Global Repigmentation Strategy of Grey Hair Follicles by Targeting Oxidative Stress and Stem Cells Protection

 , and
, and 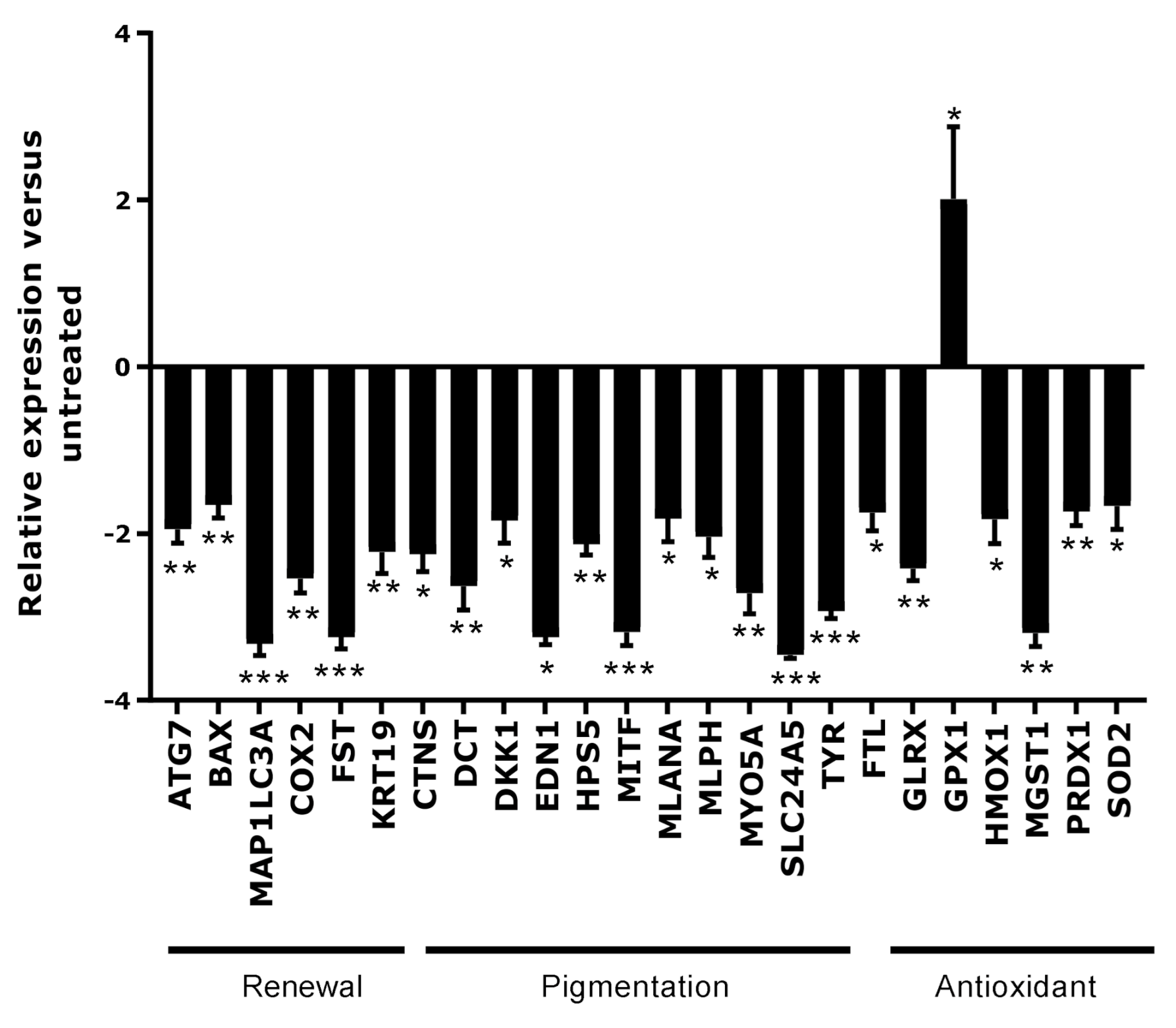{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Hair Pigmentation
1.2. Canities
2. Materials and Methods
2.1. Description and Preparation of the Active Ingredient
2.2. Evaluation of the Impact of Oxidative Stress on Microdissected Hair Follicles
2.2.1. Culture and Treatment of Hair Follicles
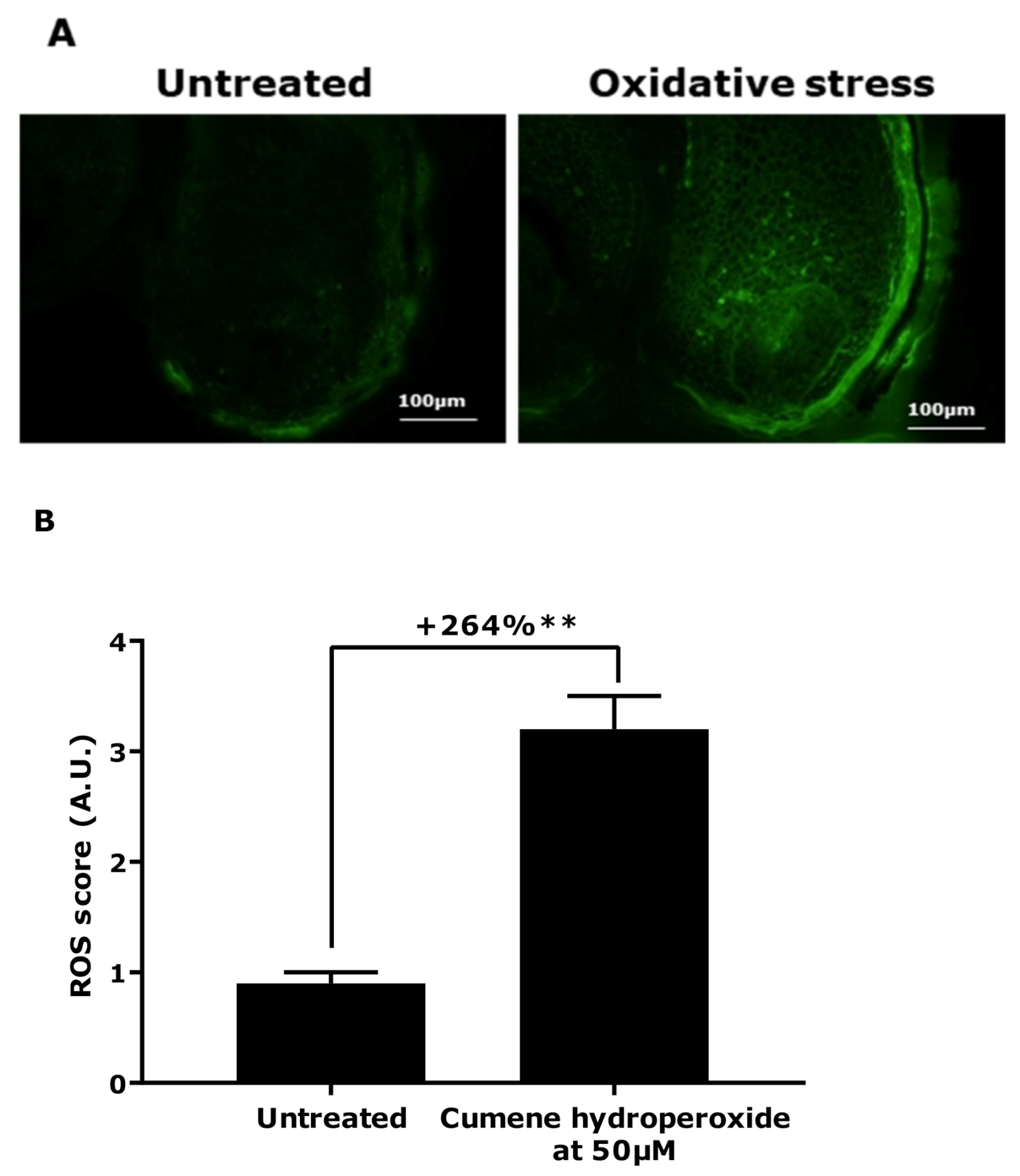
2.2.2. ROS Production Quantification Using DCFH-DA Probe
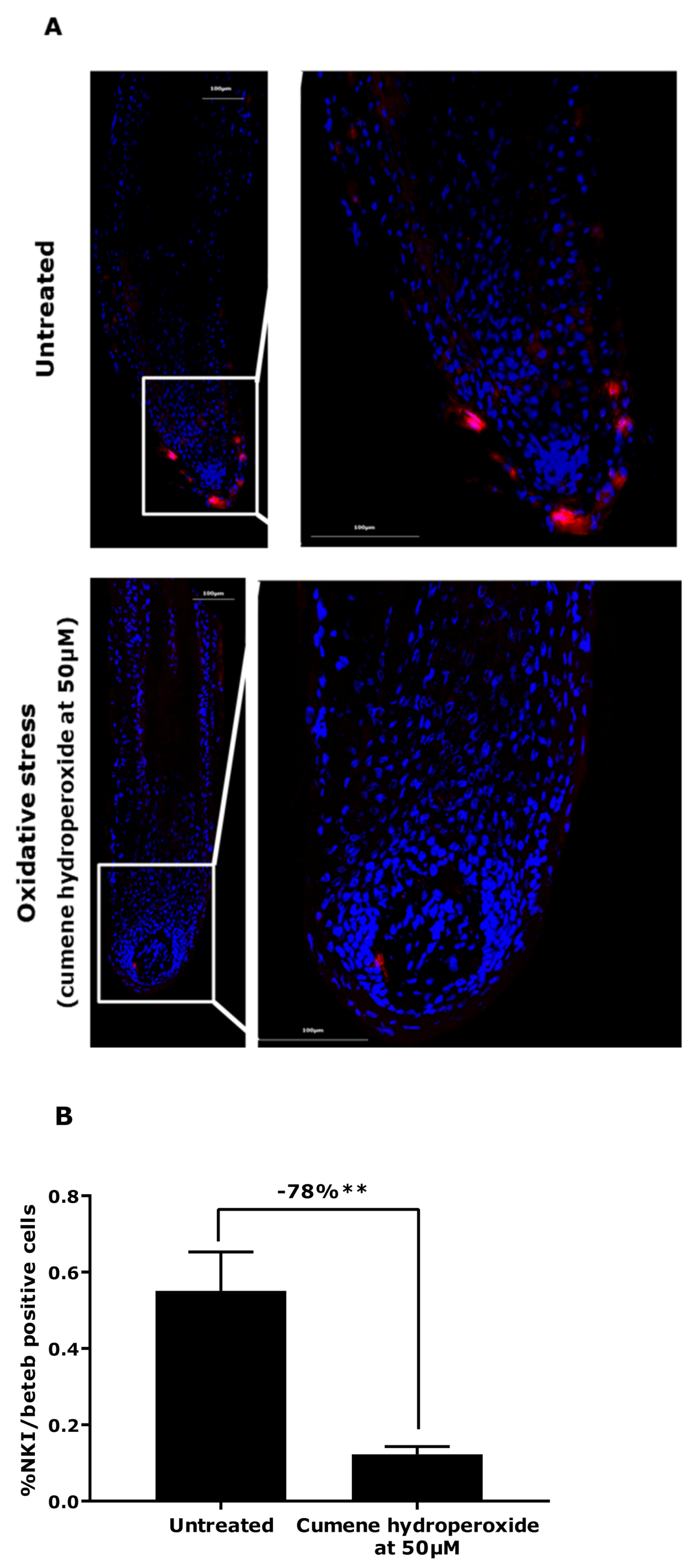
2.2.3. NKI/Beteb Immunostaining
2.3. Evaluation of Melanin Production in Ex Vivo Grey Hair Follicles
2.4. Transcriptomic Analysis in Full Ex Vivo Scalp Skin
2.5. Clinical Investigation
2.5.1. INCI Lotion Formula
2.5.2. Panel Description
2.5.3. Proportion of Grey Hair Assessment with the Scoring Method
2.5.4. Quantification of the Density of Grey Hair
2.6. Statistical Analysis
3. Results
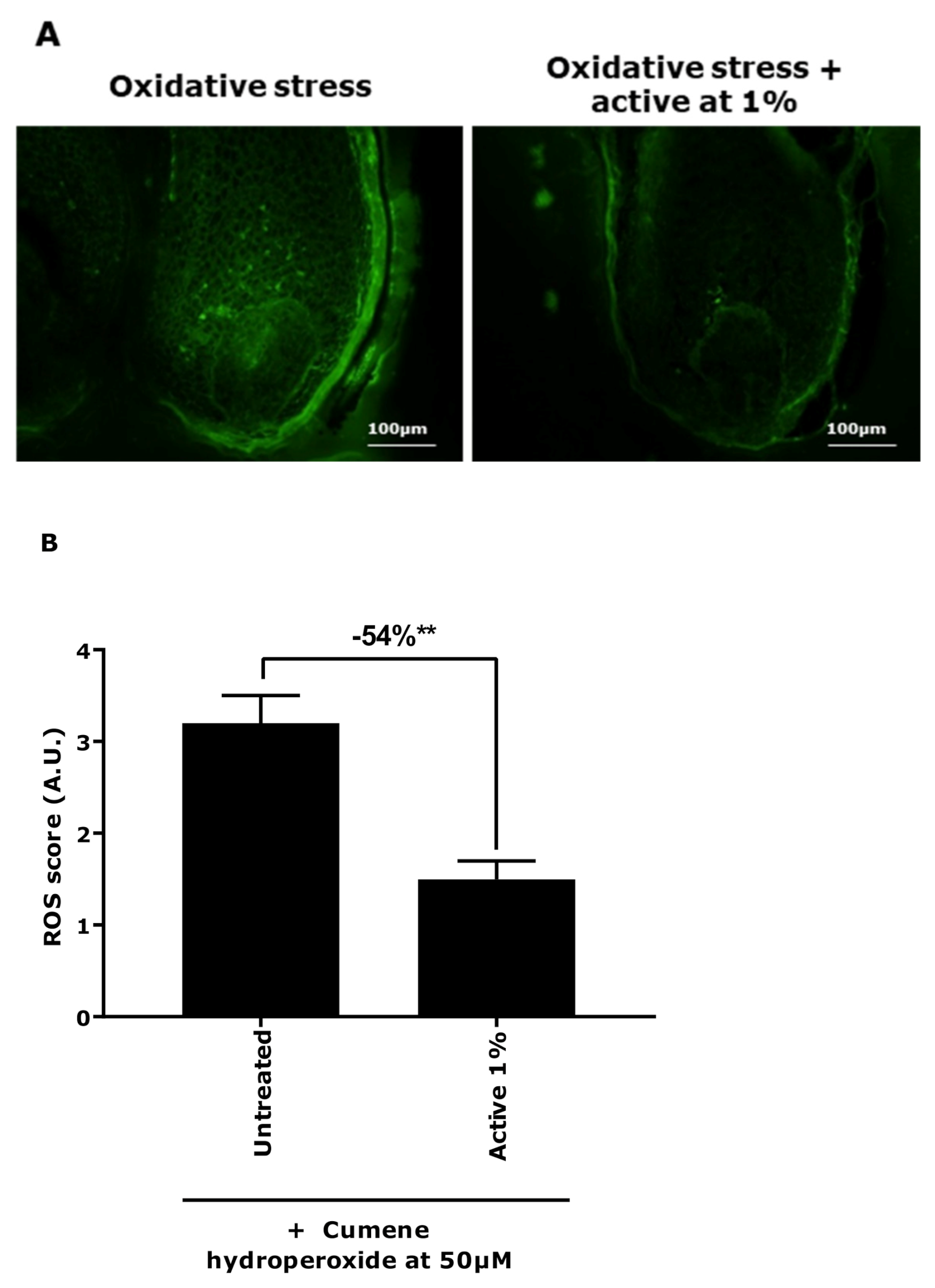
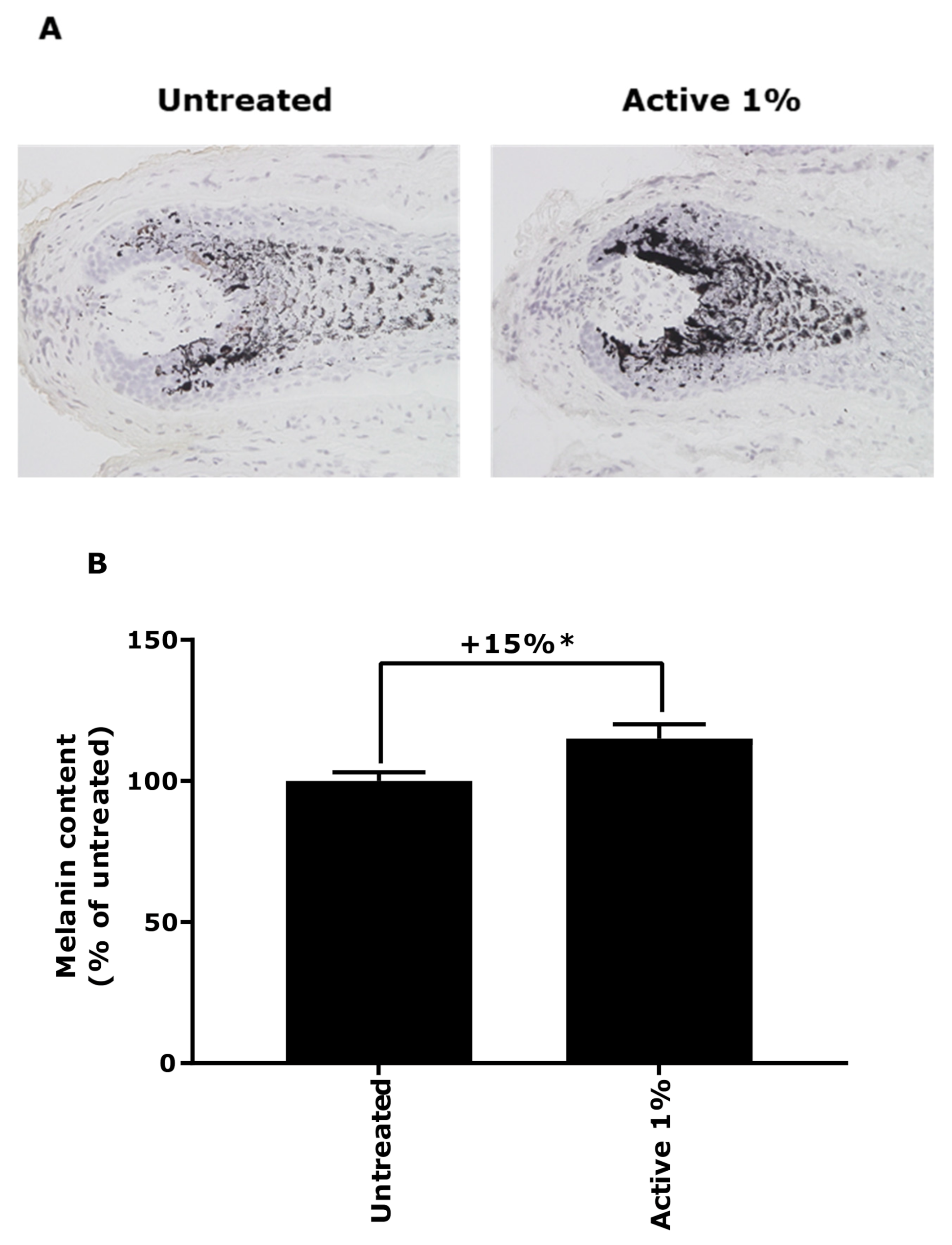
3.1. Demonstration of Oxidative Stress Effect on Ex Vivo Models
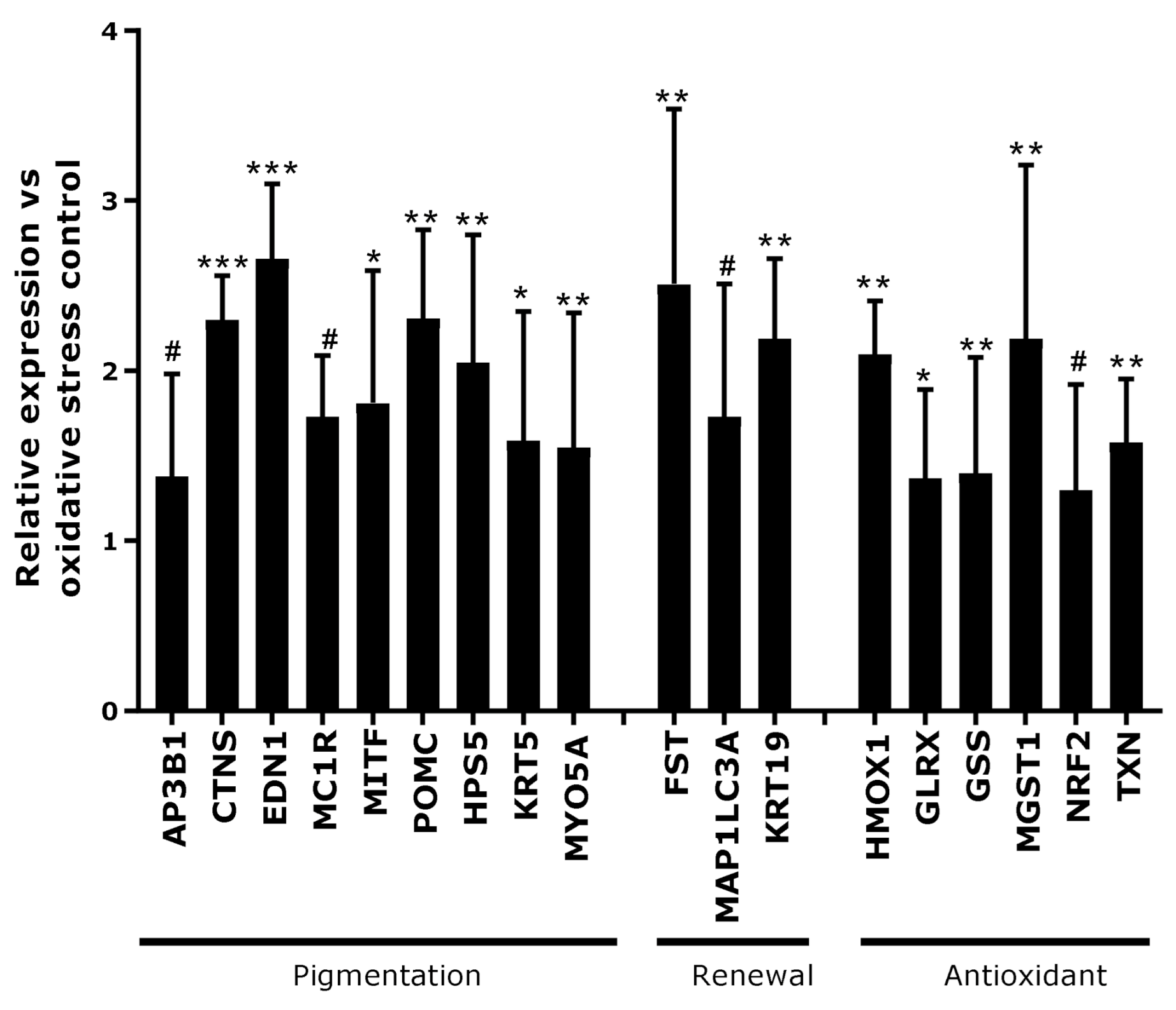
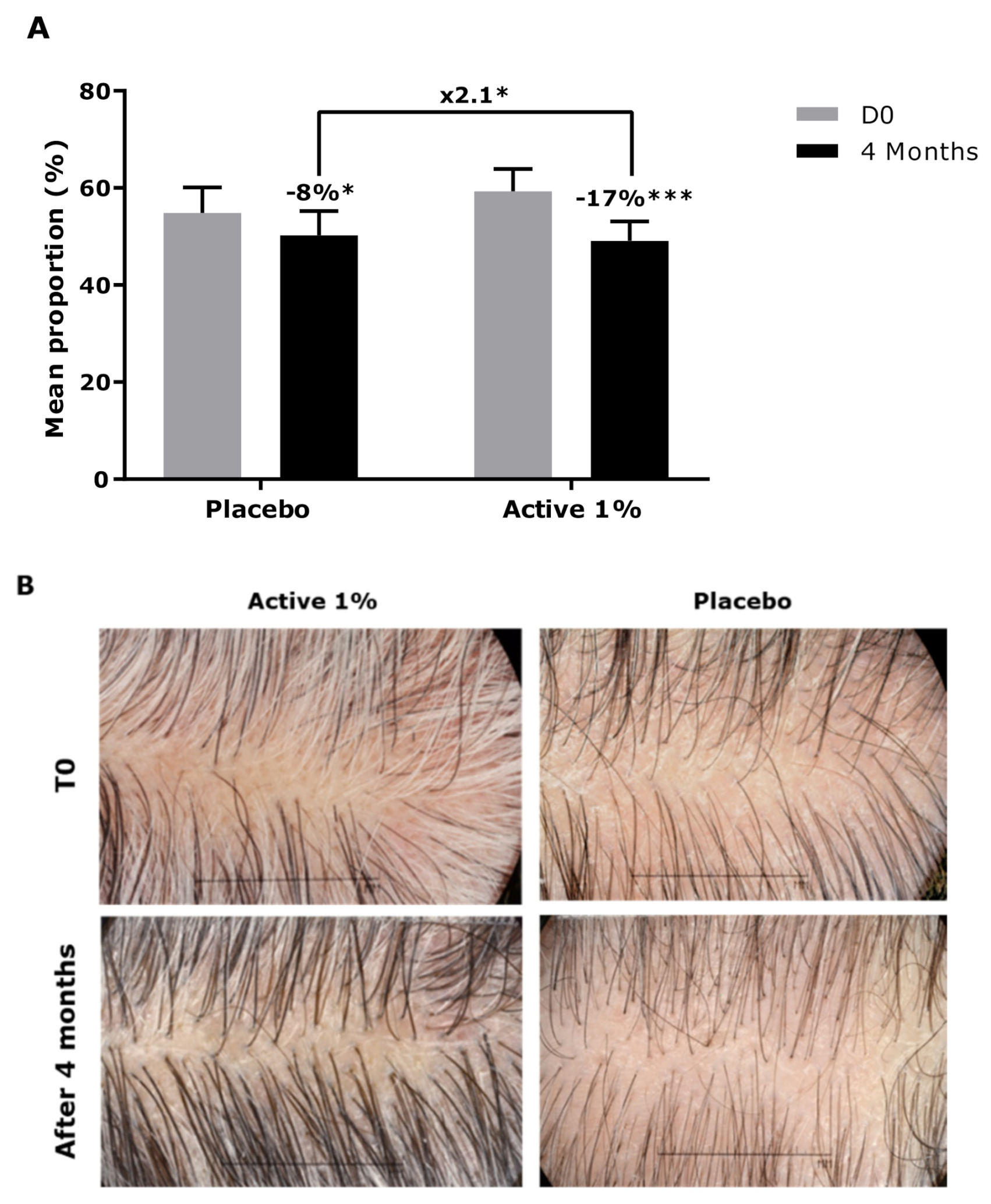
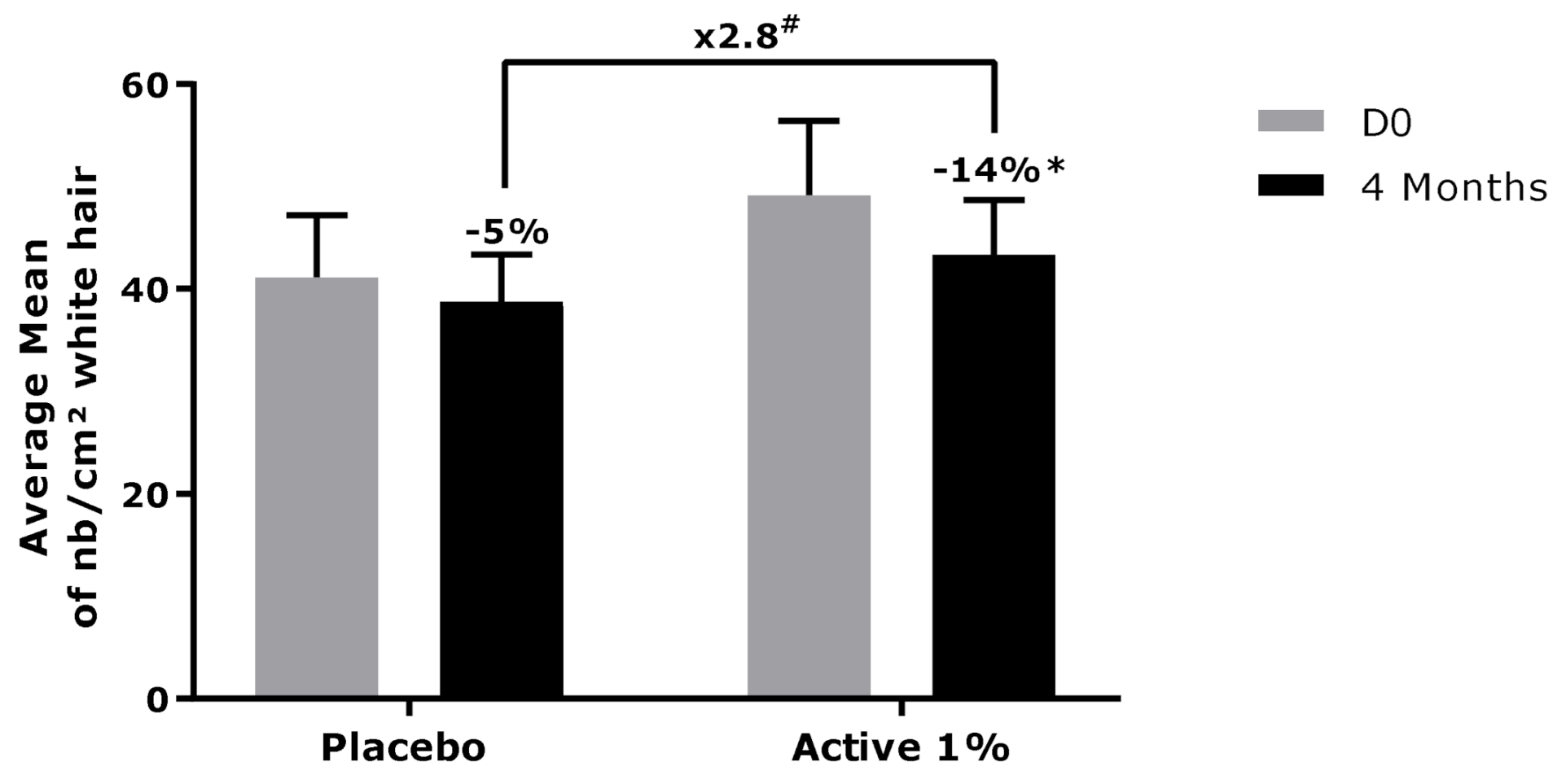
3.2. Effect of Active Ingredient on Canities
