
Insights into the Bioactivities and Chemical Analysis of Ailanthus altissima (Mill.) Swingle


 , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Taxonomy and Morphology
3. History and Distribution
4. Biology and Ecology
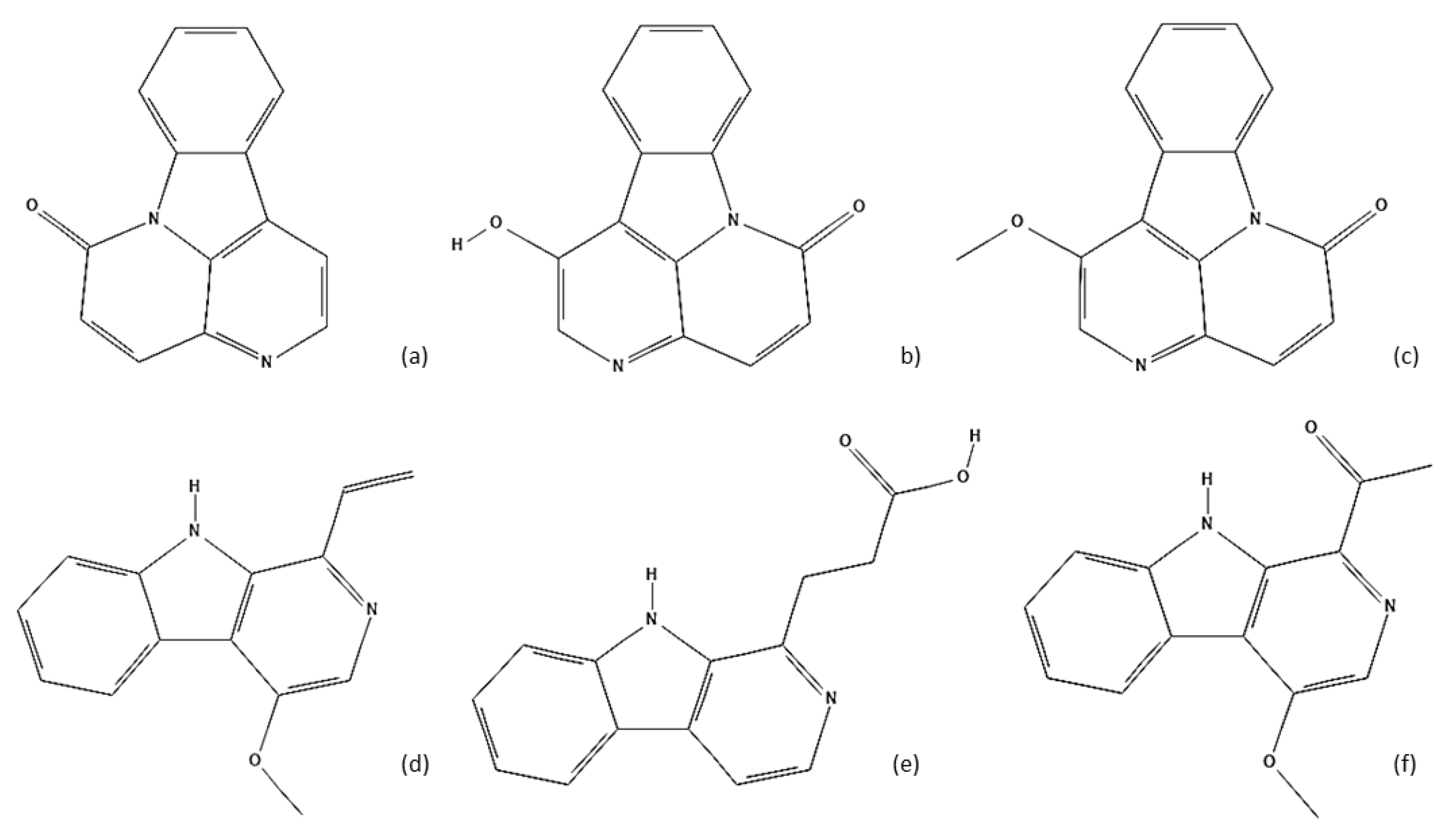
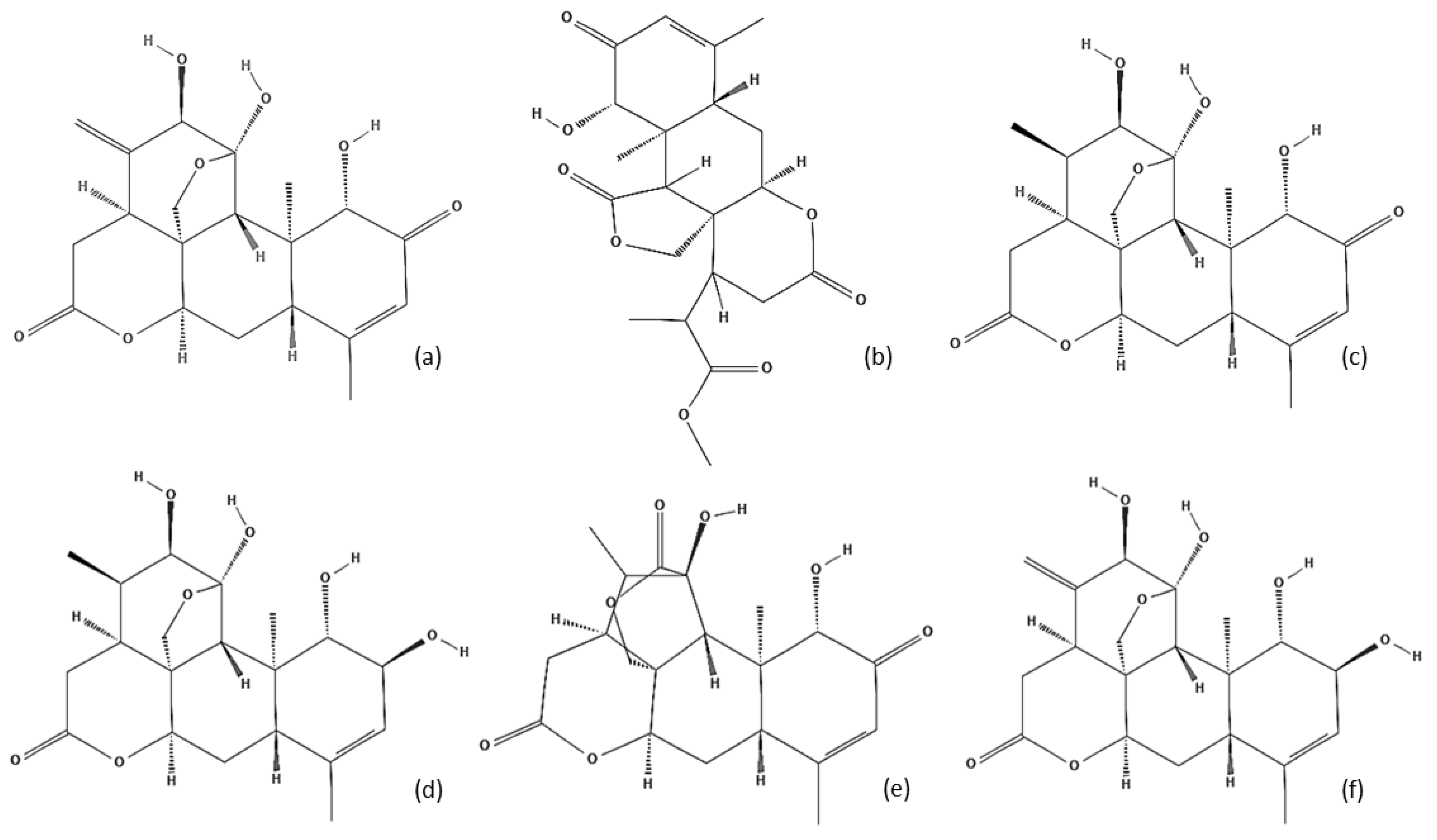
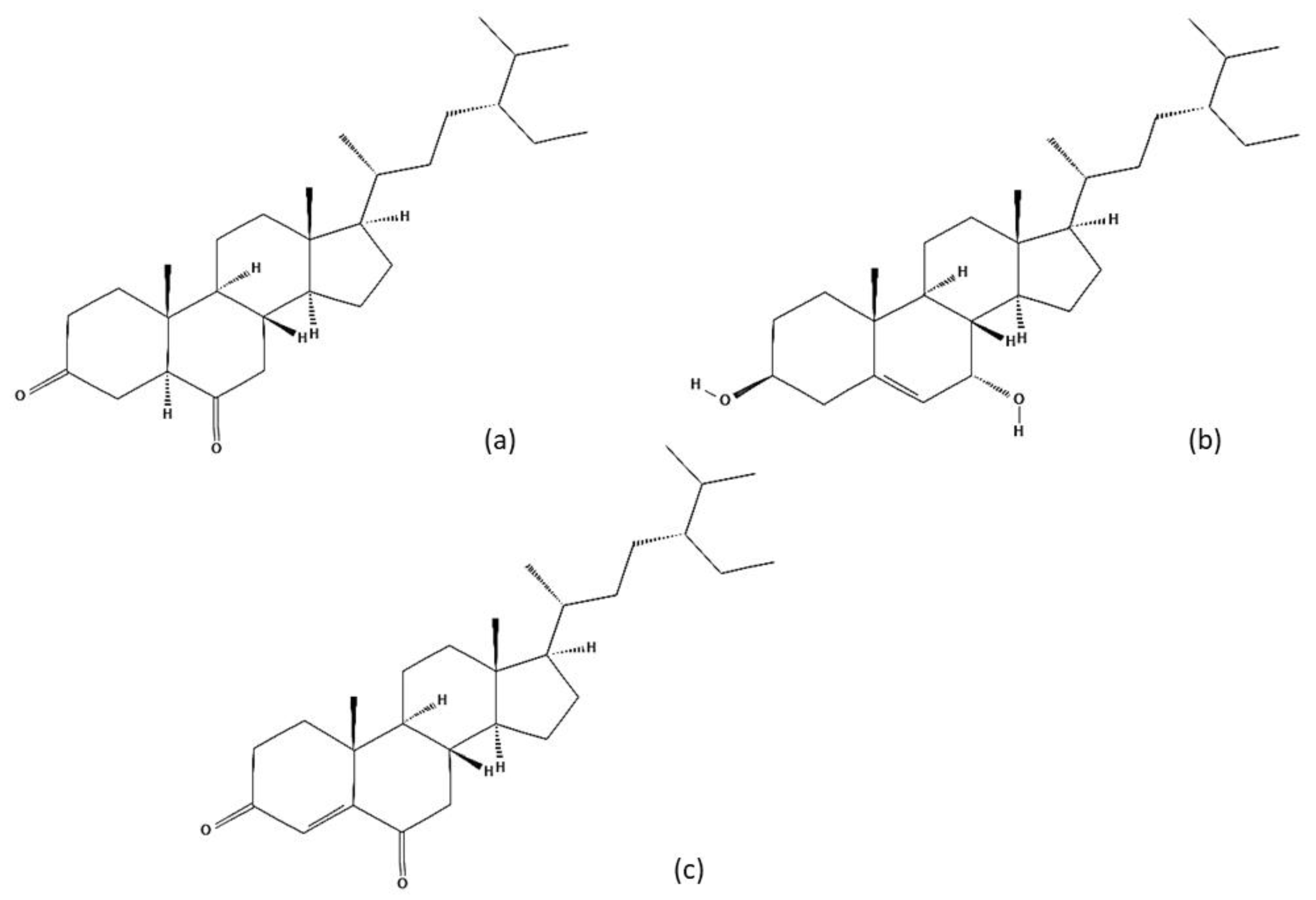
5. Chemical Characterization
6. Biological Properties
6.1. Antimicrobial Activity
6.2. Antioxidant Activity
6.3. Anti-Inflammatory Effects
6.4. Phytotoxic Effects/Phytochemical Activity
6.5. Anti-Tumour and Anti-Viral Activity
6.6. Potential and New Applications
7. Analytical Techniques for the Determination of Active Compounds
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Castro-Díez, P.; Alonso, Á.; Saldaña-López, A.; Granda, E. Effects of widespread non-native trees on regulating ecosystem services. Sci. Total. Environ. 2021, 778, 146141. [Google Scholar] [CrossRef]
- Krumm, F.; Vitkova, L. Introduced Tree Species in European Forests: Opportunities and Challenges; European Forest Institute: Joensuu, Finland, 2016; ISBN 9789525980318. [Google Scholar]
