
Identification of Scopoletin and Chlorogenic Acid as Potential Active Components in Sunflower Calathide Enzymatically Hydrolyzed Extract towards Hyperuricemia

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Primary Extract (SCHE)
2.2. Identification by UPLC–Q-Orbitrap MS
2.3. Cell Culture
2.4. Generation of Reactive Oxygen Species (ROS)
2.5. Western Blot
2.6. Effect of XO Inhibition In Vitro
2.7. Semi Flexible Docking of Xanthine Oxidase (XO)
2.8. Statistical Analysis
3. Results
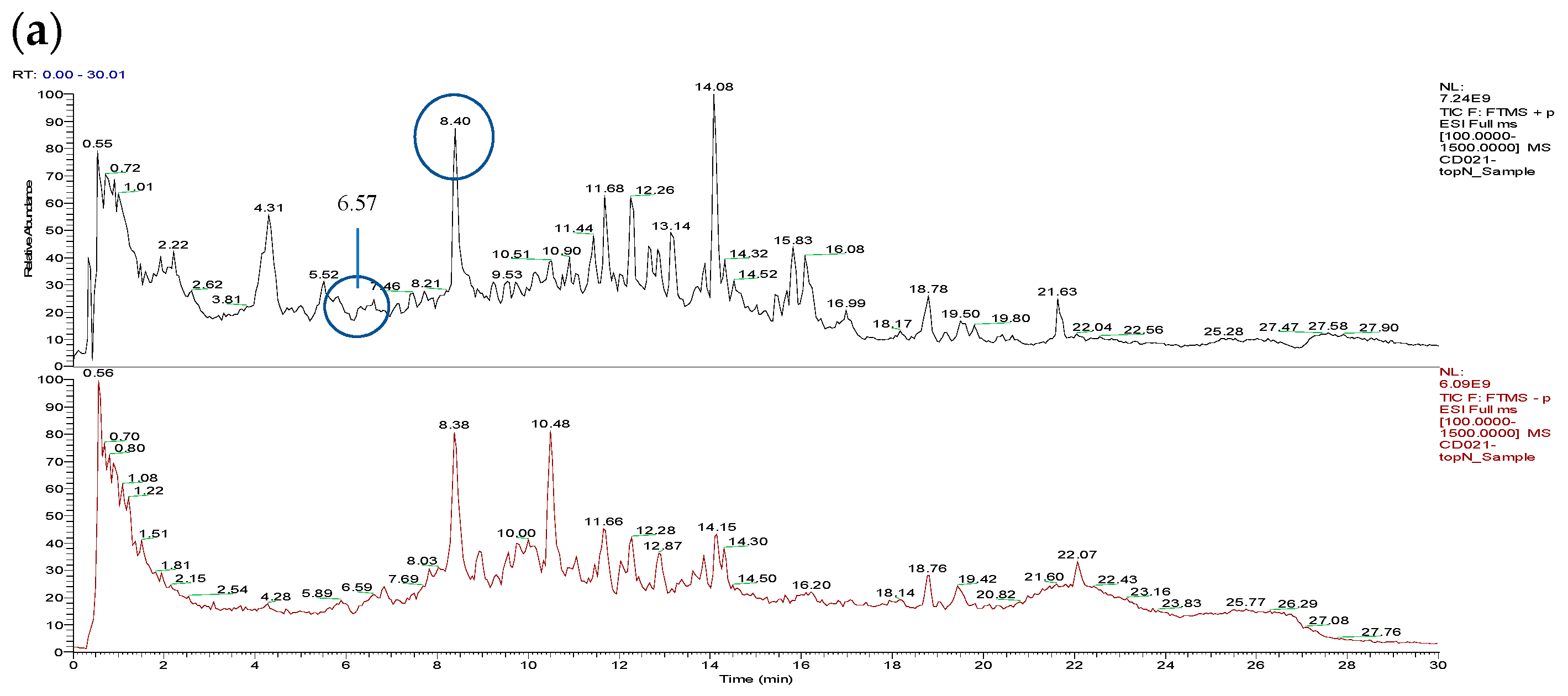
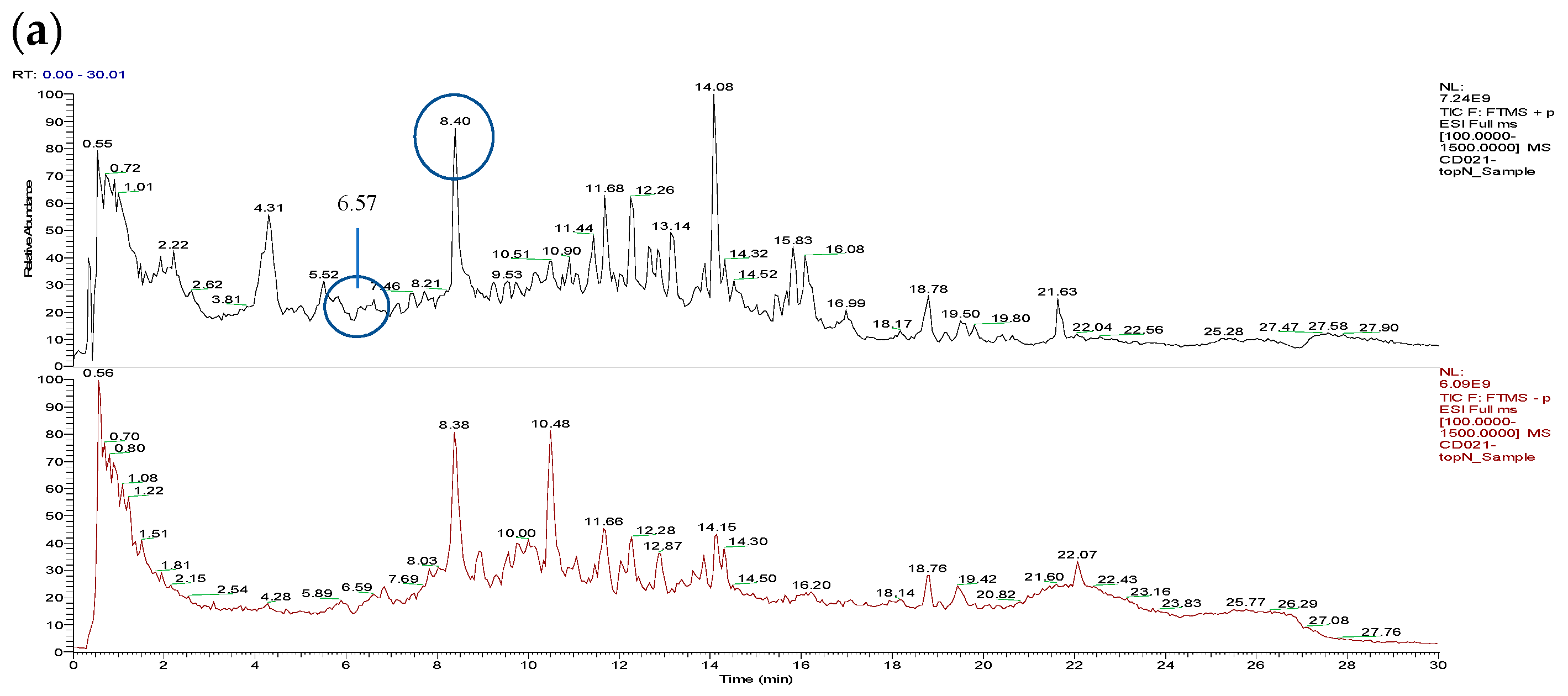
3.1. Total Ion Chromatograms (TIC)
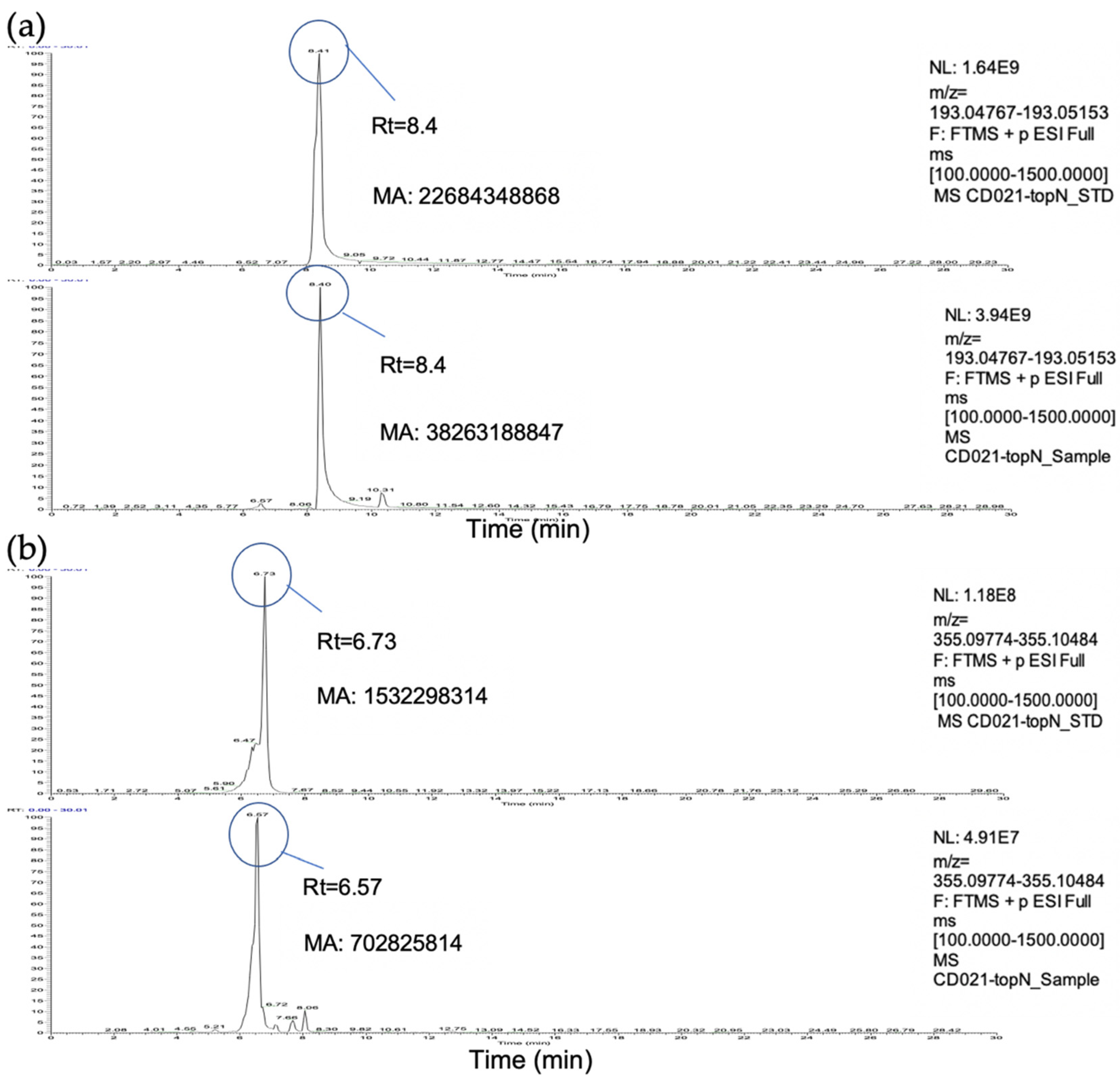
3.2. Identification and Quantification of Scopoletin and Chlorogenic Acid
3.3. Semi-Quantitative Analysis of ROS Generation
3.4. Modulation of Urate Transporters
3.5. Inhibitory Effects of Compounds on UA Formation
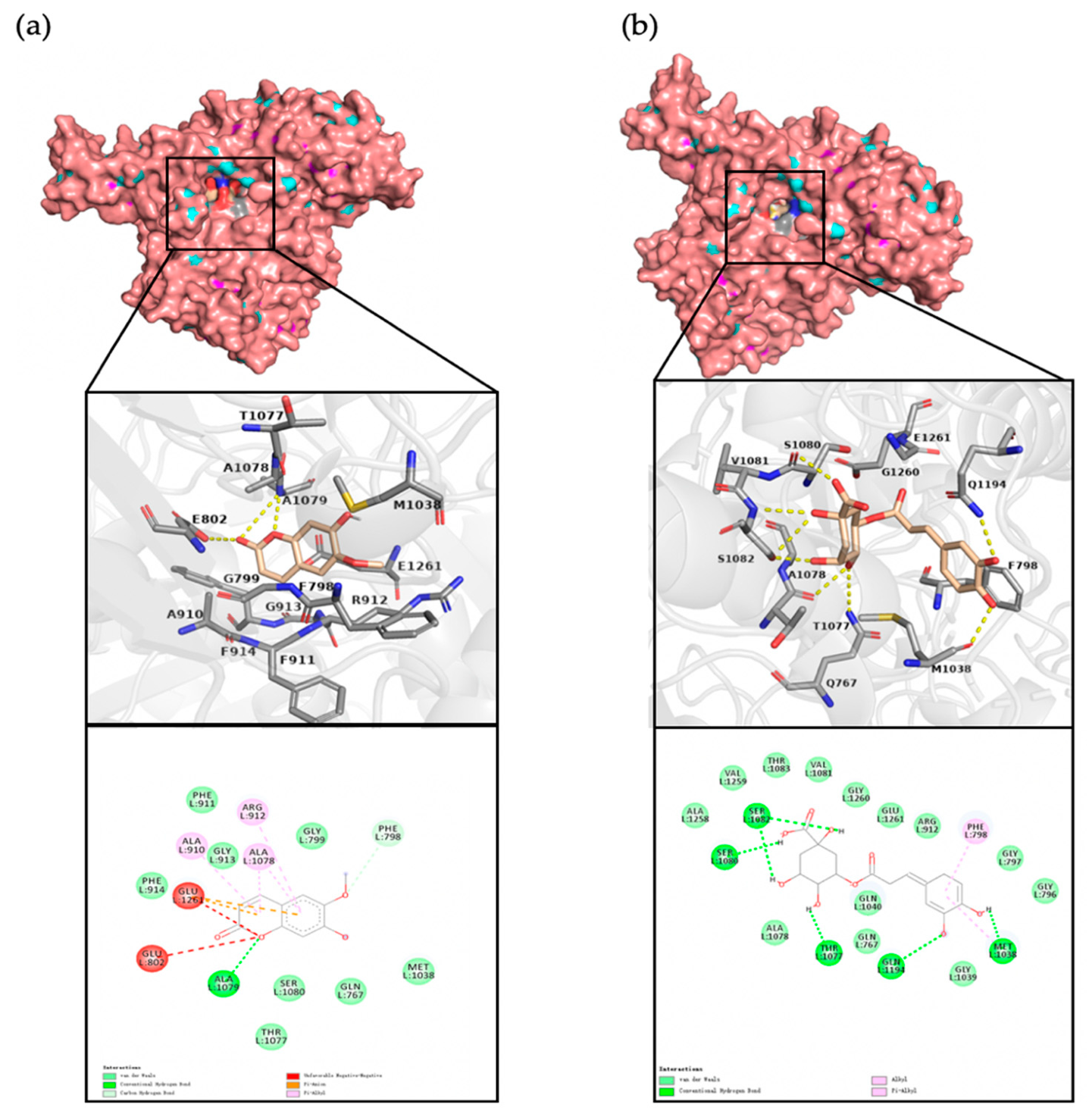
3.6. Molecular Docking and Residue Interactions
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Benn, C.L.; Dua, P.; Gurrell, R.; Loudon, P.; Pike, A.; Storer, R.I.; Vangjeli, C. Physiology of Hyperuricemia and Urate-Lowering Treatments. Front. Med. (Lausanne) 2018, 5, 160. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.K.; Ford, E.S. Prevalence of the metabolic syndrome in individuals with hyperuricemia. Am. J. Med. 2007, 120, 442–447. [Google Scholar] [CrossRef]
