
The Nasal Epithelium as a Route of Infection and Clinical Signs Changes, in Rainbow Trout (Oncorhynchus mykiss) Fingerlings Infected with Aeromonas spp

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Husbandry
2.3. Behavioral Observations and Clinical Signs
2.4. Histopathological Studies
2.5. Statistical Analysis
3. Results
3.1. Behaviors and Clinical Signs
3.2. Histopathological Lesions in Nasal and Nervous Tissue
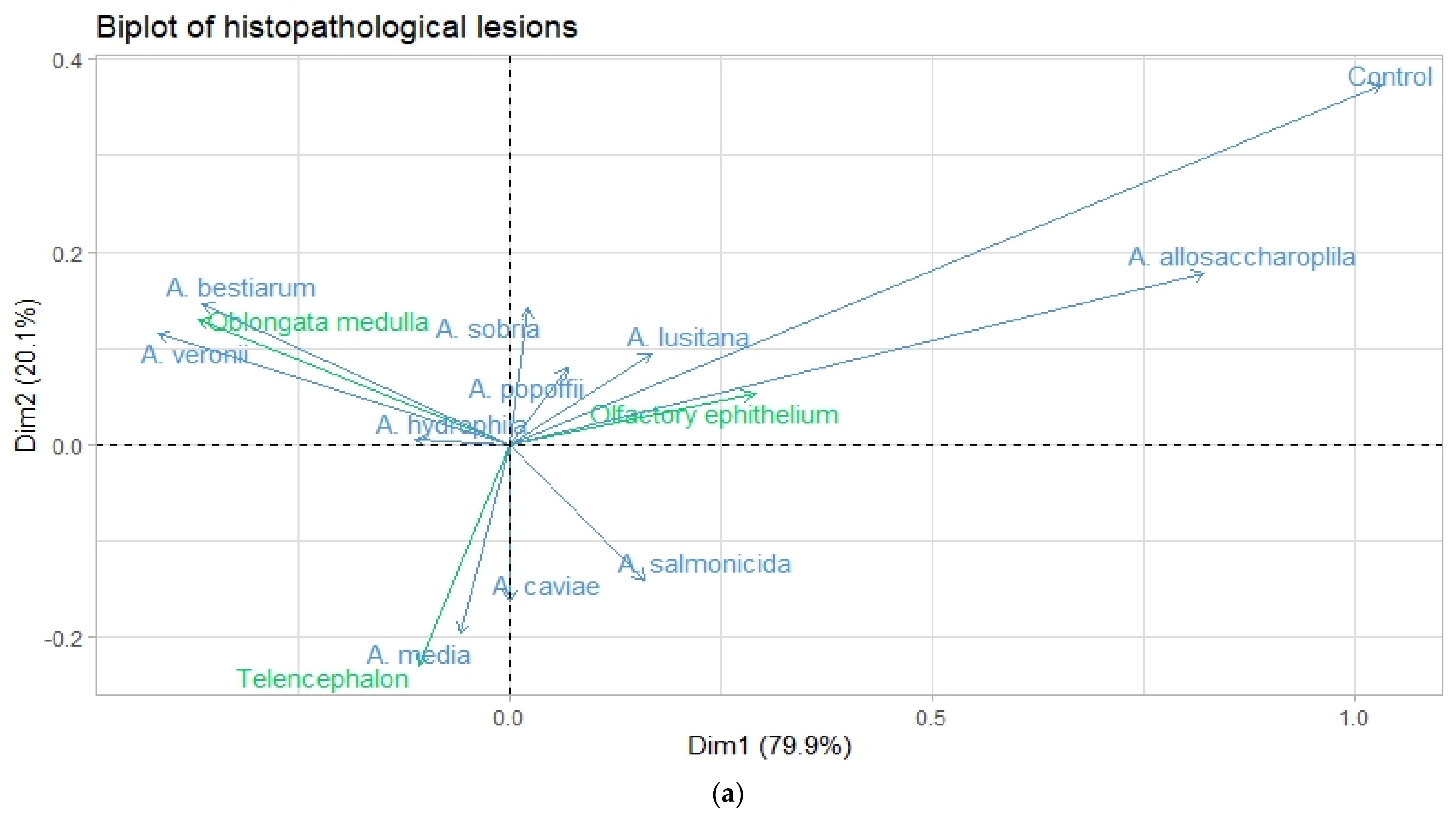
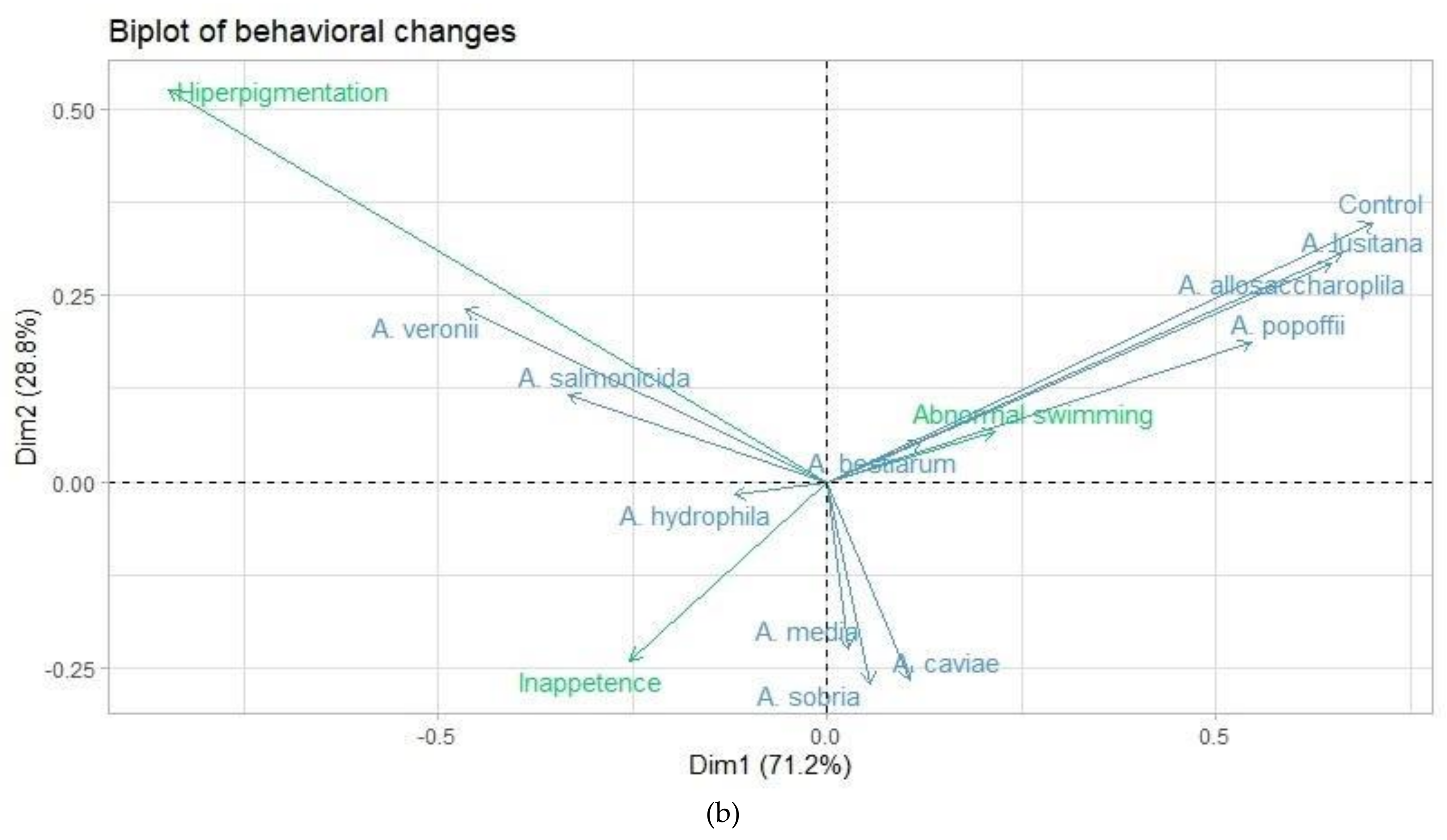
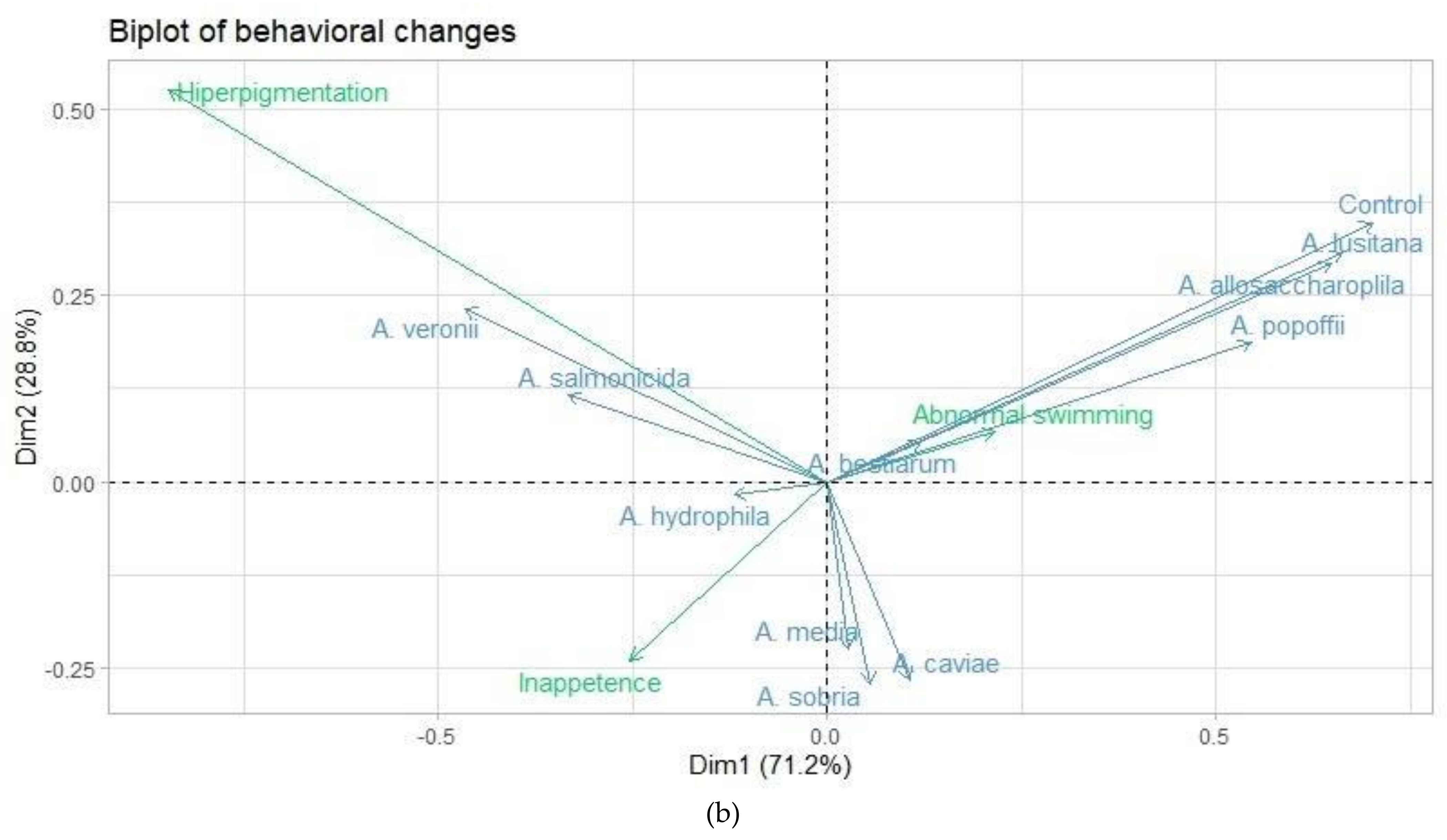
3.3. Correspondence Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, B.; Mao, C.; Feng, J.; Li, Y.; Hu, J.; Jiang, B.; Gu, Q.; Su, Y. A First report of Aeromonas veronii infection of the sea bass, Lateolabrax maculatus in China. Front. Veter-Sci. 2021, 7, 600587. [Google Scholar] [CrossRef]
- Srivastava, A.; Mistri, A.; Mittal, S.; Kumar, A.M. Alterations in the epidermis of the carp, Labeo rohita (Cyprinidae: Cy-priniformes), infected by the bacteria, Aeromonas hydrophila: A scanning electron microscopic, histopathological and im-munohistochemical investigation. J. Fish Dis. 2020, 43, 941–953. [Google Scholar] [CrossRef]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, I.; Spanggaard, B.; Appel, K.F.; Rossen, L.; Nielsen, T.; Gram, L. Phylogenetic analysis in situ identification of the in-testinal microbial community of rainbow trout (Oncorhynchus mykiss, Walbaum). J. Appl. Microbiol. 2004, 96, 117–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, B.; Austin, D.A. Bacterial Fish Pathogens, Diseases of Farmed and Wild Fish, 4th ed.; Praxis Publishing Ltd.: Chichester, UK, 2007; pp. 123–153. [Google Scholar]
