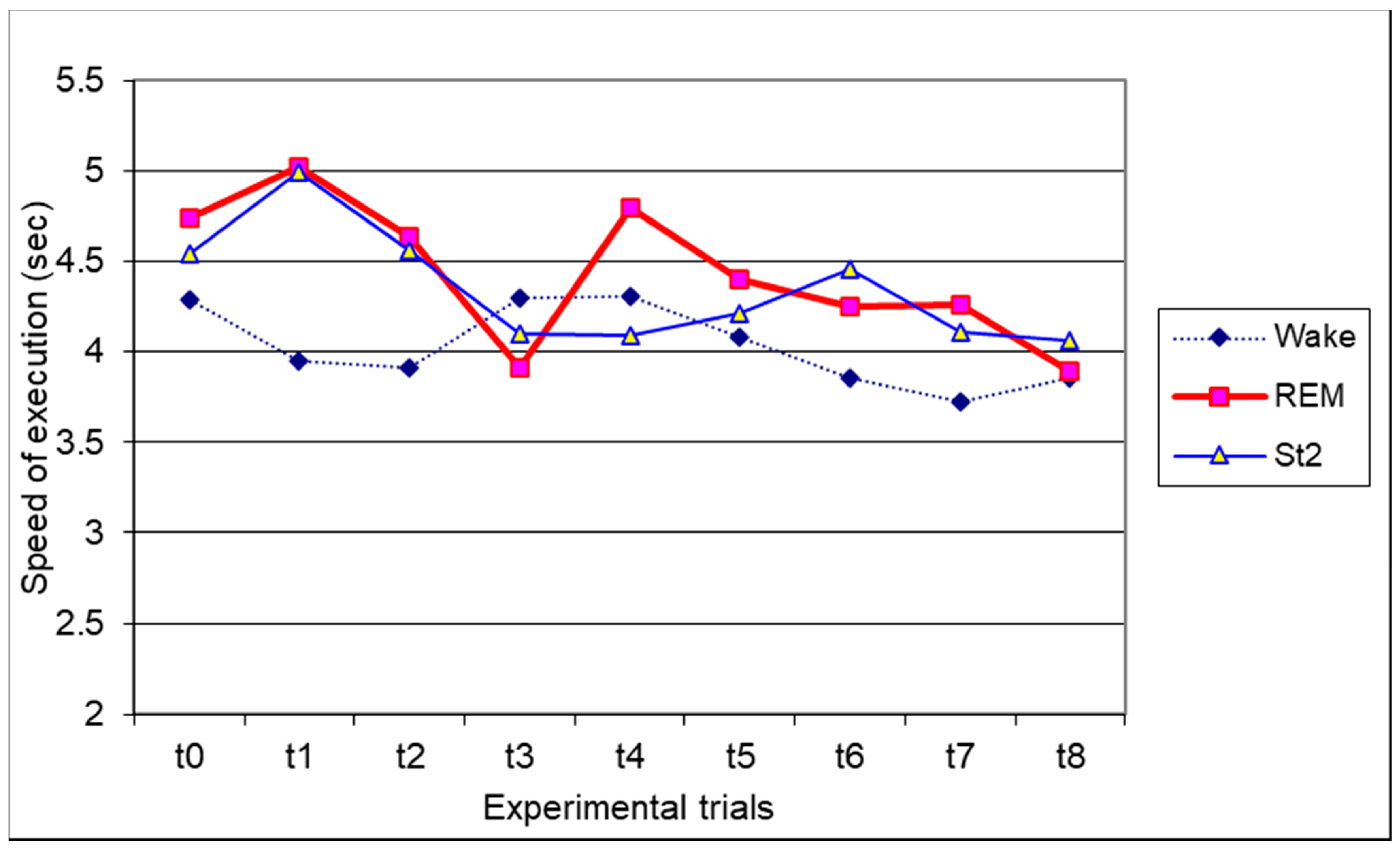
3.4. Finger Tapping Task
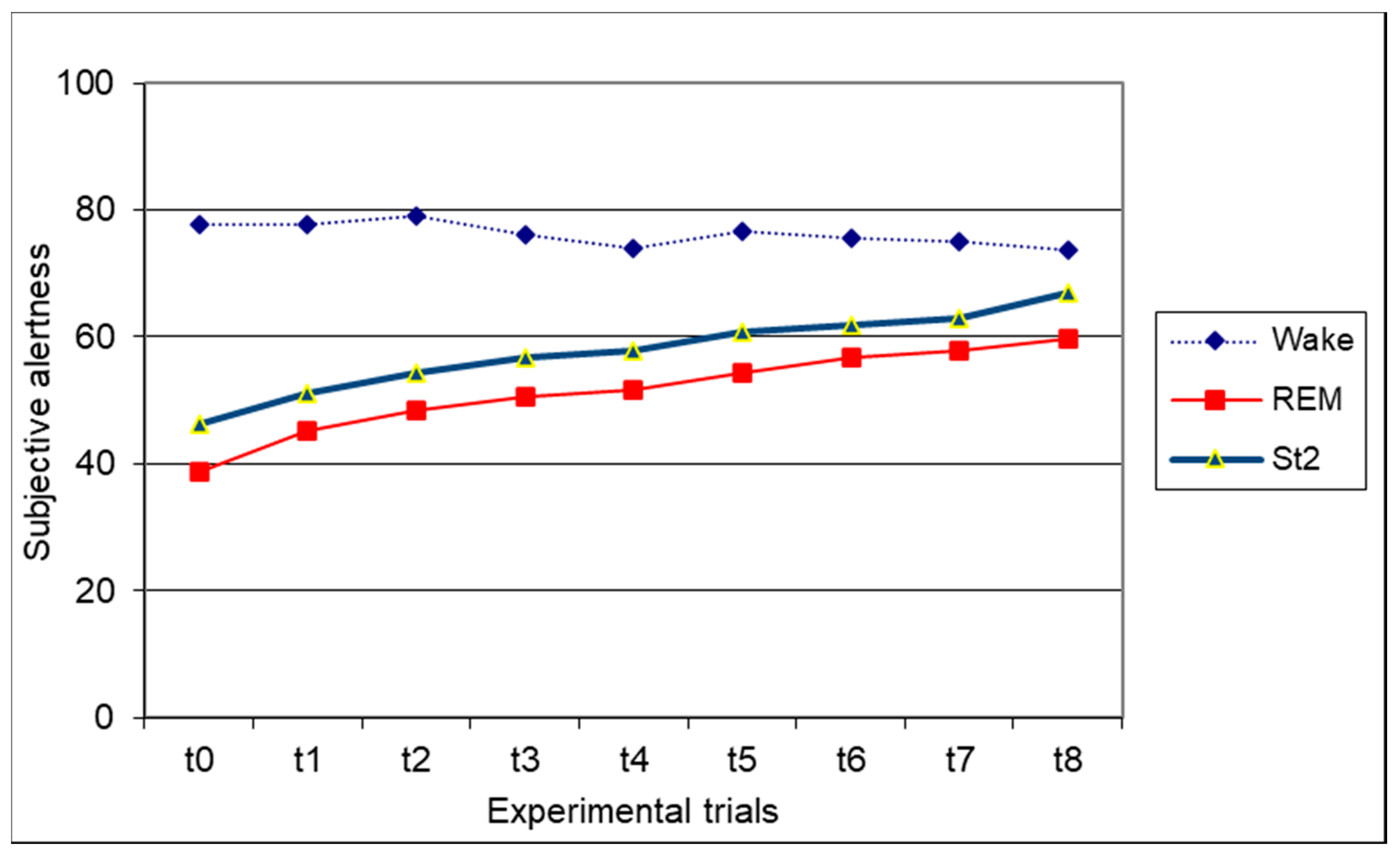
3.5. Subjective Alertness
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Broughton, R.J. Sleep disorders: Disorders or arousal? Science 1968, 159, 1070–1078. [Google Scholar] [CrossRef] [PubMed]
- Association of Sleep Disorders Centers. Diagnostic Classification of Sleep and Arousal Disorders, 1st Ed; Prepared by the Sleep Disorders Classification Committee, H.P.; Roffwarg, Chairman. Sleep 1979, 2, 1–137. [Google Scholar] [CrossRef]
- Lubin, A.; Hord, D.; Tracy, M.L.; Johnson, L.C. Effects of exercise, bedrest and napping on performance decrement during 40 hours. Psychophysiology 1976, 13, 334–339. [Google Scholar] [CrossRef]
- Tassi, P.; Muzet, A. Sleep inertia. Sleep Med. Rev. 2000, 4, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Trotti, L.M. Waking up is the hardest thing I do all day: Sleep inertia and sleep drunkenness. Sleep Med. Rev. 2017, 35, 76–84. [Google Scholar] [CrossRef]
- Hartman, B.O.; Langdon, D.E.; McKenzie, R.E. Performance on Sudden Awakening; USAF School of Aerospace Medicine Report: San Antonio, TX, USA, 1961. [Google Scholar]
- Hilditch, C.J.; McHill, A.W. Sleep Inertia: Current insights. Nat. Sci. Sleep 2019, 11, 155–165. [Google Scholar] [CrossRef] [PubMed]
- McHill, A.; Hull, J.; Czeisler, C.; Klerman, E. The effect of chronic sleep restriction and prior sleep duration on sleep inertia measured using cognitive performance. Sleep Med. 2017, 40, 163. [Google Scholar] [CrossRef]
- Peter-Derex, L.; Magnin, M.; Bastuji, H. Heterogeneity of arousals in human sleep: A stereo-electroencephalographic study. NeuroImage 2015, 123, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Hilditch, C.; Centofanti, S.; Dorrian, J.; Banks, S. A 30-minute, but not a 10-minute nighttime nap is associated with sleep inertia. Sleep 2016, 39, 675. [Google Scholar] [CrossRef] [PubMed]
