
Effects of Pulsed Electric Fields on Yeast with Prions and the Structure of Amyloid Fibrils

 ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
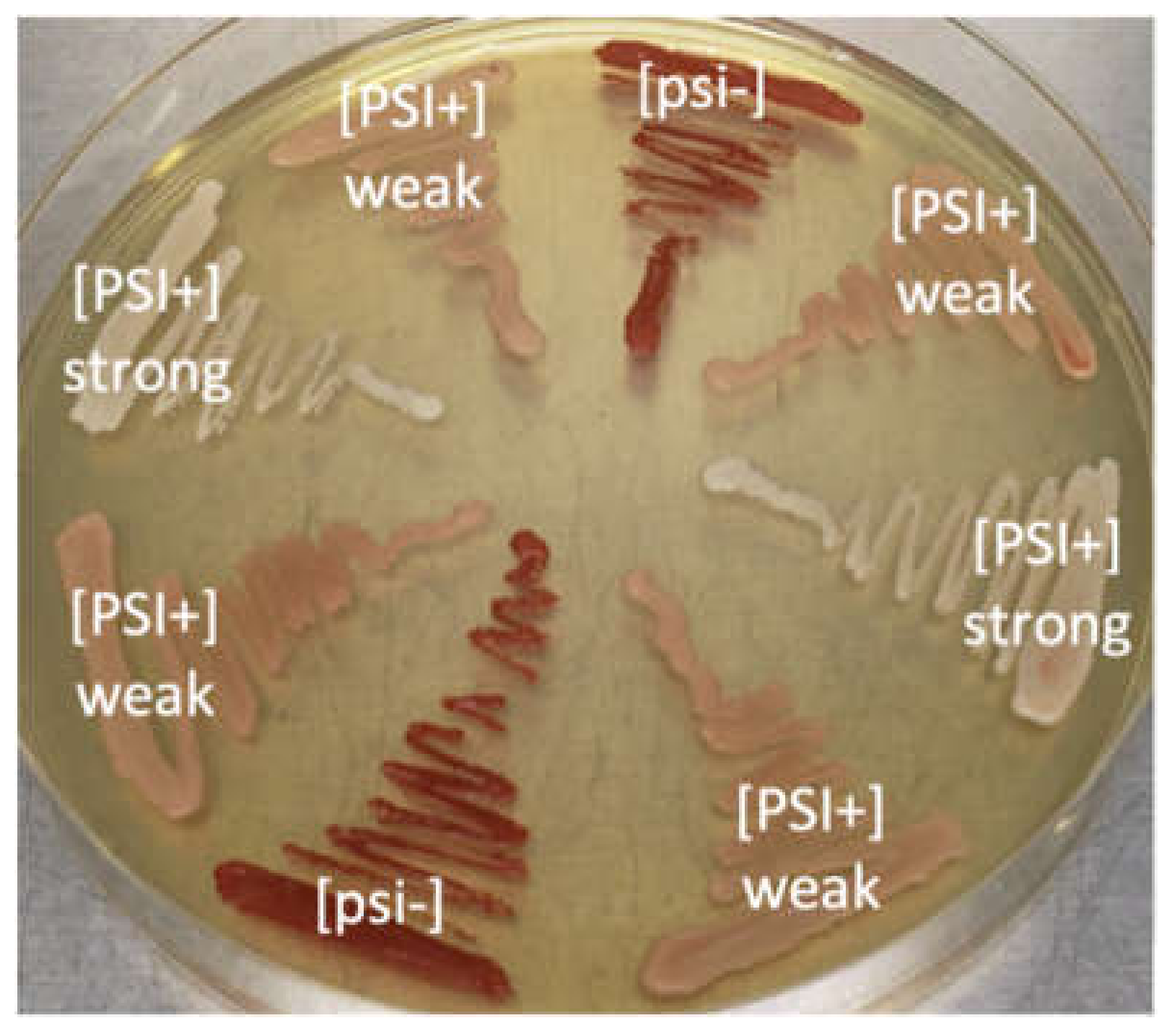
2.1. Yeast Strains. Saccharomyces Cerevisiae 74-D694
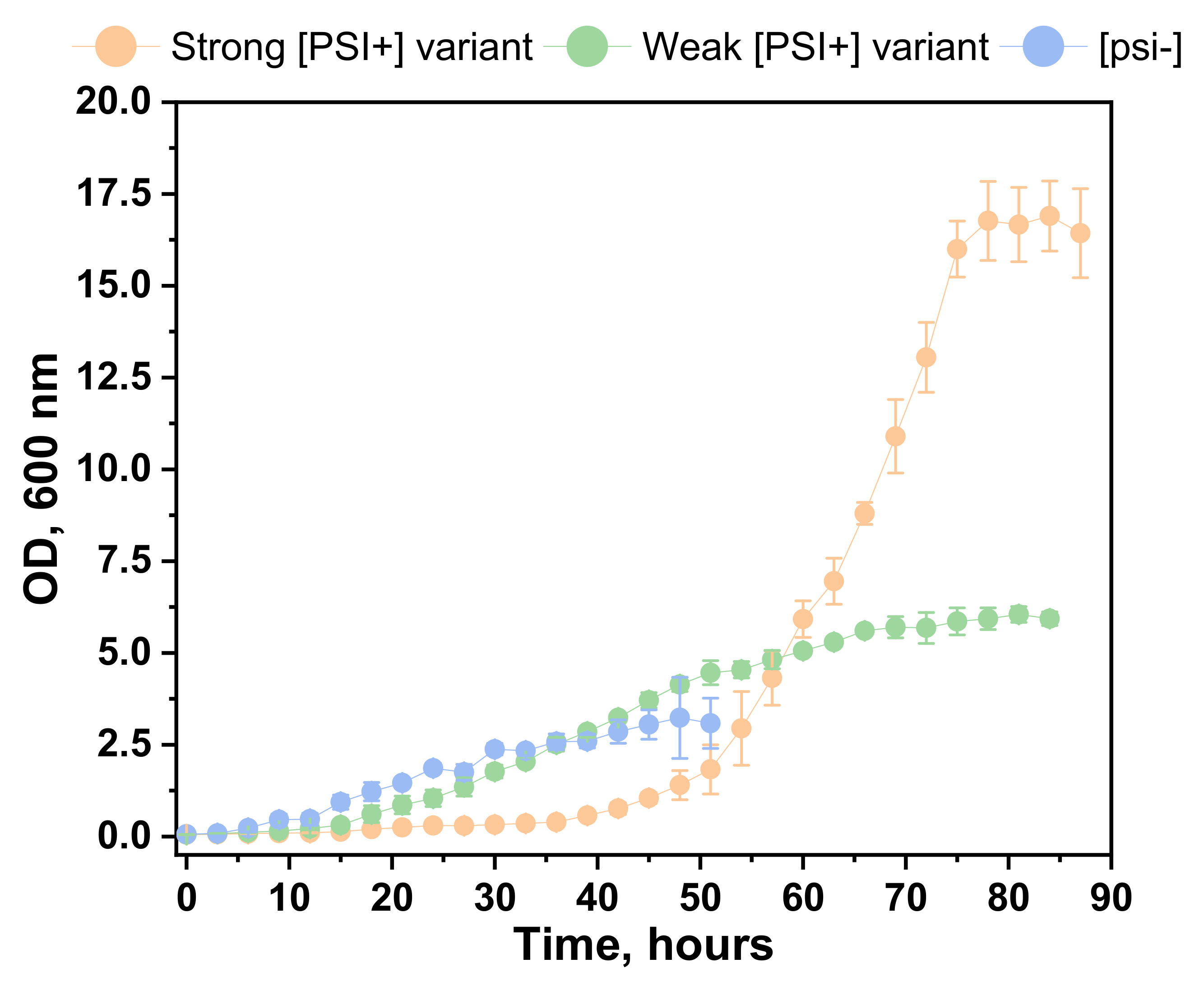
2.2. Cell Growth Dynamics
2.3. PEF Permeabilization Assay
2.4. Expression and Purification
2.5. Preparation of Sup35 Aggregates
2.6. Atomic Force Microscopy
2.7. Statistical Analysis
3. Results
4. Discussion
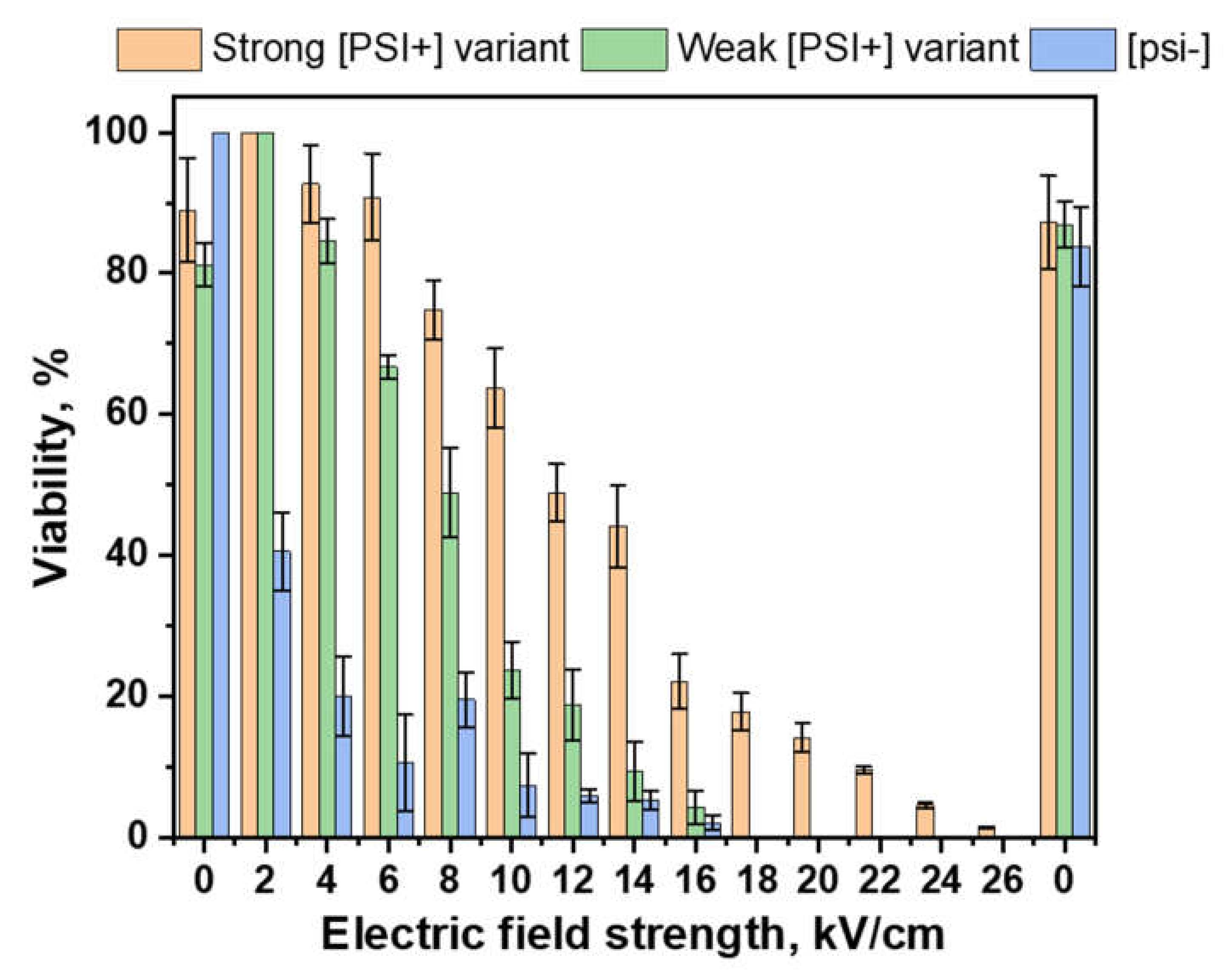
- Do [PSI+] prions increase PEF resistance in S. cerevisiae cells and how do different variants change this resistance? Our results show that S. cerevisiae cells containing [PSI+] prions exhibited extremely increased resistance to PEF treatment even in the case of weak prion variants (Figure 3). There is also a significant difference between survivability after PEF between strong and weak variants. Almost 10% viability was observed in the case of strong prions, even after applying 22 kV/cm, 1000 μs pulse duration for 5 pulses. Prions provided cells the ability to survive in such harsh treatment. Other authors agree that prions are related to stress resistance and improve cell viability in chronological aging [40,41]. Prions in the amyloid conformation are resistant to proteases, heat shock, and other inactivation methods [14]. Franzmann et al. showed that [PSI+] prion can regulate the phase separation of native Sup35 protein during stress [15]. In cell growth, Sup35 acts as a translation terminator. In the stress conditions, the prion and sensor domains act together to promote phase separation and to establish protective and reversible biomolecular condensates.