- Mendez Landa, C.E. Renal Effects of Hyperuricemia. Contrib. Nephrol. 2018, 192, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.A.; Schumacher, H.R.; Wortmann, R.L.; MacDonald, P.A.; Eustace, D.; Palo, W.A.; Streit, J.; Joseph-Ridge, N. Febuxostat Compared with Allopurinol in Patients with Hyperuricemia and Gout. N. Engl. J. Med. 2005, 353, 2450–2461. [Google Scholar] [CrossRef] [Green Version]
- Iglesias, M.T.; Lozano, J.E. Extraction and characterization of sunflower pectin. J. Food Eng. 2004, 62, 215–223. [Google Scholar] [CrossRef]
- Sobol, I.V. The Use of Highly Purified Sunflower Pectin in Functional Foods. Food Process. Technol. 2016, 43, 90–95. [Google Scholar]
- Peng, X.; Yang, G.; Shi, Y.; Zhou, Y.; Zhang, M.; Li, S. Box-Behnken design based statistical modeling for the extraction and physicochemical properties of pectin from sunflower heads and the comparison with commercial low-methoxyl pectin. Sci. Rep. 2020, 10, 3595. [Google Scholar] [CrossRef]
- Li, L.; Teng, M.; Liu, Y.; Qu, Y.; Zhang, Y.; Lin, F.; Wang, D. Anti-Gouty Arthritis and Antihyperuricemia Effects of Sunflower (Helianthus annuus) Head Extract in Gouty and Hyperuricemia Animal Models. Biomed Res. Int. 2017, 2017, 5852076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Chen, X.F.; Lu, X.; Zhao, J.Y.; Li, X.L. Sunflower head enzymatic hydrolysate relives hyperuricemia by inhibiting crucial proteins (xanthine oxidase, adenosine deaminase, uric acid transporter1) and restoring gut microbiota in mice. J. Funct. Foods 2020, 72. [Google Scholar] [CrossRef]
- Lee, P.C. Developmental changes of adenosine deaminase, xanthine oxidase, and uricase in mouse tissues. Dev. Biol. 1973, 31, 227–233. [Google Scholar] [CrossRef]