- Marzano, C.; Ferrara, M.; Moroni, F.; De Gennaro, L. Electroencephalographic sleep inertia of the awakening brain. Neuroscience 2010, 176, 308–317. [Google Scholar] [CrossRef]
- Gorgoni, M.; Ferrara, M.; D’Atri, A.; Lauri, G.; Scarpelli, S.; Truglia, I.; De Gennaro, L. EEG topography during sleep inertia upon awakening after a period of increased homeostatic sleep pressure. Sleep Med. 2015, 16, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Scheer, F.A.; Shea, T.J.; Hilton, M.F.; Shea, S.A. An endogenous circadian rhythm in sleep inertia results in greatest cognitive impairment upon awakening during the biological night. J. Biol. Rhythm. 2008, 23, 353–361. [Google Scholar] [CrossRef]
- Ritchie, H.K.; Burke, T.M.; Dear, T.B.; McHill, A.W.; Axelsson, J.; Wright, K.P.J. Impact of sleep inertia on visual selective attention for rare targets and the influence of chronotype. J. Sleep Res. 2017, 26, 551–558. [Google Scholar] [CrossRef]
- Hilditch, C.J.; Dorrian, J.; Banks, S. Time to wake up: Reactive countermeasures to sleep inertia. Ind. Health 2016, 54, 528–541. [Google Scholar] [CrossRef]
- Ferrara, M.; De Gennaro, L.; Bertini, M. Time-course of sleep inertia upon awakening from nighttime sleep with different sleep homeostasis conditions. Aviat. Space Environ. Med. 2000, 71, 225–229. [Google Scholar]
- Wertz, A.T.; Ronda, J.M.; Czeisler, C.A.; Wright, K.P.J. Effects of sleep inertia on cognition. JAMA 2006, 295, 163–164. [Google Scholar] [CrossRef]
- Seminara, J.L.; Shavelson, R.J. Effectivness of space crew performance subsequent to sudden sleep arousal. Aerosp. Med. 1969, 40, 723–727. [Google Scholar] [PubMed]
- Ferrara, M.; De Gennaro, L.; Casagrande, M.; Bertini, M. Auditory arousal thresholds after selective slow-wave sleep deprivation. Clin. Neurophysiol. 1999, 110, 2148–2152. [Google Scholar] [CrossRef]