- Can PEF induce the appearance of prions in the cell population de novo or change the type of prion from weak to strong by producing free amyloid ends for protein polymerization? There is a hypothesis that prions in the cell population can appear de novo in cells experiencing stress conditions [41,42] and prion variants can modulate prion-dependent proteomes in response to environmental stress [43], producing a facilitated adaptation of the cells by enhancing survival in rapidly changing environments [44,45,46]. We hypothesized that PEF could act as a factor capable of disrupting cell homeostasis and lead to prion induction. In the case of the cells already containing prions, it could serve as an agent that generates free amyloid ends for the more effective intake of the Sup35 to the amyloids. Analysis of the [psi−] and weak [PSI+] colonies after PEF treatment showed no signs of showing new prions or shift from weak to strong variants. This is the possibility that the PEF itself is not capable of inducing prionization in yeast cells, which makes PEF safe for the biocontrol of yeasts without the opportunity to induce prionization.
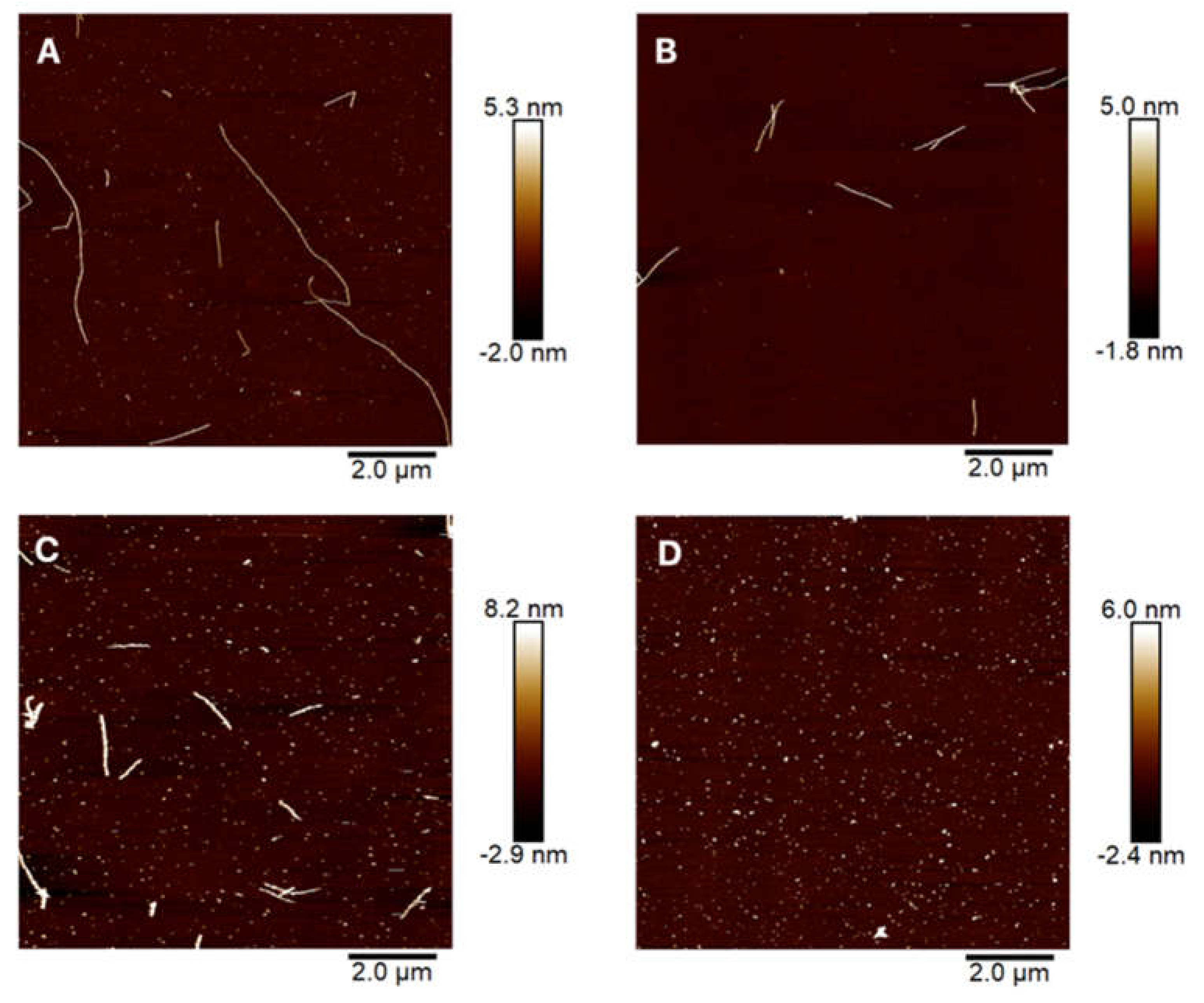
- Is it possible to disintegrate [PSI+] amyloids by applying PEF to the purified proteins? There is no covalent bond in the formation of the secondary and tertiary structure between the fibril assembling prion proteins. Stabilizing factors are electrostatic interactions, such as salt bridges and self-energy [28]. Therefore, we hypothesized that those interactions can easily be broken by applying PEF. The ability to use PEF for fibril disintegration could provide universal methods to disintegrate [PSI+] amyloid aggregates. In our research, atomic force microscopy revealed that amyloids of Sup35NM can be successfully disintegrated by using PEF (Figure 4). Even after applying 10 kV/cm, 50 μs pulse duration, and 10 pulses, we observed a length reduction in the amyloid fibrils as compared to the untreated ones. The longest fibril found in the samples was 1.45 µm long, which was around four times less than the amyloid from the control samples. This shows that, for the disintegration of the long fibrils into the smaller aggregates 10 kV/cm, 50 µs, and 10 pulses were enough. The diameter of the fibrils indicates that they belong to a strong prion variant. More than 80% of the strong prion amyloids are thin (~11.5 nm) and around 68% of the weak prions produce thick ~16 nm fibrils [47]. Our research showed that treating the fibrils with longer pulse results not only disintegrates fibrils but increases their diameter as well. The shift from 50 µs to 1000 µs resulted in the doubling of the fibril diameter. An electric field affects the charged groups of amino acids in the molecule, causing a change of the conformation for the fibril itself.