- Fernández-Bravo, A.; Figueras, M.J. An update on the genus Aeromonas: Taxonomy, epidemiology, and pathogenicity. Microorganisms 2020, 8, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallaire-Dufresne, S.; Tanaka, K.H.; Trudel, M.V.; Lafaille, A.; Charette, S.J. Virulence, genomic features, and plasticity of Aeromonas salmonicida subsp. salmonicida, the causative agent of fish furunculosis. Veter-Microbiol. 2014, 169, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zepeda-Velázquez, A.P.; Sánchez, V.V.; Ortega-Santana, C.; Rubio-Godoy, M.; de Oca-Mira, D.M.; Soriano-Vargas, E. Pathogenicity of Mexican isolates of Aeromonas sp. in immersion experimentally-infected rainbow trout (Oncorhynchus mykiss, Walbaum 1792). Acta Trop. 2017, 169, 122–124. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, R.J.M.; Pérez, H.J.A. Aislamiento de Aeromonas hydrophila en truchas Arcoíris (Oncorhynchus mykiss). Vet. Méx. 1998, 29, 117–119. [Google Scholar]
- Zepeda-Velázquez, A.P.; Vega-Sánchez, V.; Salgado-Miranda, C.; Soriano-Vargas, E. Histopathological findings in farmed rainbow trout (Oncorhynchus mykiss) naturally infected with 3 different Aeromonas species. Can. J. Vet. Res. 2015, 79, 250–254. [Google Scholar]
- Marinho-Neto, F.A.; Claudiano, G.S.; Aguinaga, J.Y.; Quiroz, V.A.C.; Kobashigawa, K.K.; Cruz, N.R.N.; Moraes, F.R.; Moraes, J.R.E. Morphological, microbiological and ultrastructural aspects of sepsis by Aeromonas hydrophila in Piaractus mesopotamicus. PLoS ONE 2019, 14, e0222626. [Google Scholar] [CrossRef] [PubMed]
- Noga, E.J. Fish Disease, Diagnosis, and Treatment, 2nd ed.; Wiley-Blackwell: Iowa, IA, USA, 2010. [Google Scholar]
- LaPatra, S.; Kao, S.; Erhardt, E.B.; Salinas, I. Evaluation of dual nasal delivery of infectious hematopoietic necrosis virus and enteric red mouth vaccines in rainbow trout (Oncorhynchus mykiss). Vaccine 2015, 33, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Salinas, I.; LaPatra, S.; Erhardt, E. Nasal vaccination of young rainbow trout (Oncorhynchus mykiss) against infectious hematopoietic necrosis and enteric red mouth disease. Dev. Comp. Immunol. 2015, 53, 105–111. [Google Scholar] [CrossRef]