- Ferrara, M.; De Gennaro, L.; Casagrande, M.; Bertini, M. Selective slow-wave sleep deprivation and time-of-night effects on cognitive performance upon awakening. Psychophysiology 2000, 37, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Burke, T.M.; Scheer, F.A.; Ronda, J.M.; Czeisler, C.A.; Wright, K.P., Jr. Sleep inertia, sleep homeostatic and circadian influences on higher-order cognitive functions. J. Sleep Res. 2015, 24, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, M.; De Gennaro, L. The sleep inertia phenomenon during the sleep-wake transition. Theoretical and operational issues. Aviat. Space Environ. Med. 2000, 71, 843–848. [Google Scholar]
- Naitoh, P.; Kelly, T.; Babkoff, H. Sleep inertia, best time not to wake up? Chronobiol. Int. 1993, 10, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Dinges, D.F. Are you awake? Cognitive Performance and Reverie During the Hypnopompic State. In Sleep and Cognition; Bootzin, R., Kihlstrom, J., Schacter, D., Eds.; American Psychological Society: Washington, DC, USA, 1990; pp. 159–175. [Google Scholar]
- Naitoh, P.; Angus, R.G. Napping and human functioning during prolonged work. In Chronobiological, Behavioral and Medical Aspects of Napping; Dinges, D.F., Broughton, R.J., Eds.; Raven Press: New York, NY, USA, 1989; pp. 221–246. [Google Scholar]
- Tassi, P.; Nicolas, A.; Dewasmes, G.; Eschenlauer, R.; Ehrhart, J.; Salame, P.; Muzet, A.; Libert, J.P. Effects of noise on sleep inertia as a function of circadian placement of a one-hour nap. Percept. Mot. Ski. 1992, 75, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Hilditch, C.J.; Dorrian, J.; Banks, S. A review of short naps and sleep inertia: Do naps of 30 min or less really avoid sleep inertia and slow wave sleep? Sleep Med. Rev. 2017, 32, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Cavallero, C.; Versace, F. Stage at awakening, sleep inertia and performance. Sleep Res. Online 2003, 5, 89–97. [Google Scholar]
- Silva, E.J.; Duffy, J.F. Sleep inertia varies with circadian phase and sleep stage in older adults. Behav. Neurosci. 2008, 122, 928–935. [Google Scholar] [CrossRef]