- Lu, J.; He, Y.; Cui, L.; Xing, X.; Liu, Z.; Li, X.; Zhang, H.; Li, H.; Sun, W.; Ji, A.; et al. Hyperuricemia predisposes to the onset of diabetes via promoting pancreatic β-cell death in uricase-deficient Male mice. Diabetes 2020, 69, 1149–1163. [Google Scholar] [CrossRef]
- Zhou, H.; Zhong, G.; Bai, J.; Li, X.; Peng, W.; Zhang, L.; Li, J. Development of a fluorescence-based assay for screening of urate transporter 1 inhibitors using 6-carboxyfluorescein. Anal. Biochem. 2021, 626, 114246. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, Z.; Li, Y.; Li, L.; Jiang, Y.; Cao, Y.; Zhou, P.; Wu, T.; Pang, J. Characterizations of the Urate Transporter, GLUT9, and Its Potent Inhibitors by Patch-Clamp Technique. SLAS Discov. 2021, 26, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Lu, Y.; Hou, J.; Liu, C.; Sun, Y. A Polysaccharide Purified from Morchella conica Pers. Prevents Oxidative Stress Induced by H2O2 in Human Embryonic Kidney (HEK) 293T Cells. Int. J. Mol. Sci. 2018, 19, 4027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.P.; Kan, R.T.; Wu, S.J.; Guo, H.; Zhao, W.Z.; Ding, L.; Zheng, F.P.; Liu, J.B. Xanthine oxidase inhibitory peptides derived from tuna protein: Virtual screening, inhibitory activity, and molecular mechanisms. J. Sci. Food Agric. 2021, 101, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Li, R.a.; Song, W.a.; Qiao, X.a.; Liu, J.b.; Liang, H.a.; Ye, M.a. Chemical profiling of Scutellaria barbata by ultra high performance liquid chromatography coupled with hybrid quadrupoleorbitrap mass spectrometry(Article). J. Chin. Pharm. Sci. 2015, 24, 635–646. [Google Scholar] [CrossRef]