- Bartkova, S.; Kokotovic, B.; Dalsgaard, I. Infection routes of Aeromonas salmonicida in rainbow trout monitored in vivo by real-time bioluminescence imaging. J. Fish Dis. 2017, 40, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Strøm, H.K.; Ohtani, M.; Nowak, B.; Boutrup, T.S.; Jones, B.; Raida, M.K.; Bojesen, A.M. Experimental infection by Yersinia ruckeri O1 biotype 2 induces brain lesions and neurological signs in rainbow trout (Oncorhynchus mykiss). J. Fish Dis. 2017, 41, 529–537. [Google Scholar] [CrossRef]
- Farto, R.; Milton, D.L.; Bermúdez, M.B.; Nieto, T.P. Colonization of turbot tissues by virulent and avirulent Aeromonas salm-onicida subsp salmonicida strains during infection. Dis. Aquat. Organ. 2011, 95, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Nordgreen, J.; Tahamtani, F.M.; Janczak, A.M.; Horsberg, T.E. Behavioural effects of the commonly used fish anaesthetic tricaine methanesulfonate (MS-222) on zebrafish (Danio rerio) and its relevance for the acetic acid pain test. PLoS ONE 2014, 9, e92116. [Google Scholar] [CrossRef]
- Aydin, S.; Çiltaş, A. Systemic infections of Aeromonas hydrophila in rainbow trout (Oncorhynchus mykiss Walbaum): Gross pathology, bacteriology, clinical pathology, histopathology, and chemotherapy. J. Anim. Vet. Adv. 2004, 3, 810–819. [Google Scholar]
- Aydoğan, A.; Metin, N. Investigation of pathological findings infected with Aeromonas salmonicida in rainbow trout (Oncorhynchus mykiss Walbaum, 1972). Kafkas Univ. Vet. Fak Derg 2010, 16, S325–S328. [Google Scholar]
