
Structure of Microbial Communities When Complementary Effluents Are Anaerobically Digested


Abstract
1. Introduction
2. Materials and Methods
2.1. Substrates Sampling
2.2. Anaerobic Digestion Experimental Set-Up
2.3. Analytical and Chromatograph Methods
2.4. Molecular Analysis
2.4.1. DNA Extraction and MiSeq Sequencing (Next-Generation Sequencing, NGS) of 16S rRNA Gene Amplicons
2.4.2. Data Analysis
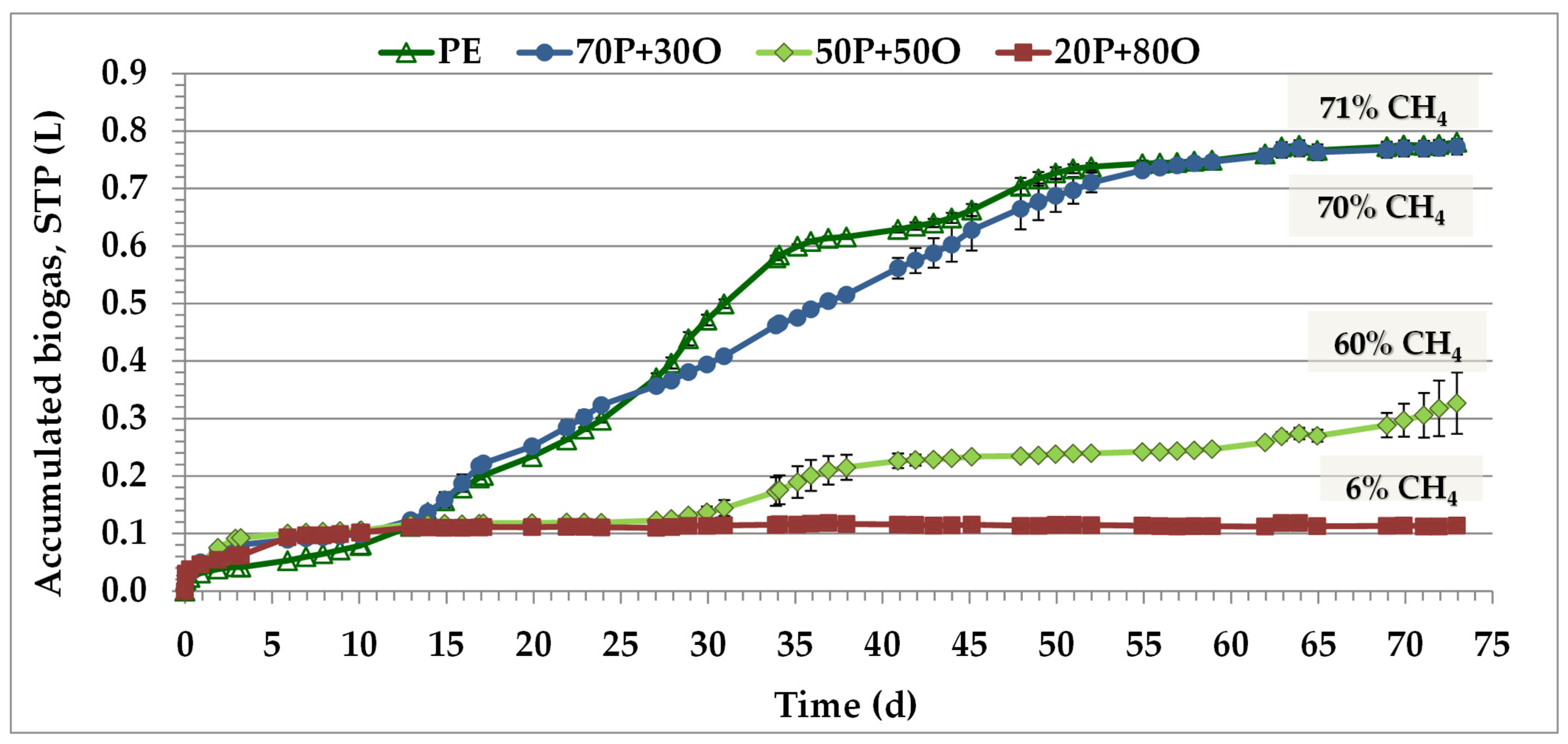
3. Results and Discussion
3.1. Chemical Composition of Effluents
3.2. Anaerobic Digestion of PE and OMW Mixture
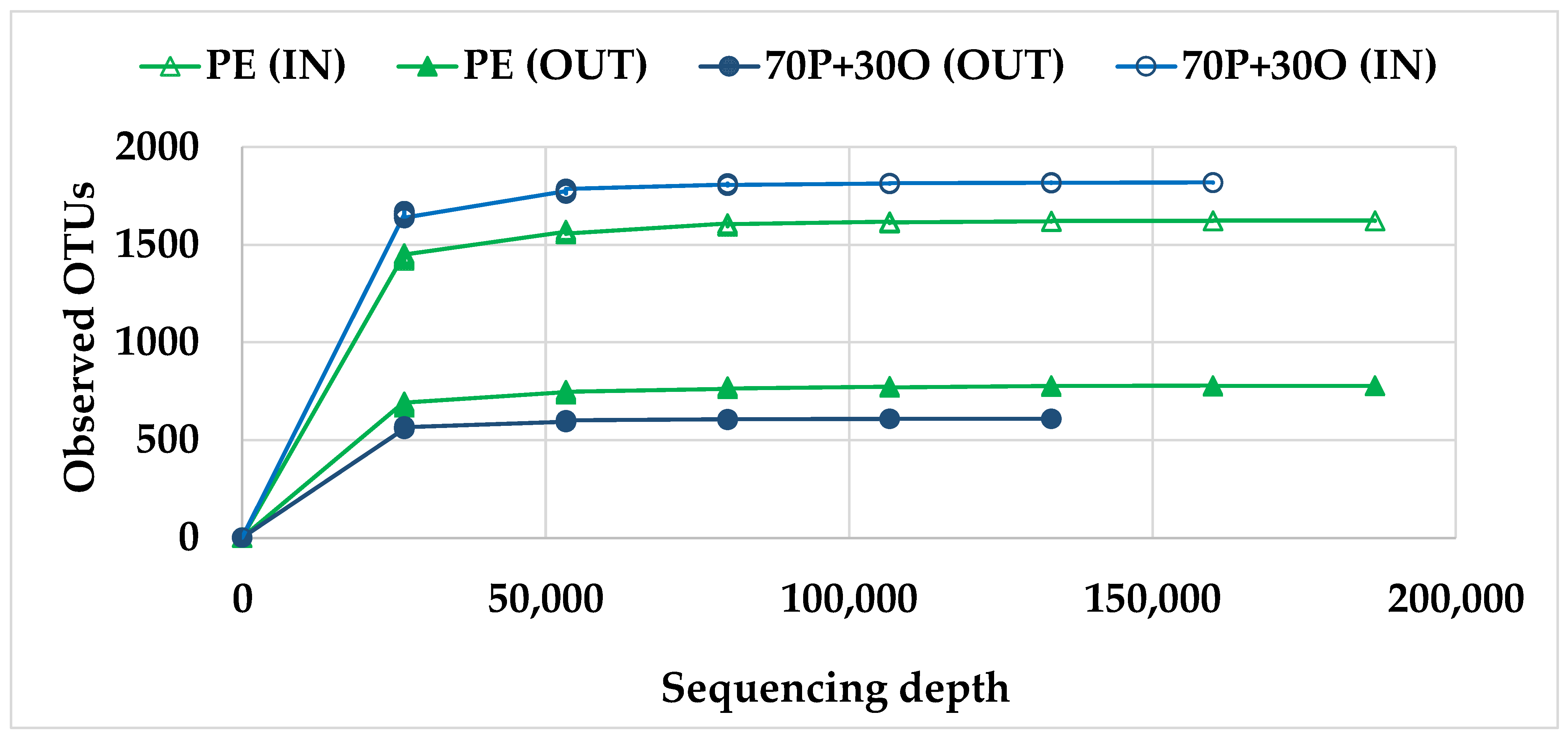
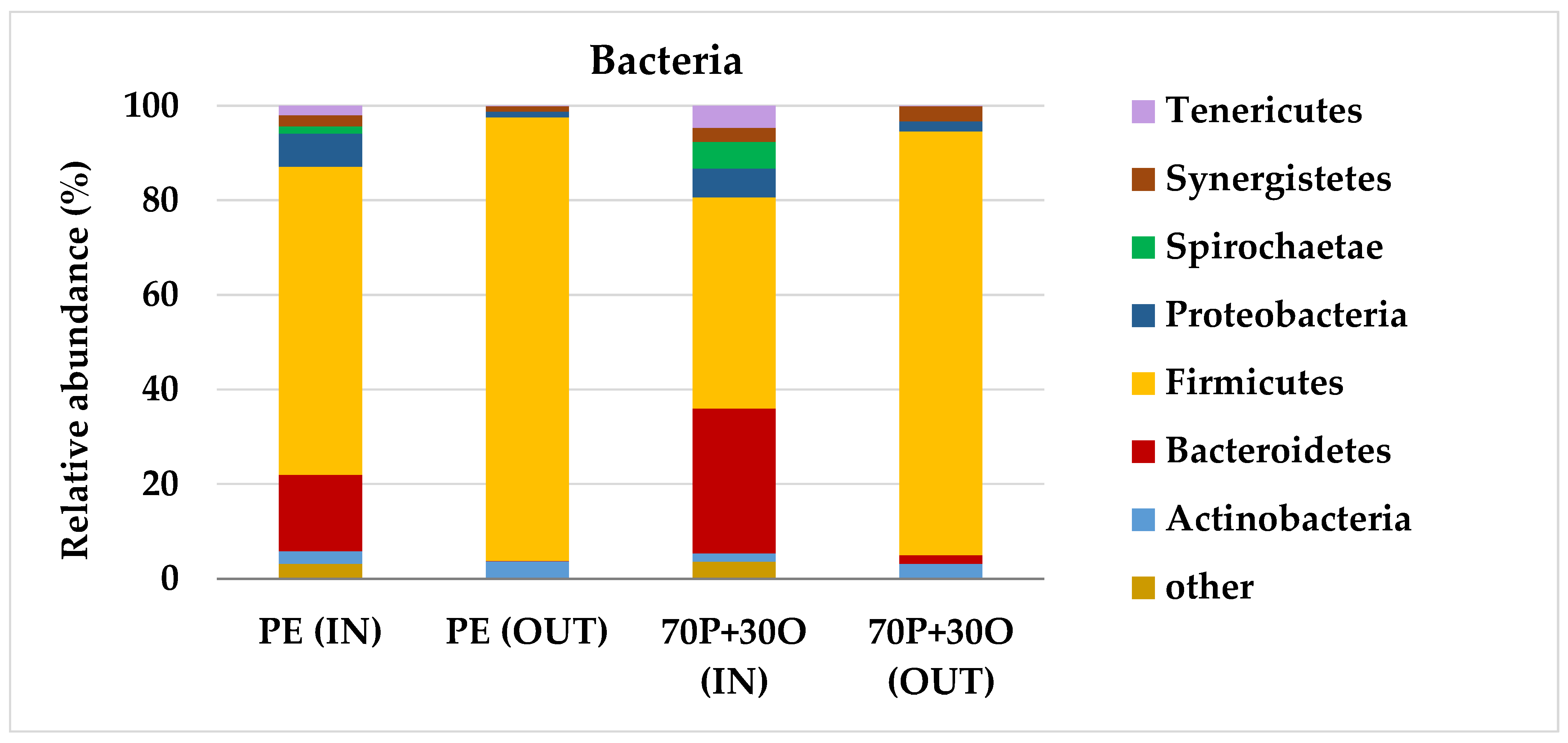