- Sriramulu, D.K.; Lee, S.G. Effect of molecular properties of the protein-ligand complex on the prediction accuracy of AutoDock. J. Mol. Graph. Model. 2021, 106, 107921. [Google Scholar] [CrossRef]
- Cao, H.; Pauff, J.; Hille, R. Substrate orientation and the origin of catalytic power in xanthine oxidoreductase. Indian J. Chem. Sect. A Inorg. Phys. Theor. Anal. 2011, 50, 355–362. [Google Scholar]
- Qi, D.; Liu, J.; Wang, C.; Wang, L.; Zhang, X.; Lin, Q.; Tu, J.; Wang, J.; Ning, X.; Cui, J. Sex-specific differences in the prevalence of and risk factors for hyperuricemia among a low-income population in China: A cross-sectional study. Postgrad. Med. 2020, 132, 559–567. [Google Scholar] [CrossRef]
- Chou, H.W.; Chiu, H.T.; Tsai, C.W.; Ting, I.W.; Yeh, H.C.; Huang, H.C.; Kuo, C.C.; Group, C.K.R. Comparative effectiveness of allopurinol, febuxostat and benzbromarone on renal function in chronic kidney disease patients with hyperuricemia: A 13-year inception cohort study. Nephrol. Dial. Transplant. 2018, 33, 1620–1627. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, J. Scopolin, a glycoside form of the phytoalexin scopoletin, is likely involved in the resistance of Nicotiana attenuata against Alternaria alternata. J. Plant Pathol. 2016, 98, 641–644. [Google Scholar] [CrossRef]
- Firmansyah, A.; Winingsih, W.; Manobi, J.D.Y. Review of Scopoletin: Isolation, Analysis Process, and Pharmacological Activity. Biointerface Res. Appl. Chem. 2021, 11, 12006–12019. [Google Scholar] [CrossRef]