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prusiner, S.B. Molecular biology of prion diseases. Science 1991, 252, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- Wickner, R.B.; Son, M.; Edskes, H.K. Prion variants of yeast are numerous, mutable, and segregate on growth, affecting prion pathogenesis, transmission barriers, and sensitivity to anti-prion systems. Viruses 2019, 11, 238. [Google Scholar] [CrossRef]
- Shewmaker, F.; McGlinchey, R.P.; Wickner, R.B. Structural insights into functional and pathological amyloid. J. Biol. Chem. 2011, 286, 16533–16540. [Google Scholar] [CrossRef] [PubMed]
- Wickner, R.B.; Edskes, H.K.; Son, M.; Bezsonov, E.E.; DeWilde, M.; Ducatez, M. Yeast Prions Compared to Functional Prions and Amyloids. J. Mol. Biol. 2018, 430, 3707–3719. [Google Scholar] [CrossRef] [PubMed]
- Bradley, M.E.; Liebman, S.W. The Sup35 domains required for maintenance of weak, strong or undifferentiated yeast [PSI+] prions. Mol. Microbiol. 2004, 51, 1649–1659. [Google Scholar] [CrossRef] [PubMed]
- Derkatch, I.L.; Chernoff, Y.O.; Kushnirov, V.V.; Inge-Vechtomov, S.G.; Liebman, S.W. Genesis and variability of [PSI] prion factors in Saccharomyces cerevisiae. Genetics 1996, 144, 1375–1386. [Google Scholar] [CrossRef] [PubMed]
- Sant’anna, R.; Fernández, M.R.; Batlle, C.; Navarro, S.; De Groot, N.S.; Serpell, L.; Ventura, S. Characterization of amyloid cores in prion domains. Sci. Rep. 2016, 6, srep34274. [Google Scholar] [CrossRef]
- Tuite, M.F.; Howard, M.J.; Xue, W.F. Dynamic prions revealed by magic. Chem. Biol. 2014, 21, 172–173. [Google Scholar] [CrossRef][Green Version]
- Vos, T.; Flaxman, A.D.; Naghavi, M.; Lozano, R.; Michaud, C.; Ezzati, M.; Shibuya, K.; Salomon, J.A.; Abdalla, S.; Aboyans, V.; et al. Years lived with disability (YLDs) for 1160 sequelae of 289 diseases and injuries 1990-2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2163–2196. [Google Scholar] [CrossRef]
- Frederick, K.K.; Debelouchina, G.T.; Kayatekin, C.; Dorminy, T.; Jacavone, A.C.; Griffin, R.G.; Lindquist, S. Distinct prion strains are defined by amyloid core structure and chaperone binding site dynamics. Chem. Biol. 2014, 21, 295–305. [Google Scholar] [CrossRef]
- Linden, R.; Martins, V.R.; Prado, M.A.M.; Cammarota, M.; Izquierdo, I.; Brentani, R.R. Physiology of the prion protein. Physiol. Rev. 2008, 88, 673–728. [Google Scholar] [CrossRef] [PubMed]
- Castle, A.R.; Gill, A.C. Physiological functions of the cellular prion protein. Front. Mol. Biosci. 2017, 4, 19. [Google Scholar] [CrossRef]
- Wulf, M.A.; Senatore, A.; Aguzzi, A. The biological function of the cellular prion protein: An update. BMC Biol. 2017, 15, 34. [Google Scholar] [CrossRef] [PubMed]