- Castro-Díez, P.; Godoy, O.; Saldaña, A.; Richardson, D.M. Predicting invasiveness of Australian acacias on the basis of their native climatic affinities, life history traits and human use. Divers. Distrib. 2011, 17, 934–945. [Google Scholar] [CrossRef]
- De Wit, M.P.; Crookes, D.J.; Van Wilgen, B.W. Conflicts of interest in environmental management: Estimating the costs and benefits of a tree invasion. Biol. Invasions 2001, 3, 167–178. [Google Scholar] [CrossRef]
- Dickie, I.A.; Bennett, B.M.; Burrows, L.E.; Nuñez, M.A.; Peltzer, D.A.; Porté, A.; Richardson, D.M.; Rejmánek, M.; Rundel, P.W.; van Wilgen, B.W. Conflicting values: Ecosystem services and invasive tree management. Biol. Invasions 2014, 16, 705–719. [Google Scholar] [CrossRef]
- Sladonja, B.; Sušek, M.; Guillermic, J. Review on Invasive Tree of Heaven (Ailanthus altissima (Mill.) Swingle) Conflicting Values: Assessment of Its Ecosystem Services and Potential Biological Threat. Environ. Manag. 2015, 56, 1009–1034. [Google Scholar] [CrossRef] [PubMed]
- Kowarik, I.; Säumel, I. Biological flora of Central Europe: Ailanthus altissima (Mill.) Swingle. Perspect. Plant Ecol. Evol. Syst. 2007, 8, 207–237. [Google Scholar] [CrossRef]
- Constán-Nava, S.; Bonet, A.; Pastor, E.; Lledó, M.J. Long-term control of the invasive tree Ailanthus altissima: Insights from Mediterranean protected forests. For. Ecol. Manag. 2010, 260, 1058–1064. [Google Scholar] [CrossRef]
- Almeida, M.T.; Mouga, T.; Barracosa, P. The weathering ability of higher plants. The case of Ailanthus altissima (Miller) Swingle. Int. Biodeterior. Biodegrad. 1994, 33, 333–343. [Google Scholar] [CrossRef]
- Kožuharova, E.; Lebanova, H.; Getov, I.; Benbassat, N.; Kochmarov, V. Ailanthus altissima (Mill.) Swingle—A terrible invasive pest in Bulgaria or potential useful medicinal plant? Bothalia J. 2014, 44, 213–230. [Google Scholar]
- International Plant Nutrition Institute. International Plant Names Index. Available online: http://www.ipni.org (accessed on 1 October 2021).