- Ding, Z.; Dai, Y.; Wang, Z. Hypouricemic action of scopoletin arising from xanthine oxidase inhibition and uricosuric activity. Planta Med. 2005, 71, 183–185. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Ma, Y.; Yang, Z.; Mao, J.; Zheng, Y. Antihyperuricemic efficacy of Scopoletin-loaded Soluplus micelles in yeast extract/potassium oxonate-induced hyperuricemic mice. Drug Dev. Ind. Pharm. 2020, 46, 1550–1557. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhao, J.; Deng, W.; Li, K.; Liu, H. Effects of chlorogenic acid-enriched extract from Eucommia ulmoides Oliver leaf on growth performance and quality and oxidative status of meat in finishing pigs fed diets containing fresh or oxidized corn oil. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1116–1125. [Google Scholar] [CrossRef]
- Farah, A.; Monteiro, M.; Donangelo, C.M.; Lafay, S. Chlorogenic acids from green coffee extract are highly bioavailable in humans. J. Nutr. 2008, 138, 2309–2315. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Dong, B.; Yuan, X.; Kuang, Q.; Zhao, Q.; Yang, M.; Liu, J.; Zhao, B. Enrichment and separation of chlorogenic acid from the extract of Eupatorium adenophorum Spreng by macroporous resin. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1008, 58–64. [Google Scholar] [CrossRef]