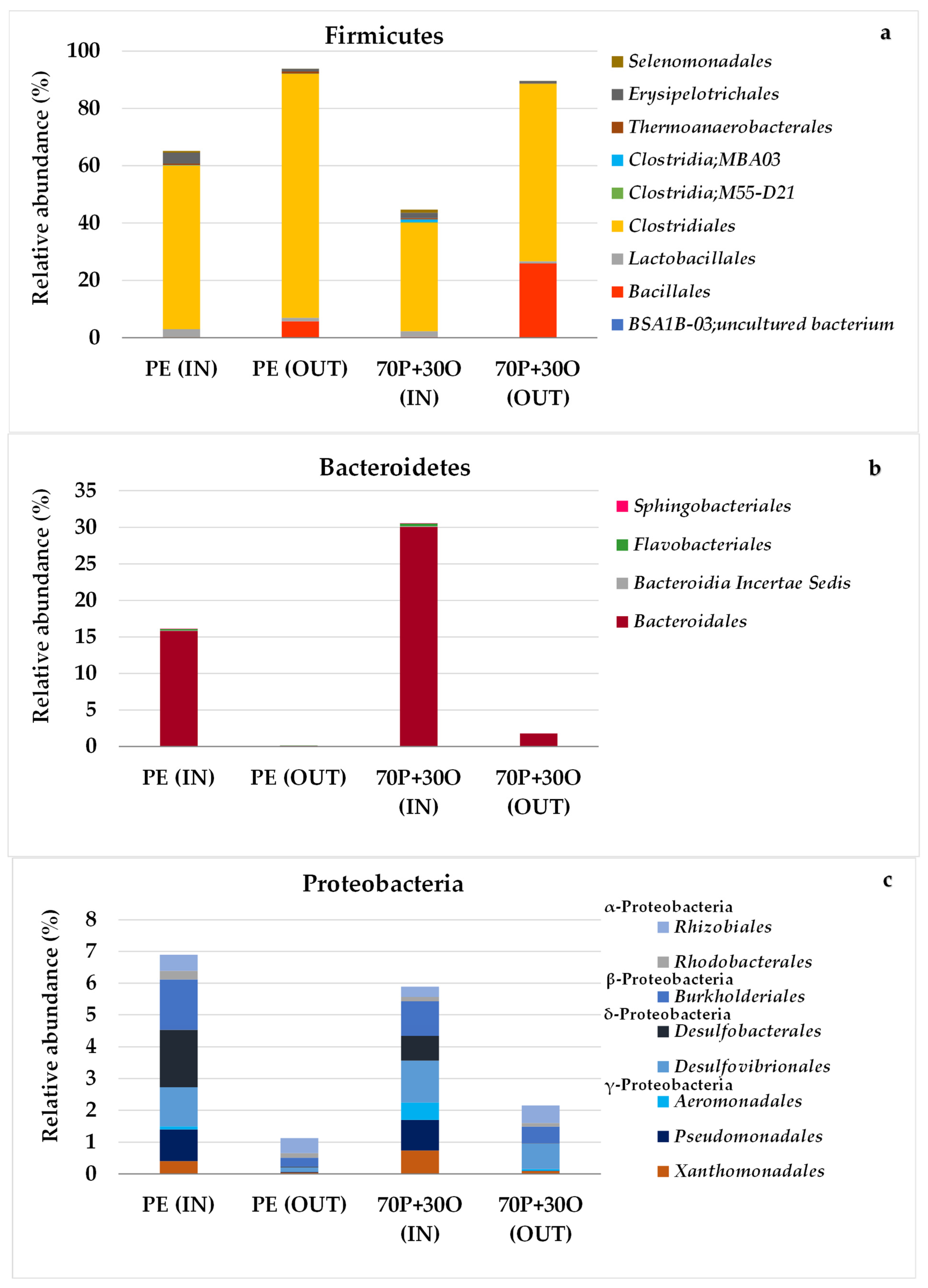
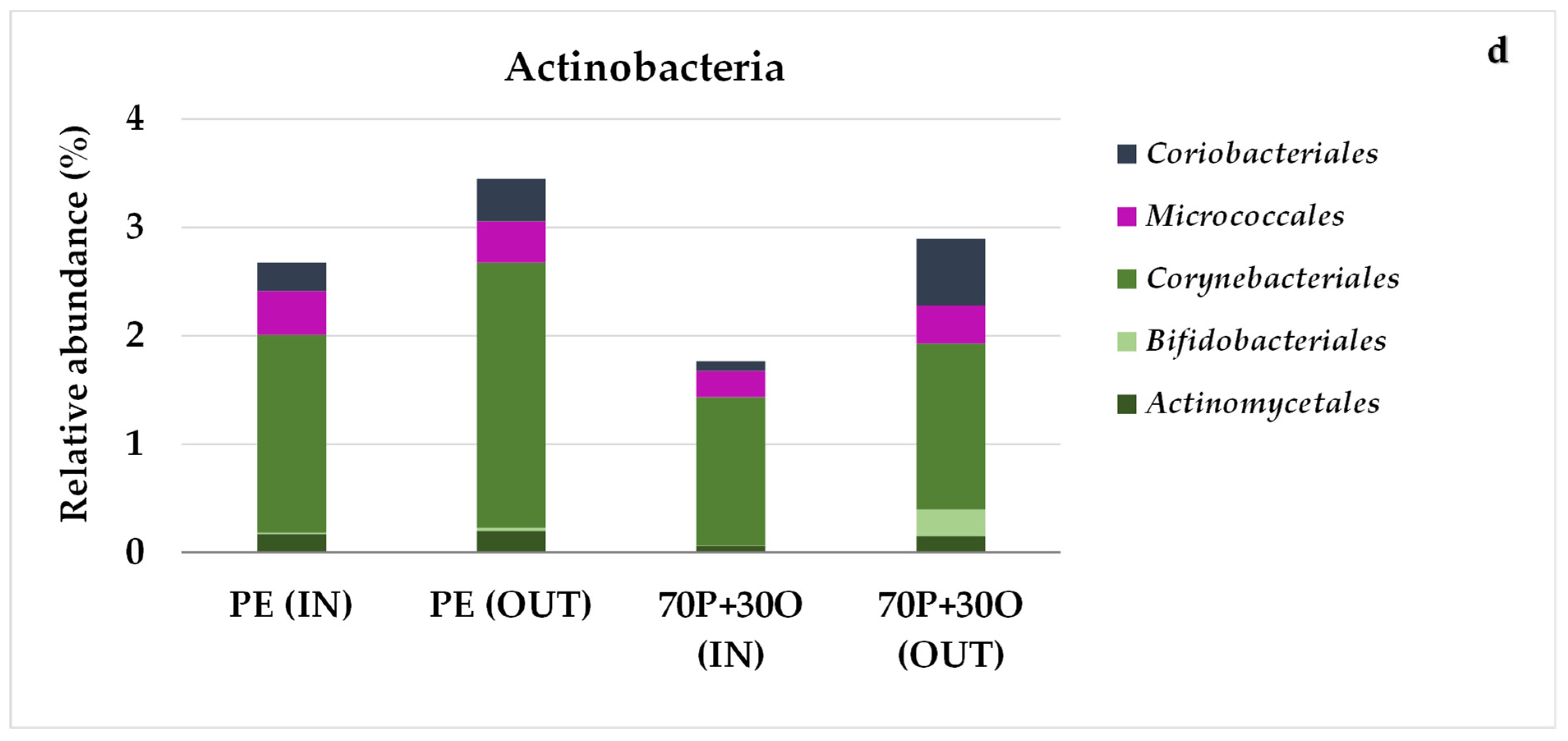
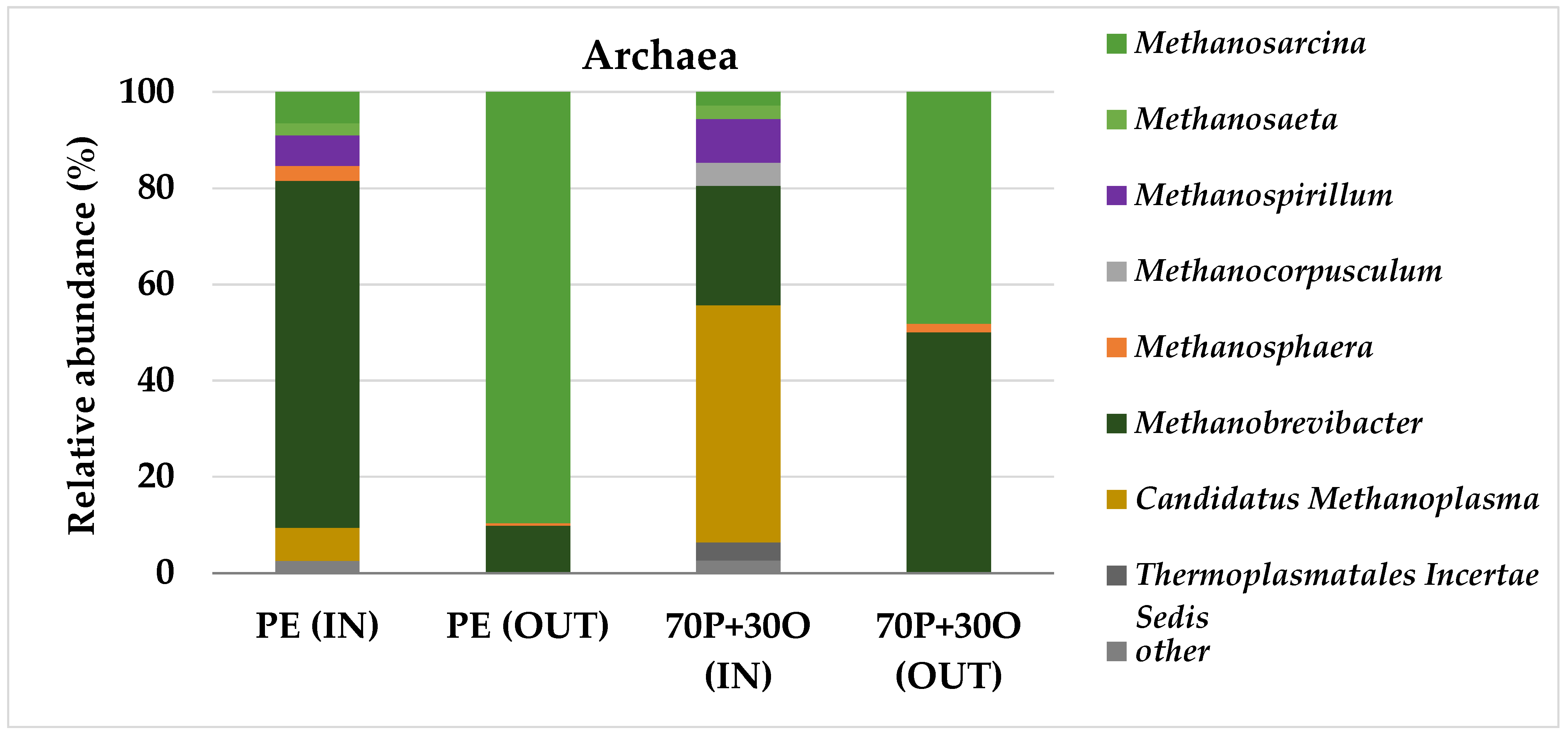
3.3. Molecular Characterization of Microbial Communities
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Q.; He, J.; Tian, M.; Mao, Z.; Tang, L.; Zhang, J.; Zhang, H. Enhancement of methane production from cassava residues by biological pretreatment using a constructed microbial consortium. Bioresour. Technol. 2011, 102, 8899–8906. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. 2019. Available online: http://www.fao.org/faostat/en/#data/QA (accessed on 15 February 2019).