- Inglis, V.; Roberts, R.J.; Bromage, N.R. Bacterial Diseases of Fish, 1st ed.; Iowa State University Press: Iowa City, IA, USA, 1993; pp. 80–121. [Google Scholar]
- Lago, E.P.; Nieto, T.P.; Farto, R. Virulence factors of Aeromonas salmonicida subsp. salmonicida strains associated with infections in turbot Psetta maxima. Dis. Aquat. Org. 2012, 99, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, R.J. Fish Pathology, 3rd ed.; Wiley-Blackwell: Iowa, IA, USA, 2001. [Google Scholar]
- Miller, K.M.; Günther, P.O.; Li, S.; Kaukinen, H.K.; Ming, J.T. Molecular indices of viral disease development in wild migrating salmon. Conserv Physiol. 2017, 5, cox036. [Google Scholar] [CrossRef] [PubMed]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Kassambara, A.; Mundt, F. Factoextra: Extract and visualize the results of multivariate data analyses. R Package Version 2017, 1, 337–354. [Google Scholar]
- Speare, D. Differences in patterns of meningoencephalitis due to bacterial kidney disease in farmed Atlantic and chinook salmon. Res. Veter- Sci. 1997, 62, 79–80. [Google Scholar] [CrossRef]
- Hamdani, E.H.; Døving, K.B. The functional organization of the fish olfactory system. Prog. Neurobiol. 2007, 82, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Xu, S.; Hu, Q.; Hu, M.; Zhou, L.; Qin, X.; Jia, J.; Hu, G. Structure and function analysis of various brain subregions and pituitary in grass carp (Ctenopharyngodon idellus). Comp. Biochem. Physiol. Part D Genom. Proteom. 2020, 33, 100653. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhao, J.; Bo, Y.; Liu, Z.; Wu, K.; Gong, C. Aeromonas hydrophila induces intestinal inflammation in grass carp (Cte-nopharyngodon idella): An experimental model. Aquaculture 2014, 434, 171–178. [Google Scholar] [CrossRef]