- Ferraz-Filha, Z.S.; Ferrari, F.C.; Araújo, M.C.d.P.M.; Bernardes, A.C.F.P.F.; Saúde-Guimarães, D.A. Effects of the Aqueous Extract from Tabebuia roseoalba and Phenolic Acids on Hyperuricemia and Inflammation. Evid.-Based Complement. Alternat. Med. 2017, 2017, 2712108–2712110. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Zhang, B.; Zhao, X.; Lin, Y.; Wang, J.; Wang, X.; Hu, N.; Wang, S. Chlorogenic acid supplementation ameliorates hyperuricemia, relieves renal inflammation, and modulates intestinal homeostasis. Food Funct. 2021. [Google Scholar] [CrossRef]
- Meng, Z.-Q.; Tang, Z.-H.; Yan, Y.-X.; Guo, C.-R.; Cao, L.; Ding, G.; Huang, W.-Z.; Wang, Z.-Z.; Wang, K.D.G.; Xiao, W.; et al. Study on the anti-gout activity of chlorogenic acid: Improvement on hyperuricemia and gouty inflammation. Am. J. Chin. Med. 2014, 42, 1471–1483. [Google Scholar] [CrossRef]
- Dhiman, P.; Malik, N.; Khatkar, A.; Kulharia, M. Antioxidant, xanthine oxidase and monoamine oxidase inhibitory potential of coumarins: A review. Curr. Org. Chem. 2017, 21, 294–304. [Google Scholar] [CrossRef]
- Song, D.; Zhao, X.; Wang, F.; Wang, G. A brief review of urate transporter 1 (URAT1) inhibitors for the treatment of hyperuricemia and gout: Current therapeutic options and potential applications. Eur. J. Pharmacol. 2021, 907, 174291. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.J.; Takeda, M.; Enomoto, A.; Fujimura, M.; Miyazaki, H.; Anzai, N.; Endou, H. Interactions of urate transporter URAT1 in human kidney with uricosuric drugs. Nephrology 2011, 16, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Claverie-Martin, F.; Trujillo-Suarez, J.; Gonzalez-Acosta, H.; Aparicio, C.; Justa Roldan, M.L.; Stiburkova, B.; Ichida, K.; Martín-Gomez, M.A.; Herrero Goñi, M.; Carrasco Hidalgo-Barquero, M.; et al. URAT1 and GLUT9 mutations in Spanish patients with renal hypouricemia. Clin. Chim. Acta 2018, 481, 83–89. [Google Scholar] [CrossRef]