- Scheckel, C.; Aguzzi, A. Prions, prionoids and protein misfolding disorders. Nat. Rev. Genet. 2018, 19, 405–418. [Google Scholar] [CrossRef]
- Franzmann, T.M.; Jahnel, M.; Pozniakovsky, A.; Mahamid, J.; Holehouse, A.S.; Nüske, E.; Richter, D.; Baumeister, W.; Grill, S.W.; Pappu, R.V.; et al. Phase separation of a yeast prion protein promotes cellular fitness. Science 2018, 359, eaao5654. [Google Scholar] [CrossRef] [PubMed]
- Miklavčič, D.; Mali, B.; Kos, B.; Heller, R.; Serša, G. Electrochemotherapy: From the drawing board into medical practice. Biomed. Eng. Online 2014, 13, 29. [Google Scholar] [CrossRef] [PubMed]
- Mir, L.M. Therapeutic perspectives of in vivo cell electropermeabilization. Bioelectrochemistry 2001, 53, 1–10. [Google Scholar] [CrossRef]
- Jakštys, B.; Ruzgys, P.; Tamošiūnas, M.; Šatkauskas, S. Different Cell Viability Assays Reveal Inconsistent Results After Bleomycin Electrotransfer In Vitro. J. Membr. Biol. 2015, 248, 857–863. [Google Scholar] [CrossRef] [PubMed]
- De Backer, M.D.; Maes, D.; Vandoninck, S.; Logghe, M.; Contreras, R.; Luyten, W.H.M.L. Transformation of Candida albicans by electroporation. Yeast 1999, 15, 1609–1618. [Google Scholar] [CrossRef]
- Tee, K.L.; Grinham, J.; Othusitse, A.M.; González-Villanueva, M.; Johnson, A.O.; Wong, T.S. An Efficient Transformation Method for the Bioplastic-Producing “Knallgas” Bacterium Ralstonia eutropha H16. Biotechnol. J. 2017, 12, 1700081. [Google Scholar] [CrossRef]
- Holo, H.; Nes, I.F. High-frequency transformation, by electroporation, of Lactococcus lactis subsp. cremoris grown with glycine in osmotically stabilized media. Appl. Environ. Microbiol. 1989, 55, 3119–3123. [Google Scholar] [CrossRef]
- Chang, D.C.; Saunders, J.A.; Chassy, B.M.; Sowers, A.E. Guide to Electroporation and Electrofusion; Academic Press: Cambridge, MA, USA, 2012; ISBN 9780121680411. [Google Scholar]
- Mekid, H.; Mir, L.M. In vivo cell electrofusion. Biochim. Biophys. Acta-Gen. Subj. 2000, 1524, 118–130. [Google Scholar] [CrossRef]
- Rems, L.; Ušaj, M.; Kandušer, M.; Reberšek, M.; Miklavčič, D.; Pucihar, G. Cell electrofusion using nanosecond electric pulses. Sci. Rep. 2013, 3, 3382. [Google Scholar] [CrossRef]
- Kotnik, T.; Frey, W.; Sack, M.; Haberl Meglič, S.; Peterka, M.; Miklavčič, D. Electroporation-based applications in biotechnology. Trends Biotechnol. 2015, 33, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Toepfl, S.; Siemer, C.; Saldaña-Navarro, G.; Heinz, V. Overview of Pulsed Electric Fields Processing for Food. In Emerging Technologies for Food Processing; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Robin, A.; Golberg, A. Pulsed Electric Fields and Electroporation Technologies in Marine Macroalgae Biorefineries. In Handbook of Electroporation; Springer: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Guest, W.C.; Cashman, N.R.; Plotkin, S.S. Electrostatics