- Bøgwald, J.; Dalmo, R.A. Review on Immersion Vaccines for Fish: An Update. Microorganisms 2019, 7, 627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Wang, X.; Liu, Y.; Fu, Q.; Tian, C.; Wu, C.; Shi, H.; Yuan, Z.; Tan, S.; Liu, S.; et al. Transcriptome analysis reveals enrichment of genes associated with auditory system in swim bladder of channel catfish. Comp. Biochem. Physiol. Part D Genom. Proteom. 2018, 27, 30–39. [Google Scholar]
- Kostyniuk, D.J.; Culbert, B.M.; Mennigen, J.A.; Gilmour, K.M. Social status affects lipid metabolism in rainbow trout, Oncorhynchus mykiss. Am. J. Physiol. Integr. Comp. Physiol. 2018, 315, R241–R255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vissio, P.G.; Darias, M.J.; Di Yorio, M.P.; Sirkin, D.I.P.; Delgadin, T.H. Fish skin pigmentation in aquaculture: The influence of rearing conditions and its neuroendocrine regulation. Gen. Comp. Endocrinol. 2021, 301, 113662. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.T.; Techatanakitarnan, C.; Jindakittikul, P.; Thaiprayoon, A.; Taengphu, S.; Charoensapsri, W.; Khunrae, P.; Rattanarojpong, T.; Senapin, S. Aeromonas jandaei and Aeromonas veronii caused disease and mortality in Nile tilapia, Oreochromis niloticus (L.). J. Fish Dis. 2017, 40, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Sreedharan, K.; Philip, R.; Sarojani, I.; Singh, B. Characterization, and virulence potential of phenotypically diverse Aeromonas veronii isolates recovered from moribund freshwater ornamental fishes of Kerala, India. Antonie Van Leeuwenhoek 2013, 103, 53–67. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavior or Clinical Sign | Definition | Reference(s) |
|---|---|---|
| Loss of appetite | Fish refuse to eat and/or lose interest in food (anorexia) | [12,17,19,20,21] |
| Abnormal swimming (Erratic swimming) | Loss of consistency, regularity and/or uniformity in movement; fish swimming in different directions | [17,22] |