- Cao, H.; Hall, J.; Hille, R. Substrate Orientation and Specificity in Xanthine Oxidase: Crystal Structures of the Enzyme in Complex with Indole-3-acetaldehyde and Guanine. Biochemistry 2014, 53, 533–541. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, H.; Lv, S.; Fu, X.; Li, W. Identification of Scopoletin and Chlorogenic Acid as Potential Active Components in Sunflower Calathide Enzymatically Hydrolyzed Extract towards Hyperuricemia. Appl. Sci. 2021, 11, 10306. https://doi.org/10.3390/app112110306
Dai H, Lv S, Fu X, Li W. Identification of Scopoletin and Chlorogenic Acid as Potential Active Components in Sunflower Calathide Enzymatically Hydrolyzed Extract towards Hyperuricemia. Applied Sciences. 2021; 11(21):10306. https://doi.org/10.3390/app112110306
Chicago/Turabian StyleDai, Huining, Shuai Lv, Xueqi Fu, and Wannan Li. 2021. "Identification of Scopoletin and Chlorogenic Acid as Potential Active Components in Sunflower Calathide Enzymatically Hydrolyzed Extract towards Hyperuricemia" Applied Sciences 11, no. 21: 10306. https://doi.org/10.3390/app112110306
APA StyleDai, H., Lv, S., Fu, X., & Li, W. (2021). Identification of Scopoletin and Chlorogenic Acid as Potential Active Components in Sunflower Calathide Enzymatically Hydrolyzed Extract towards Hyperuricemia. Applied Sciences, 11(21), 10306. https://doi.org/10.3390/app112110306