- Kundu, P.; Laskar, S. A brief resume on the genus Ailanthus: Chemical and pharmacological aspects. Phytochem. Rev. 2010, 9, 379–412. [Google Scholar] [CrossRef]
- Hu, S.Y. Ailanthus. Arnoldia 1979, 39, 29–50. [Google Scholar]
- Castroviejo, S. (coord. gen.). In Flora Iberica; Real Jardín Botánico, Spanish National Research Council: Madrid, Spain, 1986–2012.
- Bashir, A.; Mohi-ud-din, R.; Farooq, S.; Bhat, Z.A. Pharmacognostic Standardization and Phytochemical Evaluation of Ailanthus altissima (Mill.) Swingle leaves. J. Drug Deliv. Ther. 2019, 9, 179–183. [Google Scholar]
- Pan, E.; Bassuk, N. Establishment and Distribution of Ailanthus altissima in the Urban Environment. J. Environ. Hortic. 1986, 4, 1–4. [Google Scholar] [CrossRef]
- Arnaboldi, F.; Conedera, M.; Maspoli, G. Distribuzione e potenziale invasivo di Ailanthus altissima (Mill.) Swingle nel Ticino centrale. Boll. Soc. Ticin. Sci. Nat. 2002, 90, 93–101. [Google Scholar]
- Tabacchi, E.; Planty-Tabacchi, A.M. Recent changes in Riparian vegetation: Possible consequences on dead wood processing along rivers. River Res. Appl. 2003, 19, 251–263. [Google Scholar] [CrossRef]
- Facelli, J.M.; Pickett, S.T.A.; Facelli, J.M. Indirect Effects of Litter on Woody Seedlings Subject to Herb Competition. Oikos 1991, 62, 129. [Google Scholar] [CrossRef]
- Facelli, J.M. Multiple indirect effects of plant litter affect the establishment of woody seedlings in old fields. Ecology 1994, 75, 1727–1735. [Google Scholar] [CrossRef]
- Ranft, H.; Dässler, H.-G. Rauchhärtetest an Gehölzen im SO2-Kabinenversuch: Fume-Hardiness Test Carried out on Woods in a SO2-Cabin-Trial. Flora 1970, 159, 573–588. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Ma, S.; Zhao, Q.; Wu, J.; Duan, L.; Xie, Y.; Wang, S. Traditional uses, phytochemistry, and pharmacology of Ailanthus altissima (Mill.) Swingle bark: A comprehensive review. J. Ethnopharmacol. 2021, 275, 114121. [Google Scholar] [CrossRef]
- Du, Y.Q.; Yan, Z.Y.; Chen, J.J.; Wang, X.B.; Huang, X.X.; Song, S.J. The identification of phenylpropanoids isolated from the root bark of Ailanthus altissima (Mill.) Swingle. Nat. Prod. Res. 2021, 35, 1139–1146. [Google Scholar] [CrossRef]
- Yang, X.L.; Yuan, Y.L.; Zhang, D.M.; Li, F.; Ye, W.C. Shinjulactone O, a new quassinoid from the root bark of Ailanthus altissima. Nat. Prod. Res. 2014, 28, 1432–1437. [Google Scholar] [CrossRef]
- Zhang, D.D.; Bai, M.; Yan, Z.Y.; Huang, X.X.; Song, S.J. Chemical constituents from Ailanthus altissima (Mill.) Swingle and chemotaxonomic significance. Biochem. Syst. Ecol. 2020, 93, 104174. [Google Scholar] [CrossRef]
- Ni, J.C.; Shi, J.T.; Tan, Q.W.; Chen, Q.J. Two new compounds from the fruit of Ailanthus altissima. Nat. Prod. Res. 2019, 33, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.C.; Shi, J.T.; Tan, Q.W.; Chen, Q.J. Phenylpropionamides, piperidine, and phenolic derivatives from the fruit of Ailanthus altissima. Molecules 2017, 22, 2107. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.C.; Shao, J.H.; Li, X.; Xu, J.; Zhang, P. Antimicrobial constituents from fruits of Ailanthus altissima SWINGLE. Arch. Pharm. Res. 2005, 28, 1147–1151. [Google Scholar] [CrossRef]
- Bory, G.; Clair-Maczulajtys, D. Composition du nectar et rôle des nectaires extrafloraux chez l’Ailanthus glandulosa. Can. J. Bot. 1986, 64, 247–253. [Google Scholar] [CrossRef]
- Heisey, R.M.; Heisey, T.K. Herbicidal effects under field conditions of Ailanthus altissima bark extract, which contains ailanthone. Plant Soil 2003, 256, 85–99. [Google Scholar] [CrossRef]
- De Feo, V.; De Martino, L.; Quaranta, E.; Pizza, C. Isolation of phytotoxic compounds from tree-of-heaven (Ailanthus altissima swingle). J. Agric. Food Chem. 2003, 51, 1177–1180. [Google Scholar] [CrossRef]