in the stability and misfolding of the prion protein: Salt bridges, self energy, and solvation. Biochem. Cell Biol. 2010, 88, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Serio, T.R. [PIN + ]ing down the mechanism of prion appearance. FEMS Yeast Res. 2018, 18, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wickner, R.B.; Dyda, F.; Tycko, R. Amyloid of Rnq1p, the basis of the [PIN+] prion, has a parallel in-register β-sheet structure. Proc. Natl. Acad. Sci. USA 2008, 105, 2403–2408. [Google Scholar] [CrossRef]
- Derkatch, I.L.; Bradley, M.E.; Hong, J.Y.; Liebman, S.W. Prions Affect the Appearance of Other Prions: The Story of [PIN] University of Illinois at Chicago. Cell 2001, 106, 171–182. [Google Scholar] [CrossRef]
- Novickij, V.; Grainys, A.; Butkus, P.; Tolvaišienė, S.; Švedienė, J.; Paškevičius, A.; Novickij, J. High-frequency submicrosecond electroporator. Biotechnol. Biotechnol. Equip. 2016, 30, 607–613. [Google Scholar] [CrossRef]
- Espargaró, A.; Villar-Piqué, A.; Sabaté, R.; Ventura, S. Yeast prions form infectious amyloid inclusion bodies in bacteria. Microb. Cell Fact. 2012, 11, 89. [Google Scholar] [CrossRef] [PubMed]
- Studier, F.W. Protein production by auto-induction in high density shaking cultures. Protein Expr. Purif. 2005, 41, 207–234. [Google Scholar] [CrossRef] [PubMed]
- Tuite, M.F.; Cox, B.S. Propagation of yeast prions. Nat. Rev. Mol. Cell Biol. 2003, 4, 878–890. [Google Scholar] [CrossRef] [PubMed]
- Shorter, J.; Lindquist, S. Prions as adaptive conduits of memory and inheritance. Nat. Rev. Genet. 2005, 6, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Chien, P.; Weissman, J.S.; DePace, A.H. Emerging principles of conformation-based prion inheritance. Annu. Rev. Biochem. 2004, 73, 617–656. [Google Scholar] [CrossRef]
- Chernoff, Y.O. Amyloidogenic domains, prions and structural inheritance: Rudiments of early life or recent acquisition? Curr. Opin. Chem. Biol. 2004, 8, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Uptain, S.M.; Sawicki, G.J.; Caughey, B.; Lindquist, S. Strains of [PSI+] are distinguished by their efficiencies of prion-mediated conformational conversion. EMBO J. 2001, 20, 6236–6245. [Google Scholar] [CrossRef]
- Speldewinde, S.H.; Grant, C.M. The frequency of yeast [PSI+] prion formation is increased during chronological ageing. Microb. Cell 2017, 4, 127–132. [Google Scholar] [CrossRef]
- Tyedmers, J.; Madariaga, M.L.; Lindquist, S. Prion switching in response to environmental stress. PLoS Biol. 2008, 6, e294. [Google Scholar] [CrossRef]
- Chernova, T.A.; Chernoff, Y.O.; Wilkinson, K.D. Prion-based memory of heat stress in yeast. Prion 2017, 11, 151–161. [Google Scholar] [CrossRef]