| Hyperpigmentation and/or darkened skin | Fingerlings show an excess of melanin pigmentation in the skin, giving them a darker color | [1,9,12,23] |
| Aeromonas Species | N | % of Fish with Clinical Sign or Behavioral Changes 1 | ||
|---|---|---|---|---|
| Darkened Skin/Hyperpigmentation | Lack of Appetite/Inappetence | Abnormal Swimming | ||
| A. allosaccharophila | 90 | 0 | 1.11 | 25.55 |
| A. bestiarum | 90 | 6.66 | 32.22 | 83.33 |
| A. caviae | 90 | 0 | 38.88 | 60 |
| A. hydrophila | 90 | 11.11 | 46.66 | 74.44 |
| A. lusitana | 90 | 0 | 1.11 | 34.44 |
| A. media | 90 | 1.11 | 16.66 | 25.55 |
| A. popoffii | 90 | 0 | 3.33 | 31.11 |
| A. salmonicida | 90 | 18.88 | 47.7 | 66.66 |
| A. sobria | 90 | 1.11 | 46.66 | 67.77 |
| A. veronii | 90 | 15.55 | 28.88 | 38.88 |
| Control group | 90 | 0 | 0 | 8.88 |
| Aeromonas Species | N | % of Fish with Histopathological Lesions 1 | ||
|---|---|---|---|---|
| Rosette | Telencephalon | Oblongata Medulla | ||
| A. allosaccharophila | 90 | 30 | 3.3 | 1.1 |
| A. bestiarum | 90 | 26.6 | 21.1 | 40 |
| A. caviae | 90 | 28.8 | 22.2 | 16.6 |
| A. hydrophila | 90 | 27.7 | 18.8 | 22.2 |
| A. lusitana | 90 | 50 | 18.8 | 22.2 |
| A. media | 90 | 35.5 | 31.1 | 22.2 |
| A. popoffii | 90 | 7.7 | 3.3 | 4.4 |
| A. salmonicida | 90 | 41.1 | 25.5 | 15.5 |
| A. sobria | 90 | 20 | 7.7 | 12.2 |
| A. veronii | 90 | 11.1 | 10 | 17.7 |
| Control group | 90 | 0 | 0 | 0 |
| Item | Histopathological Lesions | Behavioral Changes |
|---|---|---|
| X2 (observed value) | 68.20 | 123.64 |
| X2 (critical value) | 31.41 | 31.41 |
| DF | 20 | 20 |
| p-value | 0.0001 | 0.0001 |
| α | 0.05 | 0.05 |
| Item | Histopathological Lesions | |
|---|---|---|
| Eigen value | 0.079 | 0.020 |
| Inertia (%) | 79.94 | 20.05 |
| Cumulative (%) | 79.94 | 100.00 |
| Item | Behavioral changes | |
| Dimension 1 | Dimension 2 | |