- Tan, Q.W.; Ni, J.C.; Shi, J.T.; Zhu, J.X.; Chen, Q.J. Two novel quassinoid glycosides with antiviral activity from the samara of Ailanthus altissima. Molecules 2020, 25, 5679. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, W.J.; Su, C.; Zhang, D.M.; Xu, L.P.; He, R.R.; Wang, L.; Zhang, J.; Zhang, X.Q.; Ye, W.C. Cytotoxic quassinoids from Ailanthus altissima. Bioorg. Med. Chem. Lett. 2013, 23, 654–657. [Google Scholar] [CrossRef]
- Okunade, A.L.; Bikoff, R.E.; Casper, S.J.; Oksman, A.; Goldberg, D.E.; Lewis, W.H. Antiplasmodial activity of extracts and quassinoids isolated from seedlings of Ailanthus altissima (Simaroubaceae). Phyther. Res. 2003, 17, 675–677. [Google Scholar] [CrossRef] [PubMed]
- Fukamiya, N.; Lee, K.H.; Muhammad, I.; Murakami, C.; Okano, M.; Harvey, I.; Pelletier, J. Structure-activity relationships of quassinoids for eukaryotic protein synthesis. Cancer Lett. 2005, 220, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Al-Snafi, A.E. The pharmacological importance of Antirrhinum majus—A review. Asian J. Pharm. Sci. Technol. 2015, 5, 313–320. [Google Scholar]
- Swingle, M. Analgesic, antipyretic and antiulcer activities of Ailanthus altissima. Phytopharmacology 2012, 3, 341–350. [Google Scholar]
- Ghannoum, M.A.; Rice, L.B. Antifungal agents: Mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clin. Microbiol. Rev. 1999, 12, 501–517. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Kim, E.L.; Kang, S.C. Antibacterial and antioxidant properties of Ailanthus altissima swingle leave extract to reduce foodborne pathogens and spoiling bacteria. J. Food Saf. 2009, 29, 499–510. [Google Scholar] [CrossRef]
- Albouchi, F.; Hassen, I.; Casabianca, H.; Hosni, K. Phytochemicals, antioxidant, antimicrobial and phytotoxic activities of Ailanthus altissima (Mill.) Swingle leaves. S. Afr. J. Bot. 2013, 87, 164–174. [Google Scholar] [CrossRef]
- Aissani, N.; Jabri, M.; Mabrouk, M.; Sebai, H. Antioxidant Potential and Antimicrobial Activity of Ailanthus altissima (Mill.) Swingle Extracts against Pseudomonas aeruginosa. J. Mol. Biol. Biotech. 2018, 3, 3. [Google Scholar]
- Končić, M.Z.; Kremer, D.; Gruz, J.; Strnad, M.; Biševac, G.; Kosalec, I.; Šamec, D.; Piljac-Žegarac, J.; Karlović, K. Antioxidant and antimicrobial properties of Moltkia petraea (Tratt.) Griseb. flower, leaf and stem infusions. Food Chem. Toxicol. 2010, 48, 1537–1542. [Google Scholar] [CrossRef]
- Chanwitheesuk, A.; Teerawutgulrag, A.; Kilburn, J.D.; Rakariyatham, N. Antimicrobial gallic acid from Caesalpinia mimosoides Lamk. Food Chem. 2007, 100, 1044–1048. [Google Scholar] [CrossRef]
- Betts, J.W.; Kelly, S.M.; Haswell, S.J. Antibacterial effects of theaflavin and synergy with epicatechin against clinical isolates of Acinetobacter baumannii and Stenotrophomonas maltophilia. Int. J. Antimicrob. Agents 2011, 38, 421–425. [Google Scholar] [CrossRef]
- Tian, F.; Li, B.; Ji, B.; Yang, J.; Zhang, G.; Chen, Y.; Luo, Y. Antioxidant and antimicrobial activities of consecutive extracts from Galla chinensis:The polarity affects the bioactivities. Food Chem. 2009, 113, 173–179. [Google Scholar] [CrossRef]
- Hosni, K.; Hassen, I.; Sebei, H.; Casabianca, H. Secondary metabolites from Chrysanthemum coronarium (Garland) flowerheads: Chemical composition and biological activities. Ind. Crop. Prod. 2013, 44, 263–271. [Google Scholar] [CrossRef]
- Polumbryk, M.; Ivanov, S.; Polumbryk, O. Antioxidants in food systems. Mechanism of action. Ukr. J. Food Sci. 2013, 1, 15–40. [Google Scholar]
- Olszowy, M. What is responsible for antioxidant properties of polyphenolic compounds from plants? Plant Physiol. Biochem. 2019, 144, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A.; Cimpeanu, C.; Predoi, G. Antioxidant Capacity Determination in Plants and Plant-Derived Products: A Review. Oxid. Med. Cell. Longev. 2016, 2016, 9130976. [Google Scholar] [CrossRef] [PubMed]