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tobin, D.J. The cell biology of human hair follicle pigmentation. Pigment Cell Melanoma Res. 2010, 24, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.J. Human hair pigmentation biological aspects. Int. J. Cosmet. Sci. 2008, 30, 233–257. [Google Scholar] [CrossRef]
- Qiu, W.; Chuong, C.-M.; Lei, M. Regulation of melanocyte stem cells in the pigmentation of skin and its appendages: Biological patterning and therapeutic potentials. Exp. Dermatol. 2019, 28, 395–405. [Google Scholar] [CrossRef]
- Tobin, D.; Paus, R. Graying: Gerontobiology of the hair follicle pigmentary unit. Exp. Gerontol. 2001, 36, 29–54. [Google Scholar] [CrossRef]
- Slominski, A.; Paus, R. Melanogenesis Is Coupled to Murine Anagen: Towar B New Concepts for the Role of Melanocytes and the Regulation of Melanogenesis in Hair Growth. J. Investig. Dermatol. 1993, 101, 8. [Google Scholar] [CrossRef]
- Li, H.; Hou, L. Regulation of melanocyte stem cell behavior by the niche microenvironment. Pigment Cell Melanoma Res. 2018, 31, 556–569. [Google Scholar] [CrossRef]
- Commo, S.; Gaillard, O.; Bernard, B.A. Human hair greying is linked to a specific depletion of hair follicle melanocytes affecting both the bulb and the outer root sheath. Br. J. Dermatol. 2004, 150, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Singal, A.; Daulatabad, D.; Grover, C.; Sharma, S.B.; Chhillar, N. Assessment of oxidative stress in patients with premature canities. Int. J. Trichology 2015, 7, 91. [Google Scholar] [CrossRef]
- Pandhi, D.; Khanna, D. Premature graying of hair. Indian J. Dermatol. Venereol. Leprol. 2013, 79, 641. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.M.; Decker, H.; Hartmann, H.; Chavan, B.; Rokos, H.; Spencer, J.D.; Hasse, S.; Thornton, M.J.; Shalbaf, M.; Paus, R.; et al. Senile hair graying: H2O2-mediated oxidative stress affects human hair color by blunting methionine sulfoxide repair. FASEB J. 2009, 23, 2065–2075. [Google Scholar] [CrossRef]
- Arck, P.C.; Overall, R.; Spatz, K.; Liezman, C.; Handjiski, B.; Klapp, B.F.; Birch-Machin, M.A.; Peters, E.M.J. Towards a “free radical theory of graying”: Melanocyte apoptosis in the aging human hair follicle is an indicator of oxidative stress induced tissue damage. FASEB J. 2006, 20, 1567–1569. [Google Scholar] [CrossRef] [PubMed]
- Trüeb, R.M. The impact of oxidative stress on hair. Int. J. Cosmet. Sci. 2015, 37, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Luo, L.-F.; Liu, X.-M.; Zhou, Q.; Xu, S.-Z.; Lei, T.-C. Premature Graying as a Consequence of Compromised Antioxidant Activity in Hair Bulb Melanocytes and Their Precursors. PLoS ONE 2014, 9, e93589. [Google Scholar] [CrossRef]