- Dinges, D.F.; Orne, M.T.; Orne, E.C. Assessing performance upon abrupt awakening from naps during quasi-continuous operations. Behav. Res. Methods Instrum. Comput. 1985, 17, 37–45. [Google Scholar] [CrossRef]
- Akerstedt, T.; Gillberg, M. Effects of sleep deprivation on memory and sleep latencies in connection with repeated awakening from sleep. Psychophysiology 1979, 16, 49–52. [Google Scholar] [CrossRef]
- Rosa, R.R.; Bonnet, M.H.; Warm, J.S. Recovery of performance during sleep following sleep deprivation. Psychophysiology 1983, 20, 152–159. [Google Scholar] [CrossRef]
- Stones, M.J. Memory performance after arousal from different sleep stages. Br. J. Psychol. 1977, 68, 177–181. [Google Scholar] [CrossRef]
- Grosvenor, A.; Lack, L.C. The effect of sleep before or after learning on memory. Sleep 1984, 7, 155–167. [Google Scholar] [CrossRef]
- Stickgold, R.; Scott, L.; Rittenhouse, C.; Hobson, J.A. Sleep-induced changes in associative memory. J. Cogn. Neurosci. 1999, 11, 182–193. [Google Scholar] [CrossRef]
- Kolff, M.; Hofman, W.; Kerkhof, G.; Coenen, A. Effects on sleep-wake states on reaction times and priming effects in a semantic priming paradigm. Sleep Hypn. 2003, 5, 72–77. [Google Scholar]
- Kolff, M.; Hofman, W.; Kerkhof, G.; Coenen, A. The time course of sleep inertia in a semantic priming paradigm. Sleep Hypn. 2003, 5, 78–82. [Google Scholar]
- Tulving, E. How many memory systems are there? Am. Psychol. 1985, 40, 385–398. [Google Scholar] [CrossRef]
- Schacter, D.L.; Tulving, E. (Eds.) Memory Systems; The MIT Press: Cambridge, MA, USA, 1994. [Google Scholar]
- Jewett, M.E.; Wyatt, J.K.; Ritz-De Cecco, A.; Khalsa, S.B.; Dijk, D.J.; Czeisler, C.A. Time course of sleep inertia dissipation in human performance and alertness. J. Sleep Res. 1999, 8, 1–8. [Google Scholar] [CrossRef]