| Eigen value | 0.095 | 0.038 |
| Inertia (%) | 71.17 | 28.82 |
| Cumulative (%) | 71.17 | 100.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez de Anda, F.R.; Vega-Sánchez, V.; Reyes-Rodríguez, N.E.; Martínez-Juárez, V.M.; Ángeles-Hernández, J.C.; Acosta-Rodríguez, I.; Campos-Montiel, R.G.; Zepeda-Velázquez, A.P. The Nasal Epithelium as a Route of Infection and Clinical Signs Changes, in Rainbow Trout (Oncorhynchus mykiss) Fingerlings Infected with Aeromonas spp. Appl. Sci. 2021, 11, 9159. https://doi.org/10.3390/app11199159
Gómez de Anda FR, Vega-Sánchez V, Reyes-Rodríguez NE, Martínez-Juárez VM, Ángeles-Hernández JC, Acosta-Rodríguez I, Campos-Montiel RG, Zepeda-Velázquez AP. The Nasal Epithelium as a Route of Infection and Clinical Signs Changes, in Rainbow Trout (Oncorhynchus mykiss) Fingerlings Infected with Aeromonas spp. Applied Sciences. 2021; 11(19):9159. https://doi.org/10.3390/app11199159
Chicago/Turabian StyleGómez de Anda, Fabián Ricardo, Vicente Vega-Sánchez, Nydia Edith Reyes-Rodríguez, Víctor Manuel Martínez-Juárez, Juan Carlos Ángeles-Hernández, Ismael Acosta-Rodríguez, Rafael German Campos-Montiel, and Andrea Paloma Zepeda-Velázquez. 2021. "The Nasal Epithelium as a Route of Infection and Clinical Signs Changes, in Rainbow Trout (Oncorhynchus mykiss) Fingerlings Infected with Aeromonas spp" Applied Sciences 11, no. 19: 9159. https://doi.org/10.3390/app11199159
APA StyleGómez de Anda, F. R., Vega-Sánchez, V., Reyes-Rodríguez, N. E., Martínez-Juárez, V. M., Ángeles-Hernández, J. C., Acosta-Rodríguez, I., Campos-Montiel, R. G., & Zepeda-Velázquez, A. P. (2021). The Nasal Epithelium as a Route of Infection and Clinical Signs Changes, in Rainbow Trout (Oncorhynchus mykiss) Fingerlings Infected with Aeromonas spp. Applied Sciences, 11(19), 9159. https://doi.org/10.3390/app11199159