- Tang, L.; Li, J.; Lin, X.; Wu, W.; Kang, K.; Fu, W. Oxidation Levels Differentially Impact Melanocytes: Low versus High Concentration of Hydrogen Peroxide Promotes Melanin Synthesis and Melanosome Transfer. Dermatology 2012, 224, 145–153. [Google Scholar] [CrossRef]
- Jiménez-Cervantes, C.; Martinez-Esparza, M.; Pérez, C.; Daum, N.; Solano, F.; Garcia-Borron, J.C. Melanogenesis inhibition by oxidative stress: Transient downregulation of melanocyte differentiation markers and possible involvement of microphthalmia transcription factor. J. Cell Sci. 2001, 114, 2335–2344. [Google Scholar]
- Paul, F.; Auriol, D.; Oriol, E.; Monsan, P. Production and Purification of Dextransucrase from Leuconostoc mesenteroides, NRRL B 512 (F). Ann. N. Y. Acad. Sci. 1984, 434, 267–270. [Google Scholar] [CrossRef]
- Langan, E.A.; Philpott, M.P.; Kloepper, J.E.; Paus, R. Human hair follicle organ culture: Theory, application and perspectives. Exp. Dermatol. 2015, 24, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Singal, A.; Daulatabad, D.; Grover, C. Graying severity score: A useful tool for evaluation of premature canities. Indian Dermatol. Online J. 2016, 7, 164. [Google Scholar] [CrossRef]