- Dermeche, S.; Nadour, M.; Larroche, C.; Moulti-Mati, F.; Michaud, P. Olive mill wastes: Biochemical characterizations and valorization strategies. Process. Biochem. 2013, 48, 1532–1552. [Google Scholar] [CrossRef]
- El Mekawy, A.; Diels, L.; Bertin, L.; De Wever, H.; Pant, D. Potential biovalorization techniques for olive mill biorefinery wastewater. Biofuels Bioprod. Bioref. 2014, 8, 283–293. [Google Scholar] [CrossRef]
- Eroglu, E.; Eroglu, I.; Gündüz, U.; Türker, L.; Yücel, M. Biological hydrogen production from olive mill wastewater with two-stage processes. Int. J. Hydrogen Energy 2006, 31, 1527–1535. [Google Scholar] [CrossRef]
- Paraskeva, P.; Diamadopoulos, E. Technologies for olive mill wastewater (OMW) treatment: A review. J. Chem. Technol. Biotechnol. 2006, 81, 1475–1485. [Google Scholar] [CrossRef]
- Eusébio, A.; Tacão, M.; Chaves, S.; Tenreiro, R.; Almeida-Vara, E. Molecular assessment of microbiota structure and dynamics along mixed olive oil and winery wastewaters biotreatment. Biodegradation 2011, 22, 773–795. [Google Scholar] [CrossRef] [PubMed]
- Tsagaraki, E.; Lazarides, H.N.; Petrotos, K.B. Olive Mill Wastewater Treatment. In Utilization of By-Products and Treatment of Waste in the Food Industry; Oreopoulou, V., Russ, W., Eds.; Springer: New York, NY, USA, 2007; Volume 8, pp. 133–157. [Google Scholar] [CrossRef]
- Kapellakis, I.E.; Tsagarakis, K.P.; Crowther, J.C. Olive oil history, production and by-product management. Rev. Environ. Sci. Biotechnol. 2008, 7, 1–26. [Google Scholar] [CrossRef]
- McNamara, C.J.; Anastasiou, C.C.; O’Flaherty, V.; Mitchell, R. Bioremediation of olive mill wastewater. Int. Biodeterior. Biodegradation. 2008, 61, 127–134. [Google Scholar] [CrossRef]
- Rahmanian, N.; Jafari, S.M.; Galamakis, C.M. Recovery and Removal of Phenolic Compounds of Olive Mill Wastewater. J. Am. Oil Chem. Soc. 2014, 91, 1–18. [Google Scholar] [CrossRef]
- Bernet, N.; Béline, F. Challenges and innovations on biological treatment of livestock effluents. Bioresour. Technol. 2009, 100, 5431–5436. [Google Scholar] [CrossRef]
- Prapaspongsa, T.; Poulsen, T.G.; Hansen, J.A.; Christensen, P. Energy production, nutrient recovery and greenhouse gas emission potentials from integrated pig manure management systems. Waste Manag. Res. 2010, 28, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Boopathy, R. Biological treatment of swine waste using anaerobic baffled reactors. Bioresour. Technol. 1998, 64, 1–6. [Google Scholar] [CrossRef]
- Hwang, K.; Song, M.; Kim, W.; Kim, M.; Hwang, S. Effects of prolonged starvation on methanogenic population dynamics in anaerobic digestion of swine wastewater. Bioresour. Technol. 2010, 101, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Venkiteshwaran, K.; Bocher, B.; Maki, J.; Zitomer, D. Relating Anaerobic Digestion Microbial Community and Process Function. Microbiol. Insights 2015, 8, 37–44. [Google Scholar] [CrossRef]
- Cai, L.; Li, H.; Yao, H. A Membership-Fusing Model for Characterizing the Shift of Methanogen Community in a Three-Stage Sludge-Treatment Process. Appl. Sci. 2020, 10, 4274. [Google Scholar] [CrossRef]
- Sulbarán-Rangel, B.; Aguirre, J.S.A.; Breton-Deval, L.; del Real-Olvera, J.; Tun, K.J.G. Improvement of Anaerobic Digestion of Hydrolysed Corncob Waste by Organosolv Pretreatment for Biogas Production. Appl. Sci. 2020, 10, 2785. [Google Scholar] [CrossRef]
- Gelegenis, J.; Georgakakis, D.; Angelidaki, I.; Christopoulou, N.; Goumenaki, M. Optimization of biogas production from olive oil mill wastewater, by codigesting with diluted poultry manure. Appl. Energy 2007, 84, 646–663. [Google Scholar] [CrossRef]