- Luís, Â.; Gil, N.; Amaral, M.E.; Domingues, F.; Duarte, A.P. Ailanthus altissima (Miller) Swingle: A source of bioactive compounds with antioxidant activity. BioResources 2012, 7, 2105–2120. [Google Scholar] [CrossRef]
- Lungu, L.; Savoiu, M.R.; Manolescu, B.N.; Farcasanu, I.C.; Popa, C.V. Phytotoxic and Antioxidant activities of leaf extracts of Ailanthus altissima swingle. Rev. Chim. 2016, 67, 1928–1931. [Google Scholar]
- EL Ayeb-Zakhama, A.; Chahdoura, H.; Ziani, B.E.C.; Snoussi, M.; Khemiss, M.; Flamini, G.; Harzallah-Skhiri, F. Ailanthus altissima (Miller) Swingle seed oil: Chromatographic characterization by GC-FID and HS-SPME-GC-MS, physicochemical parameters, and pharmacological bioactivities. Environ. Sci. Pollut. Res. 2019, 26, 14137–14147. [Google Scholar] [CrossRef]
- Yeung, Y.; Aziz, F.; Guerrero-Castilla, A.; Arguelles, S. Signaling Pathways in Inflammation and Anti-inflammatory Therapies. Curr. Pharm. Des. 2018, 24, 1449–1484. [Google Scholar] [CrossRef]
- Jin, M.H.; Yook, J.; Lee, E.; Lin, C.X.; Quan, Z.; Son, K.H.; Bae, K.H.; Kim, H.P.; Kang, S.S.; Chang, H.W. Anti-inflammatory Activity of Ailanthus altissima in Ovalbumin-Induced Lung Inflammation. Biol. Pharm. Bull. 2006, 29, 884–888. [Google Scholar] [CrossRef]
- Cho, S.-K.; Jeong, M.; Jang, D.; Choi, J.-H. Anti-inflammatory Effects of Canthin-6-one Alkaloids from Ailanthus altissima. Planta Med. 2018, 84, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Lee, J.S.; Sezirahiga, J.; Kwon, J.; Jeong, M.; Lee, D.; Choi, J.-H.; Jang, D.S. A New Canthinone-Type Alkaloid Isolated from Ailanthus altissima Swingle. Molecules 2016, 21, 642. [Google Scholar] [CrossRef]
- Kang, T.-H.; Choi, I.-Y.; Kim, S.-J.; Moon, P.-D.; Seo, J.-U.; Kim, J.-J.; An, N.-H.; Kim, S.-H.; Kim, M.-H.; Um, J.-Y.; et al. Ailanthus altissima swingle has anti-anaphylactic effect and inhibits inflammatory cytokine expression via suppression of nuclear factor-kappaB activation. In Vitro Cell. Dev. Biol. Anim. 2010, 46, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Park, Y.; Li, M.; Kim, Y.K.; Lee, S.; Son, S.Y.; Lee, S.; Lee, J.S.; Lee, C.H.; Park, H.H.; et al. Anti-inflammatory effect of Ailanthus altissima (Mill.) Swingle leaves in lipopolysaccharide-stimulated astrocytes. J. Ethnopharmacol. 2021, 114258. [Google Scholar] [CrossRef] [PubMed]
- Fiaschetti, G.; Grotzer, M.A.; Shalaby, T.; Castelletti, D.; Arcaro, A. Quassinoids: From traditional drugs to new cancer therapeutics. Curr. Med. Chem. 2011, 18, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Morré, D.J.; Grieco, P.A.; Morré, D.M. Mode of action of the anticancer quassinoids—Inhibition of the plasma membrane NADH oxidase. Life Sci. 1998, 63, 595–604. [Google Scholar] [CrossRef]
- Corrêa, L.R.; Soares, G.L.G.; Fett-Neto, A.G. Allelopathic potential of Psychotria leiocarpa, a dominant understorey species of subtropical forests. S. Afr. J. Bot. 2008, 74, 583–590. [Google Scholar] [CrossRef][Green Version]
- Ahmed, H.M.; Yeh, J.-Y.; Tang, Y.-C.; Cheng, W.T.-K.; Ou, B.-R. Molecular screening of Chinese medicinal plants for progestogenic and anti-progestogenic activity. J. Biosci. 2014, 39, 453–461. [Google Scholar] [CrossRef]
- Lawrence, J.; Colwell, A.; Sexton, O. The Ecological Impact of Allelopathy in Ailanthus altissima (Simaroubaceae). Am. J. Bot. 1991, 78, 948–958. [Google Scholar] [CrossRef]
- Chen, Y.; Zhu, L.; Yang, X.; Wei, C.; Chen, C.; He, Y.; Ji, Z. Ailanthone induces G2/M cell cycle arrest and apoptosis of SGC-7901 human gastric cancer cells. Mol. Med. Rep. 2017, 16, 6821–6827. [Google Scholar] [CrossRef]