- Triwongwaranat, D.; Thuangtong, R.; Arunkajohnsak, S. A review of the etiologies, clinical characteristics, and treatment of canities. Int. J. Dermatol. 2019, 58, 659–666. [Google Scholar] [CrossRef]
- Krutmann, J.; Bouloc, A.; Sore, G.; Bernard, B.A.; Passeron, T. The skin aging exposome. J. Dermatol. Sci. 2017, 85, 152–161. [Google Scholar] [CrossRef]
- O’Sullivan, J.D.B.; Nicu, C.; Picard, M.; Chéret, J.; Bedogni, B.; Tobin, D.J.; Paus, R. The biology of human hair greying. Biol. Rev. 2021, 96, 107–128. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Z.; Wang, X.; Xiang, L.; Zhang, C. Dysfunction of Autophagy: A Possible Mechanism Involved in the Pathogenesis of Vitiligo by Breaking the Redox Balance of Melanocytes. Oxidative Med. Cell. Longev. 2016, 2016, 1–7. [Google Scholar] [CrossRef]
- Setaluri, V. Autophagy as a Melanocytic Self-Defense Mechanism. J. Investig. Dermatol. 2015, 135, 1215–1217. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-F.; Gruber, F.; Ni, C.; Mildner, M.; Koenig, U.; Karner, S.; Barresi, C.; Rossiter, H.; Narzt, M.-S.; Nagelreiter, I.M.; et al. Suppression of Autophagy Dysregulates the Antioxidant Response and Causes Premature Senescence of Melanocytes. J. Investig. Dermatol. 2015, 135, 1348–1357. [Google Scholar] [CrossRef]