- Dareioti, M.A.; Dokianakis, S.N.; Stamatelatou, K.; Zafiri, C.; Kornaros, M. Biogas production from anaerobic co-digestion of agroindustrial wastewaters under mesophilic conditions in a two-stage process. Desalination 2009, 248, 891–906. [Google Scholar] [CrossRef]
- Martinez-Garcia, G.; Johnson, A.C.; Bachmann, R.T.; Williams, C.J.; Burgoyne, A.; Edyvean, R.G.J. Anaerobic treatment of olive mill wastewater and piggery effluents fermented with Candida tropicalis. J. Hazard. Mater. 2009, 164, 1398–1405. [Google Scholar] [CrossRef]
- Marques, I.P.; Texeira, A.; Rodrigues, L.; Dias, S.M.; Novais, J.M. Anaerobic co-treatment of olive mill and piggery effluents. Environ. Technol. 1997, 18, 265–274. [Google Scholar] [CrossRef]
- Marques, I.P.; Texeira, A.; Rodrigues, L.; Dias, S.M.; Novais, J.M. Anaerobic treatment of olive mill wastewater with digested piggery effluent. Water Environ. Res. 1998, 70, 1056–1061. [Google Scholar] [CrossRef]
- Marques, I.P. Anaerobic digestion treatment of olive mill wastewater for effluent re-use in irrigation. Desalination 2001, 137, 233–239. [Google Scholar] [CrossRef]
- Sampaio, M.A.; Gonçalves, M.R.; Marques, I.P. Anaerobic digestion challenge of raw olive mill wastewater. Bioresour. Technol. 2011, 102, 10810–10818. [Google Scholar] [CrossRef] [PubMed]
- Marques, I.P.; Eusébio, A.; La Cara, F. Processo para a produção de biogás e obtenção de biomoléculas num só passo. Portuguese Patent 108918, 30 July 2020. [Google Scholar]
- La Cara, F.; Ionata, E.; Del Monaco, G.; Marcolongo, L.; Gonçalves, M.R.; Marques, I.P. Olive Mill Wastewater Anaerobically Digested: Phenolic Compounds with Antiradical Activity. Chem. Eng. Trans. 2012, 27, 325–330. [Google Scholar] [CrossRef]
- Bailón-Salas, A.M.; Medrano-Roldán, H.; Valle-Cervantes, S.; Ordaz-Díaz, L.A.; Urtiz-Estrada, N.; Rojas-Contreras, J.A. Review of molecular techniques for the identification of bacterial communities in biological effluent treatment facilities at pulp and paper mills. Bioresources 2017, 12, 4384–4409. [Google Scholar] [CrossRef]
- Liu, L.; Li, Y.; Li, S.; Hu, N.; He, Y.; Pong, R.; Lin, D.; Lu, L.; Law, M. Comparison of Next-Generation Sequencing Systems. BioMed Research Int. 2012, 2012, 251364. [Google Scholar] [CrossRef]
- Vacher, C.; Tamaddoni-Nezhad, A.; Kamenova, S.; Peyrard, N.; Moalic, Y.; Sabbadin, R.; Schwaller, L.; Chiquet, J.; Smith, M.A.; Vallance, J.; et al. Learning Ecological Networks from Next-Generation Sequencing Data. In Advances in Ecological Research; Woodward, G., Bohan, D.A., Eds.; Academic Press: Oxford, UK, 2016; Volume 54, pp. 1–39. [Google Scholar]
- APHA; AWWA; EWF. Standard Methods for Examination of Water and Wastewater, 22nd ed.; American Public Health Association; American Water Works Association; Water Environment Federation: Washington, DC, USA, 2012. [Google Scholar]
- Singleton, V.L.; Rossi, J.A., Jr. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Eonol. Viticult. 1965, 16, 144–158. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Roseiro, L.; Tavares, C.S.; Roseiro, J.C.; Rauter, A.P. Antioxidants from aqueous decoction of carob pods biomass (Ceretonia siliqua L.): Optimisation using response surface methodology and phenolic profile by capillary electrophoresis y. Ind. Crop. Prod. 2013, 44, 119–126. [Google Scholar] [CrossRef]
- Zhou, J.; Bruns, M.A.; Tiedje, J.M. DNA Recovery from Soils of Diverse Composition. Appl. Environ. Microbiol. 1996, 62, 316–322. [Google Scholar] [CrossRef]
- Illumina 16S Metagenomic Sequencing Library Preparation. 2013. Available online: https://emea.support.illumina.com/downloads/16s_metagenomic_sequencing_library_preparation.html (accessed on 1 December 2019).