- Ikeda, H.; Hayashi, M. The effect of self-awakening from nocturnal sleep on sleep inertia. Biol. Psychol. 2010, 83, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, H.; Hayashi, M. Effect of sleep inertia on switch cost and arousal level immediately after awakening from normal nocturnal sleep. Sleep Biol. Rhythm. 2008, 6, 120–125. [Google Scholar] [CrossRef]
- Matchock, R.L.; Mordkoff, J.T. Visual attention, reaction time, and self-reported alertness upon awakening from sleep bouts of varying lengths. Exp. Brain Res. 2007, 178, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Natale, V.; Alzani, A.; Cicogna, P.C. Cognitive efficiency and circadian typologies: A diurnal study. Personl. Individ. Differ. 2003, 35, 1089–1105. [Google Scholar] [CrossRef]
- Horne, J.; Ostberg, O. A self-assessment questionnaire to determine morningness-eveningness in human circadian rhythm. Int. J. Chronobiol. 1976, 4, 97–110. [Google Scholar]
- Goldberg, D.P.; Blackwell, B. Psychiatric illness in general practice: A detailed study using a new method of case identification. Br. Med. J. 1970, 2, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Violani, C.; Devoto, A.; Lucidi, F.; Lombardo, C.; Russo, P.M. Validity of a short insomnia questionnaire: The SQD. Brain Res. Bull. 2004, 63, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Spielberger, C.D.; Gorsuch, R.L.; Lushene, R.D. STAI Manual; Consulting Psychologist: Palto Alto, CA, USA, 1970. [Google Scholar]
- Zung, W.W. A Self-Rating Depression Scale. Arch. Gen. Psychiatry 1965, 12, 63–70. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh Inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Natale, V.; Leger, D.; Bayon, V.; Erbacci, A.; Tonetti, L.; Fabbri, M.; Martoni, M. The Consensus SleepDiary: Quantitative criteria for primary insomnia diagnosis. Psychosom. Med. 2015, 77, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Rechtschaffen, A.; Kales, A. A Manual of Standardized Terminology, Techniques and Scoring System for Sleep Stages of Human Subjects; National Institute of Health: Washington, DC, USA, 1968.