- Allwein, B.; Kelly, C.; Kammoonah, S.; Mayor, T.; Cameron, D.M. Prion-dependent proteome remodeling in response to environmental stress is modulated by prion variant and genetic background. Prion 2019, 13, 53–64. [Google Scholar] [CrossRef]
- Halfmann, R.; Jarosz, D.F.; Jones, S.K.; Chang, A.; Lancaster, A.K.; Lindquist, S. Prions are a common mechanism for phenotypic inheritance in wild yeasts. Nature 2012, 482, 363–368. [Google Scholar] [CrossRef]
- True, H.L.; Lindquist, S.L. A yeast prion provides a mechanism for genetic variation and phenotypic diversity. Nature 2000, 407, 477–483. [Google Scholar] [CrossRef] [PubMed]
- True, H.L.; Bedin, I.; Lindquist, S.L. Epigenetic regulation of translation reveals hidden genetic variation to produce complex traits. Nature 2004, 431, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Dong, J.; Wall, J.; Frederick, K.K. Amyloid fibrils embodying distinctive yeast prion phenotypes exhibit diverse morphologies. FEMS Yeast Res. 2018, 18, foy059. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Electric Field (kV/cm) | Burst Duration | Pulsed Current * (A) | Energy (J) | Energy Density (kJ/L) | Temperature Rise (°C) |
|---|---|---|---|---|---|
| 2 | 1 ms × 5 (1 Hz) | 0.02 | 0.02 | 0.4 | 0 ± 0.1 |
| 4 | 0.04 | 0.09 | 1.8 | 0.3 ± 0.2 | |
| 6 | 0.07 | 0.20 | 4 | 0.5 ± 0.2 | |
| 8 | 0.09 | 0.35 | 7 | 0.8 ± 0.2 | |
| 10 | 0.11 | 0.55 | 11 | 1 ± 0.3 | |
| 12 | 0.13 | 0.79 | 15.8 | 1.2 ± 0.2 | |
| 14 | 0.15 | 1.08 | 21.6 | 1.3 ± 0.6 | |
| 16 | 0.18 | 1.41 | 28.2 | 1.5 ± 0.2 | |
| 18 | 0.20 | 1.78 | 35.6 | 2.1 ± 0.4 | |
| 20 | 0.22 | 2.20 | 44 | 2.3 ± 0.5 | |
| 22 | 0.24 | 2.66 | 53.2 | 2.8 ± 0.4 | |
| 24 | 0.26 | 3.16 | 63.2 | 3.6 ± 0.9 | |
| 26 | 0.29 | 3.71 | 74.2 | 4.6 ± 1.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jurgelevičiūtė, J.; Bičkovas, N.; Sakalauskas, A.; Novickij, V.; Smirnovas, V.; Lastauskienė, E. Effects of Pulsed Electric Fields on Yeast with Prions and the Structure of Amyloid Fibrils. Appl. Sci. 2021, 11, 2684. https://doi.org/10.3390/app11062684
Jurgelevičiūtė J, Bičkovas N, Sakalauskas A, Novickij V, Smirnovas V, Lastauskienė E. Effects of Pulsed Electric Fields on Yeast with Prions and the Structure of Amyloid Fibrils. Applied Sciences. 2021; 11(6):2684. https://doi.org/10.3390/app11062684
Chicago/Turabian StyleJurgelevičiūtė, Justina, Nedas Bičkovas, Andrius Sakalauskas, Vitalij Novickij, Vytautas Smirnovas, and Eglė Lastauskienė. 2021. "Effects of Pulsed Electric Fields on Yeast with Prions and the Structure of Amyloid Fibrils" Applied Sciences 11, no. 6: 2684. https://doi.org/10.3390/app11062684
APA StyleJurgelevičiūtė, J., Bičkovas, N., Sakalauskas, A., Novickij, V., Smirnovas, V., & Lastauskienė, E. (2021). Effects of Pulsed Electric Fields on Yeast with Prions and the Structure of Amyloid Fibrils. Applied Sciences, 11(6), 2684. https://doi.org/10.3390/app11062684