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High resolution sample interference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2 Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Glöckner, F.O.; Yilmaz, P.; Quast, C.; Gerken, J.; Beccati, A.; Ciuprina, A.; Bruns, G.; Yarza, P.; Peplies, J.; Westram, R.; et al. 25 years of serving the community with ribosomal RNA gene reference databases and tools. J. Biotechnol. 2017, 261, 169–176. [Google Scholar] [CrossRef]
- Ergüder, T.H.; Güven, E.; Demirer, G.N. Anaerobic treatment of olive mill wastes in batch reactors. Proc. Biochem. 2000, 36, 243–248. [Google Scholar] [CrossRef]
- Boursier, H.; Béline, F.; Paul, E. Piggery wastewater characterisation for biological nitrogen removal process design. Bioresour. Technol. 2005, 96, 351–358. [Google Scholar] [CrossRef]
- Velho, V.F.; Mohedano, R.A.; Belli Filho, P.; Costa, R.H.R. The viability of treated piggery wastewater for reuse in agricultural irrigation. Int. J. Recycl. Org. Waste Agric. 2012, 1, 10. [Google Scholar] [CrossRef]
- Aboudi, K.; Gómez-Quiroga, X.; Álvarez-Gallego, C.J.; Romero-García, L.I. Insights into Anaerobic Co-Digestion of Lignocellulosic Biomass (Sugar Beet By-Products) and Animal Manure in Long-Term Semi-Continuous Assays. Appl. Sci. 2020, 10, 5126. [Google Scholar] [CrossRef]
- Solarte-Toro, J.C.; Chacón-Pérez, Y.; Cardona-Alzate, C.A. Evaluation of biogas and syngas as energy vectors for heat and power generation using lignocellulosic biomass as raw material. Electron. J. Biotechnol. 2018, 33, 52–62. [Google Scholar] [CrossRef]
- Chala, B.; Oechsner, H.; Latif, S.; Müller, J. Biogas potential of coffee processing waste in Ethiopia. Sustainability 2018, 10, 2678. [Google Scholar] [CrossRef]
- Nelson, M.C.; Morrison, M.; Yu, Z. A meta-analysis of the microbial diversity observed in anaerobic digesters. Bioresour. Technol. 2011, 102, 3730–3739. [Google Scholar] [CrossRef] [PubMed]
- Krause, L.; Diaz, N.N.; Edwards, R.A.; Gartemann, K.-H.; Krömeke, H.; Neuweger, H.; Pühler, A.; Runte, K.J.; Schlüter, A.; Stoye, J.; et al. Taxonomic composition and gene content of a methane-producing microbial community isolated from a biogas reactor. J. Biotechnol. 2008, 136, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Schlüter, A.; Bekel, T.; Diaz, N.N.; Dondrup, M.; Eichenlaub, R.; Gartemann, K.-H.; Krahn, I.; Krause, L.; Krömeke, H.; Kruse, O.; et al. The metagenome of a biogas-producing microbial community of a production-scale biogas plant fermenter analysed by the 454-pyrosequencing technology. J. Biotechnol. 2008, 136, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Gannoun, H.; Omri, I.; Chouari, R.; Eltaief, K.; Keskes, S.; Godon, J.J.; Hamdi, M.; Sghir, A.; Bouallagui, H. Microbial community structure associated with the high loading anaerobic codigestion of olive mill and abattoir wastewaters. Bioresour. Technol. 2016, 201, 337–346. [Google Scholar] [CrossRef]
- Nishimura, T.; Vertès, A.A.; Shinoda, Y.; Inui, M.; Yukawa, H. Anaerobic growth of Corynebacterium glutamicum using nitrate as a terminal electron acceptor. Appl. Microbiol. Biotechnol. 2007, 75, 889–897. [Google Scholar] [CrossRef]
- Gao, R. Microbial diversity in a full-scale anaerobic reactor treating high concentration organic cassava wastewater. Afr. J. Biotechnol. 2012, 11, 6494–6500. [Google Scholar] [CrossRef]
- Guo, W.; Li, Y.; Zhao, K.; Xu, Q.; Jiang, H.; Zhou, H. Performance and Microbial Community Analysis of Anaerobic Digestion of Vinegar Residue with Adding of Acetylene Black or Hydrochar. Waste Biomass Valorization 2020, 11, 3315–3325. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, X.; Zhou, J.; Yuan, Y.; Dai, Y.; Li, D.; Li, Z.; Liu, X.; Yan, Z. The dynamic changes and interactional networks of prokaryotic community between co-digestion and mono-digestions of corn stalk and pig manure. Bioresour. Technol. 2017, 225, 25–33. [Google Scholar] [CrossRef]
- Buhlmann, C.H.; Mickan, B.S.; Jenkins, S.N.; Tait, S.; Kahandawala, T.K.A.; Bahri, P.A. Ammonia stress on a resilient mesophilic anaerobic inoculum: Methane production, microbial community, and putative metabolic pathways. Bioresour. Technol. 2019, 275, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Hülsemann, B.; Zhou, L.; Merkle, W.; Hassa, J.; Müller, J.; Oechsner, H. Biomethane Potential Test: Influence of Inoculum and the Digestion System. Appl. Sci. 2020, 10, 2589. [Google Scholar] [CrossRef]