- Ding, H.; Yu, X.; Hang, C.; Gao, K.; Lao, X.; Jia, Y.; Yan, Z. Ailanthone: A novel potential drug for treating human cancer. Oncol. Lett. 2020, 20, 1489–1503. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Ma, X.; Lu, J.; Zhang, Y.; Liu, M.; Wang, X. Preclinical toxicology and toxicokinetic evaluation of ailanthone, a natural product against castration-resistant prostate cancer, in mice. Fitoterapia 2019, 136, 104161. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.W.; Ouyang, M.A.; Wu, Z.J. A new seco-neolignan glycoside from the root bark of Ailanthus altissima. Nat. Prod. Res. 2012, 26, 1375–1380. [Google Scholar] [CrossRef]
- Demasi, S.; Caser, M.; Vanara, F.; Fogliatto, S.; Vidotto, F.; Negre, M.; Trotta, F.; Scariot, V. Ailanthone from Ailanthus altissima (Mill.) Swingle as potential natural herbicide. Sci. Hortic. 2019, 257, 108702. [Google Scholar] [CrossRef]
- Caser, M.; Demasi, S.; Caldera, F.; Dhakar, N.K.; Trotta, F.; Scariot, V. Activity of Ailanthus altissima (Mill.) Swingle Extract as a Potential Bioherbicide for Sustainable Weed Management in Horticulture. Agronomy 2020, 10, 965. [Google Scholar] [CrossRef]
- Baptista, P.; Costa, A.P.; Simões, R.; Amaral, M.E. Ailanthus altissima: An alternative fiber source for papermaking. Ind. Crop. Prod. 2014, 52, 32–37. [Google Scholar] [CrossRef]
- Ferreira, P.J.T.; Gamelas, J.A.F.; Carvalho, M.G.V.S.; Duarte, G.V.; Canhoto, J.M.P.L.; Passas, R. Evaluation of the papermaking potential of Ailanthus altissima. Ind. Crop. Prod. 2013, 42, 538–542. [Google Scholar] [CrossRef]
- He, Q.; Xiao, H.; Li, J.; Liu, Y.; Jia, M.; Wang, F.; Zhang, Y.; Wang, W.; Wang, S. Fingerprint analysis and pharmacological evaluation of Ailanthus altissima. Int. J. Mol. Med. 2018, 41, 3024–3032. [Google Scholar] [CrossRef]
- Mastelić, J.; Jerković, I. Volatile constituents from the leaves of young and old Ailanthus altissima (Mill.) swingle tree. Croat. Chem. Acta 2002, 75, 189–197. [Google Scholar]
- Pedersini, C.; Bergamin, M.; Aroulmoji, V.; Baldini, S.; Picchio, R.; Pesce, P.G.; Ballarin, L.; Murano, E. Herbicide activity of extracts from Ailanthus altissima (Simaroubaceae). Nat. Prod. Commun. 2011, 6, 593–596. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, M.; Zhang, Z. Quantitative proteomics reveals the antifungal effect of canthin-6-one isolated from Ailanthus altissima against Fusarium oxysporum f. sp. cucumerinum In Vitro. PLoS ONE 2021, 16, e0250712. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Ye, J.; Jiang, B. Characterization of volatile constituents from the flowers and leaves of Ailanthus altissima by dynamic head-space collection/GC-MS. Asian J. Chem. 2014, 26, 6737–6739. [Google Scholar] [CrossRef]
- Caboni, P.; Ntalli, N.G.; Aissani, N.; Cavoski, I.; Angioni, A. Nematicidal activity of (E,E)-2,4-decadienal and (E)-2-decenal from Ailanthus altissima against Meloidogyne javanica. J. Agric. Food Chem. 2012, 60, 1146–1151. [Google Scholar] [CrossRef] [PubMed]
- Vidović, M.; Morina, F.; Milić, S.; Jovanović, S.V. An improved HPLC-DAD method for simultaneously measuring phenolics in the leaves of Tilia platyphyllos and Ailanthus altissima. Bot. Serbica 2015, 39, 177–186. [Google Scholar]
- Poljuha, D.; Sladonja, B.; Šola, I.; Dudaš, S.; Bilić, J.; Rusak, G.; Motlhatlego, K.E.; Eloff, J.N. Phenolic composition of leaf extracts of Ailanthus altissima (simaroubaceae) with antibacterial and antifungal activity equivalent to standard antibiotics. Nat. Prod. Commun. 2017, 12, 1609–1612. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Parts Used (Source) | |
|---|---|---|
| Alkaloids | 1-Ethyl-4-methoxy-β-carboline | Root bark |
| β-Carboline-1-propionic acid | Root bark | |
| Methyl-4-methoxy-β-caboline carboxylate | Root bark | |
| 1-(1′-2′-Dihydroxyethyl)-4-methoxy-β-carboline | Root bark | |
| 1-(2′-Hydroxyethyl)-4-methoxy-β-carboline/Crenatine | Root bark | |