- Monk, T.A. A Visual Analogue Scale technique to measure global vigor and affect. Psychiatry Res. 1989, 27, 89–99. [Google Scholar] [CrossRef]
- Wilkinson, R.T.; Stretton, M. Performance after awakening at different times of night. Psychon. Sci. 1971, 23, 283–285. [Google Scholar] [CrossRef]
- Tassi, P.; Bonnefond, A.; Engasser, O.; Hoeft, A.; Eschenlauer, R.; Muzet, A. EEG spectral power and cognitive performance during sleep inertia: The effect of normal sleep duration and partial sleep deprivation. Physiol. Behav. 2006, 87, 177–184. [Google Scholar] [CrossRef]
- Nyberg, L.; Tulving, E. Classifying human long-term memory: Evidence from converging dissociations. Eur. J. Cogn. Psychol. 1996, 8, 163–183. [Google Scholar] [CrossRef]
- Balkin, T.J.; Badia, P. Relationship between sleep inertia and sleepiness: Cumulative effects of four nights of sleep disruption/restriction on performance following abrupt nocturnal awakenings. Biol. Psychol. 1988, 27, 245–258. [Google Scholar] [CrossRef]
- Miccoli, L.; Versace, F.; Koterle, S.; Cavallero, C. Comparing sleep-loss sleepiness and sleep inertia: Lapses make the difference. Chronobiol. Int. 2008, 25, 725–744. [Google Scholar] [CrossRef]
- Achermann, P.; Werth, E.; Dijk, D.J.; Bobely, A.A. Time course of sleep inertia after nighttime and daytime sleep episodes. Arch. Ital. Biol. 1995, 134, 109–119. [Google Scholar] [PubMed]
- Folkard, S.; Åkerstedt, T. A three-process model of the regulation of alertness-sleepiness. In Sleep, Arousal, and Performance; Broughton, R.J., Ogilvie, R.D., Eds.; Birkhäuser: Boston, MA, USA, 1992; pp. 11–26. [Google Scholar]
- Rauchs, G.; Desgranges, B.; Foret, J.; Eustache, F. The relationships between memory systems and sleep stages. J. Sleep Res. 2005, 14, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Diekelmann, S.; Wilhelm, I.; Born, J. The whats and whens of sleep-dependent memory consolidation. Sleep Med. Rev. 2009, 13, 309–321. [Google Scholar] [CrossRef]
- Bastuji, H.; Perrin, F.; Garcia-Larrea, L. Event-related potentials during forced awakening: A tool for the study of acute sleep inertia. J. Sleep Res. 2003, 12, 189–206. [Google Scholar] [CrossRef] [PubMed]
- Vyazovskiy, V.V.; Cui, N.; Rodriguez, A.V.; Funk, C.; Cirelli, C.; Tononi, G. The Dynamics of Cortical Neuronal Activity in the First Minutes after Spontaneous Awakening in Rats and Mice. Sleep 2014, 37, 1337–1347. [Google Scholar] [CrossRef]
- Dorrian, S.; Lamond, N.; Dawson, D. The ability to self-monitor performance when fatigued. J. Sleep Res. 2000, 9, 137–144. [Google Scholar] [CrossRef]
- Monk, T.H.; Buysse, D.J.; Reynolds, C.F.; Berga, S.L.; Jarrett, D.B.; Begley, A.E.; Kupfer, D.J. Circadian rhythms in human performance and mood under constant conditions. J. Sleep Res. 1997, 6, 9–18. [Google Scholar] [CrossRef]
- Balkin, T.J.; Braun, A.R.; Wesensten, N.J.; Jeffries, K.; Varga, M.; Baldwin, P.; Belenky, G.; Herscovitch, P. The process of awakening: A PET study of regional brain activity patterns mediating the re-establishment of alertness and consciousness. Brain 2002, 125, 2308–2319. [Google Scholar] [CrossRef]
