
Nannochloropsis oceanica as a Sustainable Source of n-3 Polyunsaturated Fatty Acids for Enrichment of Hen Eggs

 ,
, 
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Diet Formulation and Experimental Design
2.3. Nutritional Composition and Fatty Acid Profile of N. oceanica, Hen Diets, and Eggs
2.3.1. Total Protein
2.3.2. Total Lipids
2.3.3. Ash Content
2.3.4. Fatty Acid Profile
2.4. Physical Parameters of Eggs
2.5. Statistical Analysis
3. Results and Discussion
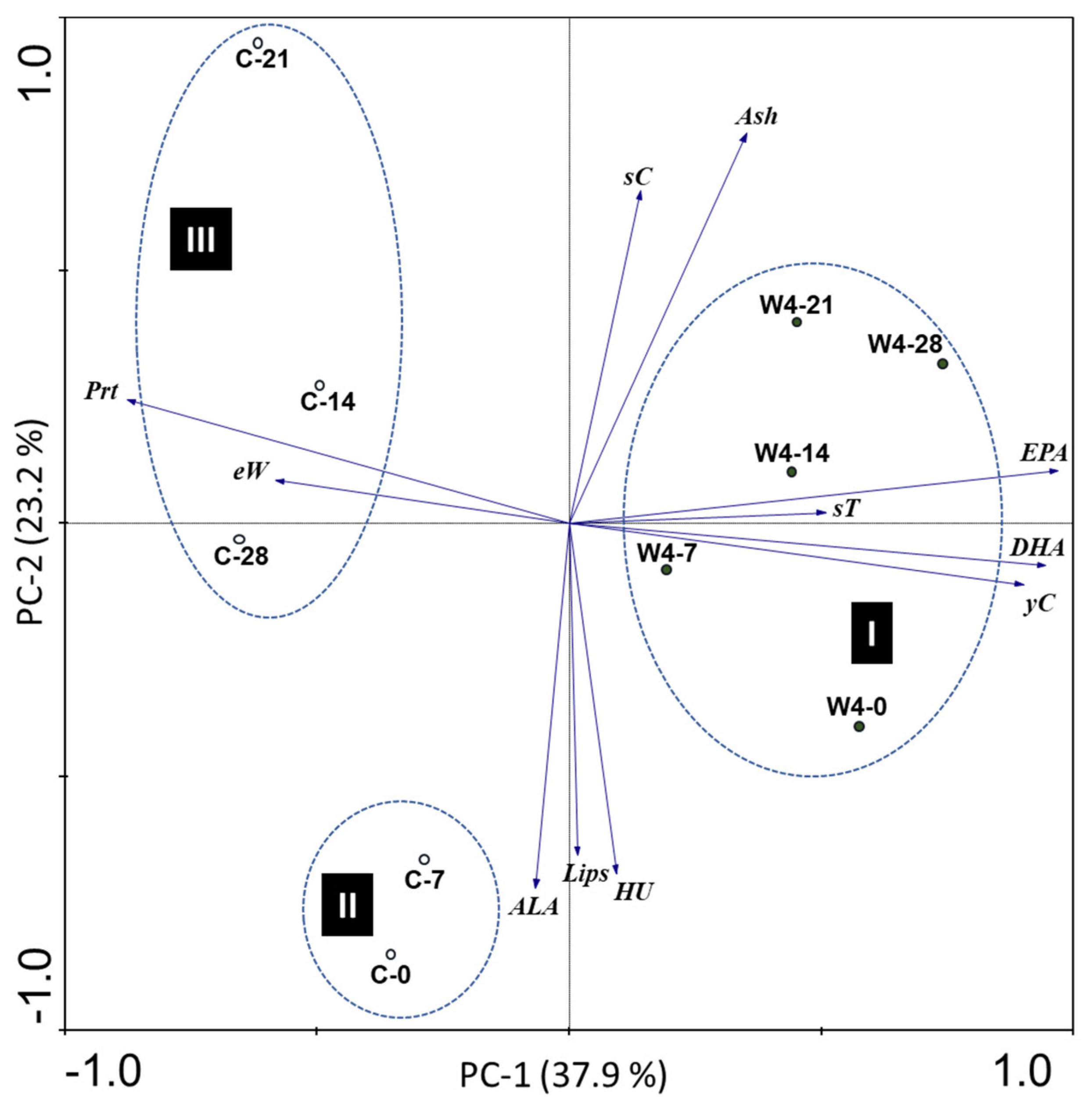