| 4-Methoxy-1-vinyl-β-carboline/Dehydrocrenatine | Dried leaves | |
| 1-Methoxycarbonyl-4,8-dimethoxy-β-caboline | Leaves | |
| 1-Methoxycarbonyl-β-carboline/1-caromethoxy-β-carboline | Leaves | |
| 1-(1-Hydroxy-2-methoxy)-ethyl-4-methoxy-β-carboline | Root bark | |
| 1-Carbamoyl-β-carboline | Root bark | |
| 1-Acetyl-4-methoxy-β-carboline | Root bark | |
| Canthine-6-one | Root bark, leaves and wood | |
| 1-Hydroxycanthine-6-one | Root bark | |
| 1-Methoxy-canthine-6-one | Root bark, dried leaves and wood | |
| 5-Hydroxymethylcanthine-6-one | Root bark | |
| Canthine-6-one-3-N-oxide | Wood | |
| 1-Methoxycanthine-6-one-3-N-oxide | Root bark | |
| Terpenoids | Ailanthone | Stem bark |
| Chapparinone | Seed | |
| Chapparin | Aerial part | |
| Shinjulactone and Shinjulactone B | Stem bark | |
| Shinjulactone A | Seed | |
| Shinjulactone C, D, E, F, M, N, G, H, I, J, K and L | Root bark | |
| ∆13(18) -Dehydroglaucarubinone | Stem bark | |
| ∆13(18) -Dehydroglaucarubolone | Stem bark | |
| Ailantinol A e B | Stem bark | |
| Ailantinol C, D, E, F, G and H | Aerial part | |
| Shinjuglycoside A, B, C and D | Seed | |
| Shinjuglycoside E and F | Root bark | |
| Shinjudilactone | Root bark | |
| Cycloart-25-ene-3β-24R-diol and Cycloart-25-ene-3β-24S-diol | Fruits | |
| 9,19-Cyclolanost-23(Z)ene-3β,25-diol | Fruits | |
| Ailantholide | Seed | |
| 3-epi-ursolic acid | Fruits | |
| 12β, 20 (S)-dihydroxy dammar-24-en-3-one | Fruits | |
| Steroids | b-Sitosterol | Leaves and fruits |
| b-Sitosterol glucoside/Daucosterol/Sitosterol-3-Ob-D-glucoside | Leaves and fruits | |
| Ailanthusterol A and B | Seed | |
| 5α-Stigmastane-3,6-dione | Fruits | |
| 5α-Stigmastane-3β, 6β-diol | Fruits | |
| 6α-Hydroxy-stigmast-4-en-3-one | Fruits | |
| Stigmast-4-ene-3β, 6α-diol | Fruits | |
| Stigmast-4-ene-3,6-dione | Fruits | |
| 6β-Hydroxy-stigmast-4-ene-3-one | Fruits | |
| Stigmast-4-ene-3β, 6β-diol | Fruits | |
| 3β-Hydroxy-stigmast-5-en-7-one | Fruits | |
| Stigmast-5-ene-3β, 7α-diol | Fruits | |
| Stigmast-5-ene-3β, 7α, 20ζ-triol | Fruits | |
| Flavonoids | Quercetin | Leaves |
| Kaempferol | Leaves | |
| Isoquercetin/Quercetin-3-O-glucoside | Leaves | |
| Kaempferol-3-O-glucoside | Leaves | |
| Rutin | Leaves | |
| Luteolin-7-O-b-(600 galloylglucopyranoside) | Leaves | |
| Miscellaneous compounds | 1-O-b-D-Glucopyranosyl-(2S,3R,4E,9E)-2-(20-R-hydroxyhexadecenoy)-4,9-octadecadiene-1,3-diol | Fruits |
| Ceryl alcohol | Leaves | |
| Ethyl gallate | Leaves | |
| Altissimacoumarin A and B | Bark | |
| Coumarin | Bark | |
| Isofraxidin | Bark | |
| Scopoletin | Bark |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caramelo, D.; Pedro, S.I.; Marques, H.; Simão, A.Y.; Rosado, T.; Barroca, C.; Gominho, J.; Anjos, O.; Gallardo, E. Insights into the Bioactivities and Chemical Analysis of Ailanthus altissima (Mill.) Swingle. Appl. Sci. 2021, 11, 11331. https://doi.org/10.3390/app112311331
Caramelo D, Pedro SI, Marques H, Simão AY, Rosado T, Barroca C, Gominho J, Anjos O, Gallardo E. Insights into the Bioactivities and Chemical Analysis of Ailanthus altissima (Mill.) Swingle. Applied Sciences. 2021; 11(23):11331. https://doi.org/10.3390/app112311331
Chicago/Turabian StyleCaramelo, Débora, Soraia I. Pedro, Hernâni Marques, Ana Y. Simão, Tiago Rosado, Celina Barroca, Jorge Gominho, Ofélia Anjos, and Eugenia Gallardo. 2021. "Insights into the Bioactivities and Chemical Analysis of Ailanthus altissima (Mill.) Swingle" Applied Sciences 11, no. 23: 11331. https://doi.org/10.3390/app112311331
APA StyleCaramelo, D., Pedro, S. I., Marques, H., Simão, A. Y., Rosado, T., Barroca, C., Gominho, J., Anjos, O., & Gallardo, E. (2021). Insights into the Bioactivities and Chemical Analysis of Ailanthus altissima (Mill.) Swingle. Applied Sciences, 11(23), 11331. https://doi.org/10.3390/app112311331