- Samuelov, L.; Sprecher, E.; Sugawara, K.; Singh, S.K.; Tobin, D.J.; Tsuruta, D.; Bíró, T.; Kloepper, J.E.; Paus, R. Topobiology of Human Pigmentation: P-Cadherin Selectively Stimulates Hair Follicle Melanogenesis. J. Investig. Dermatol. 2013, 133, 1591–1600. [Google Scholar] [CrossRef]
- Adhikari, K.; Fontanil, T.; Cal, S.; Mendoza-Revilla, J.; Fuentes-Guajardo, M.; Chacón-Duque, J.-C.; Al-Saadi, F.; Johansson, J.A.; Quinto-Sanchez, M.; Acuña-Alonzo, V.; et al. A genome-wide association scan in admixed Latin Americans identifies loci influencing facial and scalp hair features. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef]
- Meunier, M.; Scandolera, A.; Chapuis, E.; Lambert, C.; Jarrin, C.; Robe, P.; Chajra, H.; Auriol, D.; Reynaud, R. From stem cells protection to skin microbiota balance: Orobanche rapum extract, a new natural strategy. J. Cosmet. Dermatol. 2019, 18, 1140–1154. [Google Scholar] [CrossRef]
- Liao, N.; Shi, Y.; Zhang, C.; Zheng, Y.; Wang, Y.; Zhao, B.; Zeng, Y.; Liu, X.; Liu, J. Antioxidants inhibit cell senescence and preserve stemness of adipose tissue-derived stem cells by reducing ROS generation during long-term in vitro expansion. Stem Cell Res. Ther. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Xican, L.; Xie, H.; Jiang, Q.; Wei, G.; Lin, L.; Li, C.; Ou, X.; Yang, L.; Xie, Y.; Fu, Z.; et al. The mechanism of (+) taxifolin’s protective antioxidant effect for •OH-treated bone marrow-derived mesenchymal stem cells. Cell. Mol. Biol. Lett. 2017, 22. [Google Scholar] [CrossRef]
- Petruk, G.; Del Giudice, R.; Rigano, M.M.; Monti, D.M. Antioxidants from Plants Protect against Skin Photoaging. Oxidative Med. Cell. Longev. 2018, 2018, 1–11. [Google Scholar] [CrossRef]
- Ma, G.-W.; Chu, Y.-K.; Zhang, W.-J.; Qin, F.-Y.; Xu, S.-S.; Yang, H.; Rong, E.-G.; Du, Z.-Q.; Wang, S.-Z.; Li, H.; et al. Polymorphisms of FST gene and their association with wool quality traits in Chinese Meri sheep. PLoS ONE 2017, 12, e0174868. [Google Scholar] [CrossRef]
- Chandrasekaran, N.C.; Sanchez, W.Y.; Mohammed, Y.H.; Grice, J.E.; Roberts, M.S.; Barnard, R.T. Permeation of topically applied Magnesium ions through human skin is facilitated by hair follicles. Magnes. Res. 2016, 29, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Illel, B.; Schaefer, H.; Wepierre, J.; Doucet, O. Follicles Play an Important Role in Percutaneous Absorption. J. Pharm. Sci. 1991, 80, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Otberg, N.; Patzelt, A.; Rasulev, U.; Hagemeister, T.; Linscheid, M.; Sinkgraven, R.; Sterry, W.; Lademann, J. The role of hair follicles in the percutaneous absorption of caffeine. Br. J. Clin. Pharmacol. 