- Cho, S.; Im, W.; Kim, D.; Kim, M.; Shin, H.; Oh, S. Dry anaerobic digestion of food waste under mesophilic conditions: Performance and methanogenic community analysis. Bioresour. Technol. 2013, 131, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, H.; Qiu, Y.; Ren, L.; Jiang, B. Microbial characteristics in anaerobic digestion process of food waste for methane production–A review. Bioresour. Technol. 2018, 248, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Conklin, A.; Stensel, H.D.; Ferguson, J. Growth kinetics and competition between Methanosarcina and Methanosaeta in mesophilic anaerobic digestion. Water Environ. Res. 2006, 78, 486–496. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effluents | Test Time (d) | Mixture | Test Unit Label |
|---|---|---|---|
| - Olive mill wastewater | 0–73 | 100% PE | P |
| (OMW) | 70% PE + 30% OMW | 70P + 30O | |
| - Piggery effluent | 50% PE + 50% OMW | 50P + 50O | |
| (PE) | 20% PE + 80% OMW | 20P + 80O |
| Effluents | COD (g/L) | TS (g/L) | VS (g/L) | TN (g/L) | NH4+-N (g/L) |
|---|---|---|---|---|---|
| OMW | 106 ± 1 | 32 ± 0 | 26 ± 0 | 0.2 ± 0.0 | 0.001 ± 0.00 |
| PE | 93 ± 5 | 47 ± 1 | 32 ± 1 | 4.9 ± 0.3 | 3.21 ± 0.02 |
| Effluents | Acetic Acid (g/L) | Propionic Acid (g/L Aac) | Isobutyric Acid (g/L Aac) | Butyric Acid (g/L Aac) | Total (g/L Aac) | pH |
|---|---|---|---|---|---|---|
| OMW | 0.21 | 0.06 | 0.11 | 0.16 | 0.55 | 5.1 |
| PE | 1.37 | 0.56 | 2.16 | 1.54 | 5.64 | 7.3 |
| Effluents | Antioxidant Activity (mmol TEAC) | DPPH Inhibition (%) | TP (g/L) |
|---|---|---|---|
| OMW | 0.80 ± 0.07 | 61 ± 3 | 3.1 ± 0.0 |
| PE | 1.11 ± 0.01 | 81 ± 1 | 0.9 ± 0.0 |
| Mixture | COD | Total and Volatile Solids | ||||
|---|---|---|---|---|---|---|
| CODin (g/L) | CODr (%) | TSin (g/L) | TSr (%) | VSin (g/L) | VSr (%) | |
| PE | 93 ± 5 | 63 ± 6 | 47 ± 1 | 17 ± 2 | 32 ± 1 | 29 ± 2 |
| 70P + 30O | 81 ± 3 | 75 ± 0 | 40 ± 0 | 21 ± 2 | 28 ± 0 | 36 ± 4 |
| 50P + 50O | 77 ± 3 | 48 ± 1 | 38 ± 0 | 16 ± 3 | 28 ± 0 | 22 ± 3 |
| 20P + 80O | 73 ± 1 | 29 ± 2 | 33 ± 1 | 21 ± 1 | 25 ± 1 | 25 ± 0 |
| Mixture | TPin (g/L) | TPr (%) | Antioxidant Activity (mmol TEAC) | DPPH Inhibition (%) | ||
|---|---|---|---|---|---|---|
| Initial | Final | Initial | Final | |||
| PE | 0.9 ± 0.0 | 3 ± 0.0 | 1.1 ± 0.0 | 0.6 ± 0.1 | 81 ± 1 | 30 ± 8 |
| 70P + 30O | 1.7 ± 0.0 | 35 ± 0.0 | 1.1 ± 0.0 | 0.4 ± 0.1 | 79 ± 0 | 10 ± 7 |
| 50P + 50O | 2.2 ± 0.0 | 30 ± 0.0 | 1.0 ± 0.0 | 1.3 ± 0.0 | 74 ± 1 | 69 ± 2 |
| 20P + 80O | 2.7 ± 0.0 | 7 ± 0.0 | 0.9 ± 0.0 | 1.3 ± 0.0 | 68 ± 0 | 69 ± 1 |
| Mixture | pH | Total Nitrogen | Ammonia Nitrogen | VFA | ||||
|---|---|---|---|---|---|---|---|---|
| pHin | pHf | TNin (g/L) | TNr (%) | NH4+-Nin (g/L) | NH4+-Nr (%) | Total VFAin (g/L AAc) | Total VFAr (%) | |
| PE | 7.3 | 8.1 | 4.9 ± 0.3 | - | 3.2 ± 0.0 | −21 ± 4 | 5.7 | 85 |
| 70P + 30O | 6.9 | 7.9 | 1.8 ± 0.0 | 3.0 ± 0.0 | 1.3 ± 0.1 | −88 ± 38 | 3.9 | 55 |
| 50P + 50O | 6.7 | 7.3 | 1.3 ± 0.0 | 2.2 ± 1.5 | 0.8 ± 0.3 | −14 ± 6 | 3.7 | −81 |
| 20P + 80O | 6.2 | 5.7 | 0.6 ± 0.0 | 5.7 ± 0.0 | 0.3 ± 0.0 | 13 ± 5 | 2.8 | 52 |
| Mixture | Cumulative CH4 Yield (L CH4/kg SVin) | Energy Content (KWh/kg SVin) |
|---|---|---|
| PE | 434 ± 3 | 4.32 ± 0.03 |
| 70P + 30O | 489 ± 8 | 4.87 ± 0.08 |
| 50P + 50O | 178 ± 29 | 1.77 ± 0.29 |
| 20P + 80O | 6.3 ± 0.3 | 0.06 ± 0.00 |
| Sample | No. of Sequences | OTU | Shannon–Wiener Index | |
|---|---|---|---|---|
| Bacteria | Archaea | |||
| PE (IN) | 193,742 | 1623 | 3.52 | 2.95 |
| PE (OUT) | 206,714 | 779 | 1.38 | 1.45 |
| 70P + 30O (IN) | 181,834 | 1819 | 4.79 | 4.45 |
| 70P + 30O (OUT) | 140,999 | 610 | 1.64 | 2.16 |
| Total | 723,289 | 4831 | - | - |
| Sample | Relative Abundance (%) | |
|---|---|---|
| Bacteria | Archaea | |
| PE IN | 99.73 | 0.26 |
| PE OUT | 99.87 | 0.13 |
| 70P + 30O IN | 99.71 | 0.28 |
| 70P + 30O OUT | 99.92 | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eusébio, A.; Neves, A.; Marques, I.P. Structure of Microbial Communities When Complementary Effluents Are Anaerobically Digested. Appl. Sci. 2021, 11, 1293. https://doi.org/10.3390/app11031293
Eusébio A, Neves A, Marques IP. Structure of Microbial Communities When Complementary Effluents Are Anaerobically Digested. Applied Sciences. 2021; 11(3):1293. https://doi.org/10.3390/app11031293
Chicago/Turabian StyleEusébio, Ana, André Neves, and Isabel Paula Marques. 2021. "Structure of Microbial Communities When Complementary Effluents Are Anaerobically Digested" Applied Sciences 11, no. 3: 1293. https://doi.org/10.3390/app11031293
APA StyleEusébio, A., Neves, A., & Marques, I. P. (2021). Structure of Microbial Communities When Complementary Effluents Are Anaerobically Digested. Applied Sciences, 11(3), 1293. https://doi.org/10.3390/app11031293