- Vallat, R.; Meunier, D.; Nicolas, A.; Ruby, P. Hard to wake up? The cerebral correlates of sleep inertia assessed using combined behavioral, EEG and fMRI measures. NeuroImage 2019, 184, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Massimini, M.; Ferrarelli, F.; Huber, R.; Esser, S.K.; Singh, H.; Tononi, G. Breakdown of cortical effective connectivity during sleep. Science 2005, 309, 2228–2232. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sleep Parameters | Stage 2 Sleep Condition | REM Sleep Condition | t17 | p |
|---|---|---|---|---|
| LOff (time) | 24 h:27 min | 24 h:33 m | 1.71 | ns |
| LOn (time) | 7 h:30 m | 7 h:34 m | 0.74 | ns |
| SOL (min) | 10.61 ± 8.39 | 13.55 ± 6.43 | 1.44 | ns |
| TIB (min) | 417.94 ± 20.98 | 431.39 ± 14.86 | 2.04 | ns |
| TST (min) | 372.68 ± 28.26 | 375.87 ± 18.39 | 0.42 | ns |
| SE | 89.12 ± 4.18 | 87.23 ± 4.69 | 1.50 | ns |
| Stage 1% | 2.00 ± 0.85 | 2.42 ± 1.03 | 1.68 | ns |
| Stage 2% | 45.63 ± 3.92 | 46.09 ± 4.77 | 0.51 | ns |
| SWS % | 20.66 ± 3.66 | 19.29 ± 3.62 | 1.95 | ns |
| REM % | 20.56 ± 3.04 | 19.54 ± 2.45 | 1.78 | ns |
| REM Latency (min) | 97.46 ± 40.41 | 106.61 ± 38.70 | 1.18 | ns |
| WASO (min) | 16.29 ± 11.96 | 25.61 ± 19.18 | 2.06 | ns |
| t0 | t1 | t2 | t3 | t4 | t5 | t6 | t7 | t8 | F8,136 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Wake | 553 | 587 | 572 | 578 | 593 | 613 | 582 | 623 | 611 | 4.02 § |
| REM | 691 | 637 | 648 | 603 | 594 | 665 | 608 | 635 | 646 | 3.12 * |
| St2 | 670 | 653 | 642 | 608 | 650 | 656 | 619 | 646 | 665 | 1.87 |
| F2,34 | 14.77 ^ | 5.80 ′ | 5.64 ′ | 1.07 | 3.51 # | 2.68 | 1.70 | 0.6 | 2.18 |
| t0 | t1 | t2 | t3 | t4 | t5 | t6 | t7 | t8 | F8,136 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Wake | 11.3 | 11.7 | 11.4 | 11.4 | 11.3 | 12.0 | 12.1 | 11.6 | 11.0 | 0.68 |
| REM | 9.8 | 10.7 | 10.6 | 10.2 | 9.9 | 9.7 | 10.9 | 10.2 | 10.1 | 0.89 |
| St2 | 9.7 | 10.7 | 11.2 | 10.5 | 10.4 | 10.1 | 10.3 | 10.1 | 10.8 | 1.22 |
| F2,34 | 3.37 * | 1.51 | 0.47 | 1.94 | 1.67 | 4.57 * | 3.96 * | 3.02 | 0.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Occhionero, M.; Fabbri, M.; Tonetti, L.; Martoni, M.; Natale, V. Time Course of Sleep Inertia Dissipation in Memory Tasks. Appl. Sci. 2021, 11, 3354. https://doi.org/10.3390/app11083354
Occhionero M, Fabbri M, Tonetti L, Martoni M, Natale V. Time Course of Sleep Inertia Dissipation in Memory Tasks. Applied Sciences. 2021; 11(8):3354. https://doi.org/10.3390/app11083354
Chicago/Turabian StyleOcchionero, Miranda, Marco Fabbri, Lorenzo Tonetti, Monica Martoni, and Vincenzo Natale. 2021. "Time Course of Sleep Inertia Dissipation in Memory Tasks" Applied Sciences 11, no. 8: 3354. https://doi.org/10.3390/app11083354
APA StyleOcchionero, M., Fabbri, M., Tonetti, L., Martoni, M., & Natale, V. (2021). Time Course of Sleep Inertia Dissipation in Memory Tasks. Applied Sciences, 11(8), 3354. https://doi.org/10.3390/app11083354