2008, 65, 488–492. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Tollenaere, M.; Chapuis, E.; Auriol, P.; Auriol, D.; Scandolera, A.; Reynaud, R. Global Repigmentation Strategy of Grey Hair Follicles by Targeting Oxidative Stress and Stem Cells Protection. Appl. Sci. 2021, 11, 1533. https://doi.org/10.3390/app11041533
De Tollenaere M, Chapuis E, Auriol P, Auriol D, Scandolera A, Reynaud R. Global Repigmentation Strategy of Grey Hair Follicles by Targeting Oxidative Stress and Stem Cells Protection. Applied Sciences. 2021; 11(4):1533. https://doi.org/10.3390/app11041533
Chicago/Turabian StyleDe Tollenaere, Morgane, Emilie Chapuis, Pascale Auriol, Daniel Auriol, Amandine Scandolera, and Romain Reynaud. 2021. "Global Repigmentation Strategy of Grey Hair Follicles by Targeting Oxidative Stress and Stem Cells Protection" Applied Sciences 11, no. 4: 1533. https://doi.org/10.3390/app11041533
APA StyleDe Tollenaere, M., Chapuis, E., Auriol, P., Auriol, D., Scandolera, A., & Reynaud, R. (2021). Global Repigmentation Strategy of Grey Hair Follicles by Targeting Oxidative Stress and Stem Cells Protection. Applied Sciences, 11(4), 1533. https://doi.org/10.3390/app11041533