3.1. Nutritional Characterization of N. oceanica, Standard and Supplemented Diets
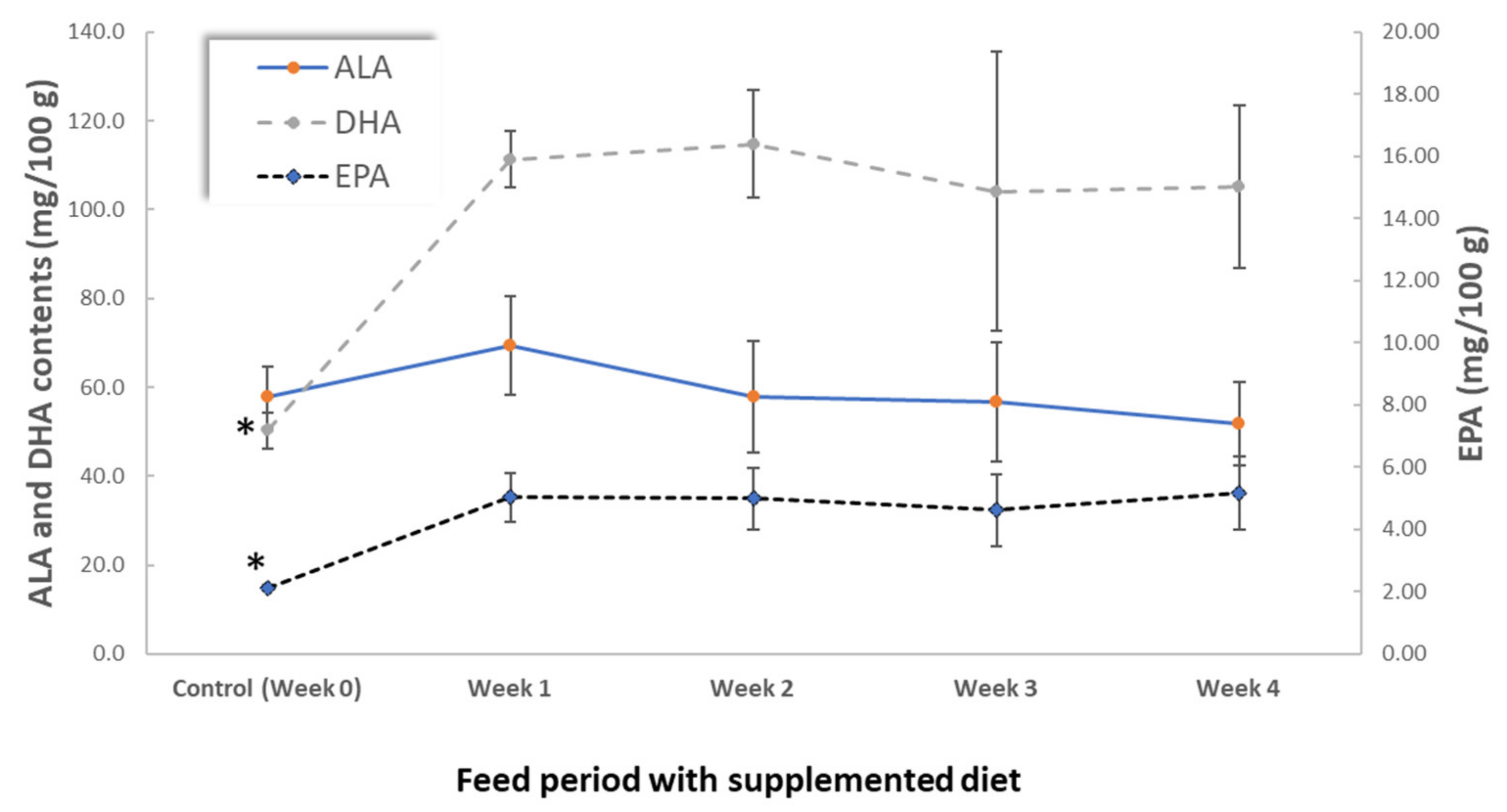
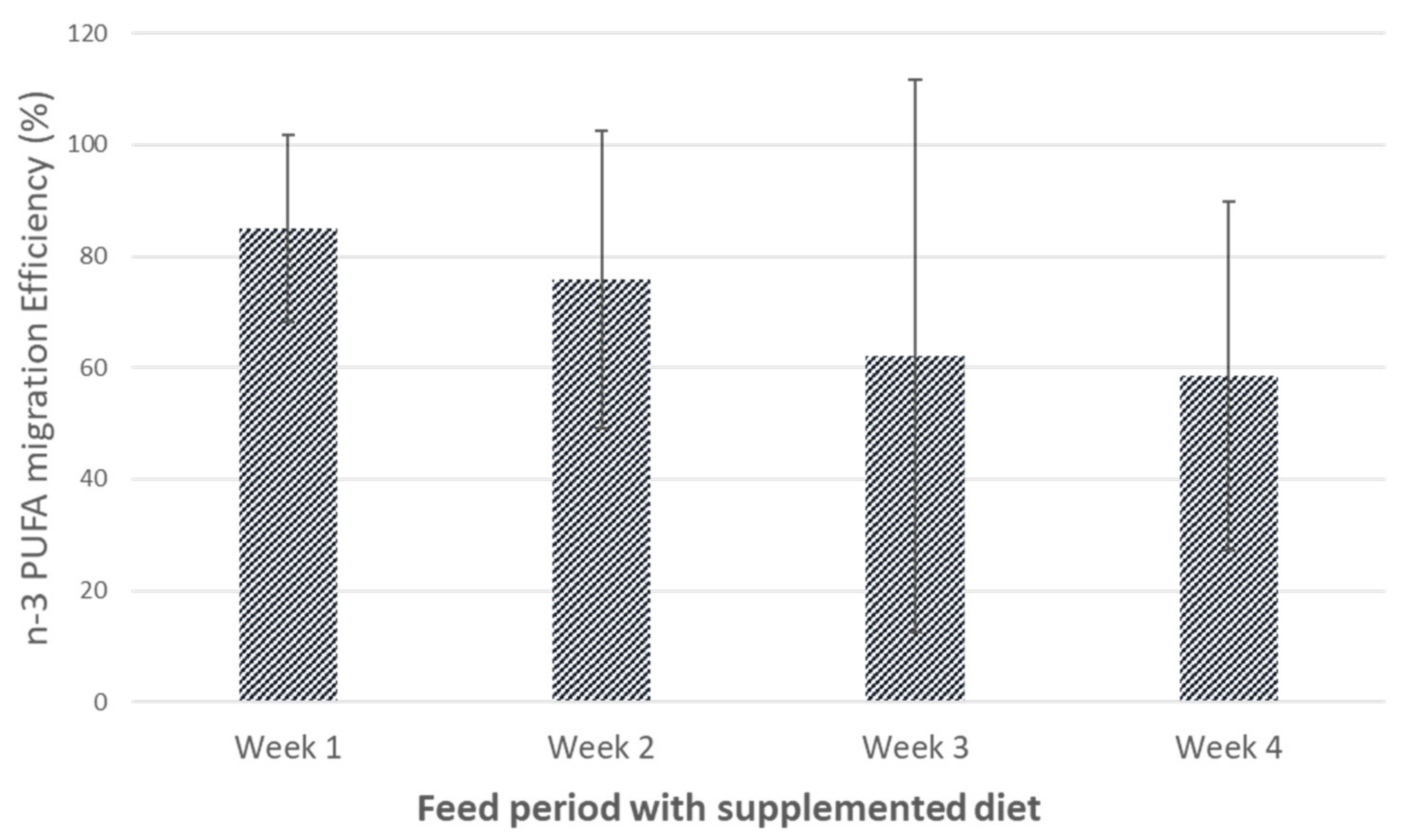
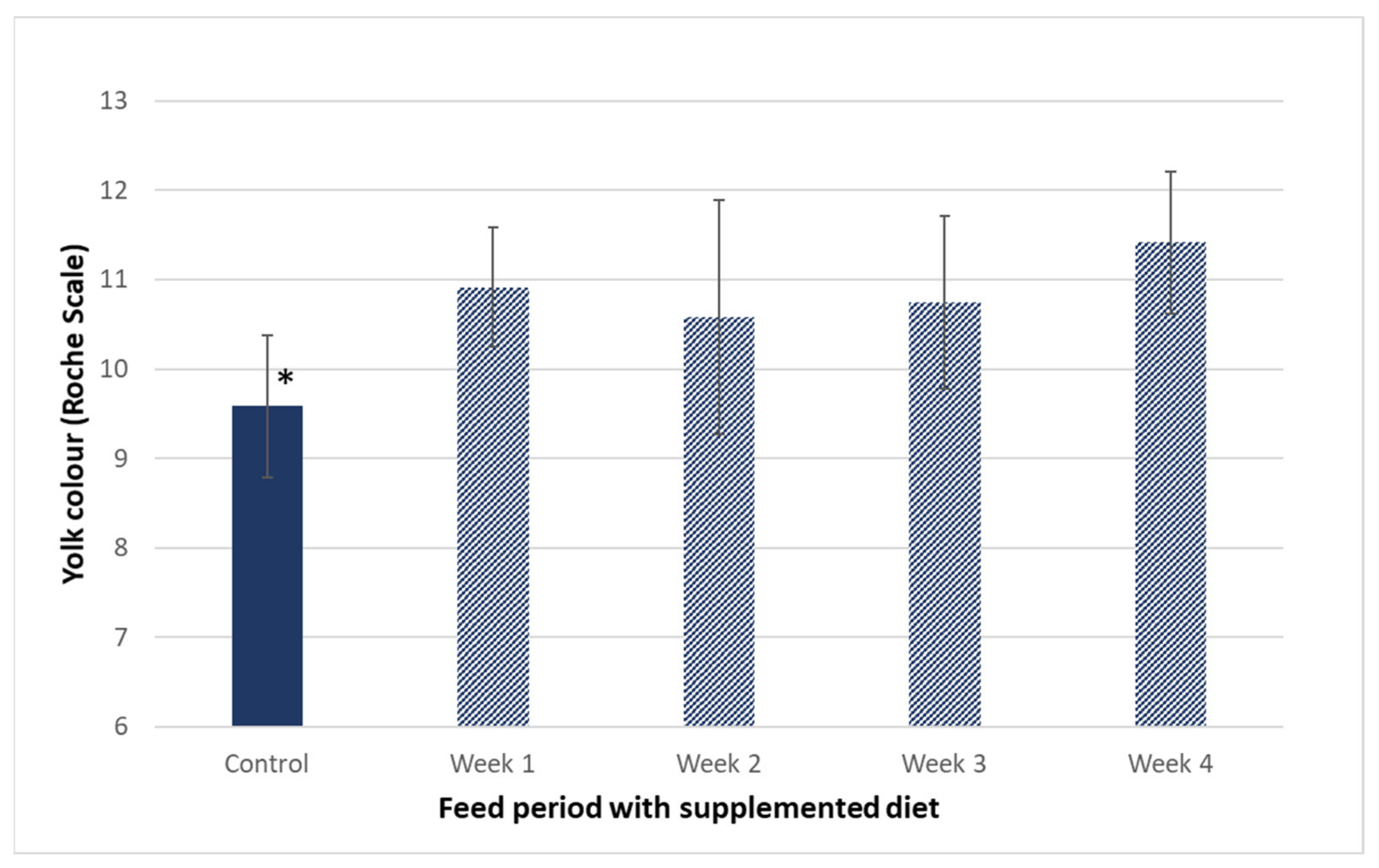
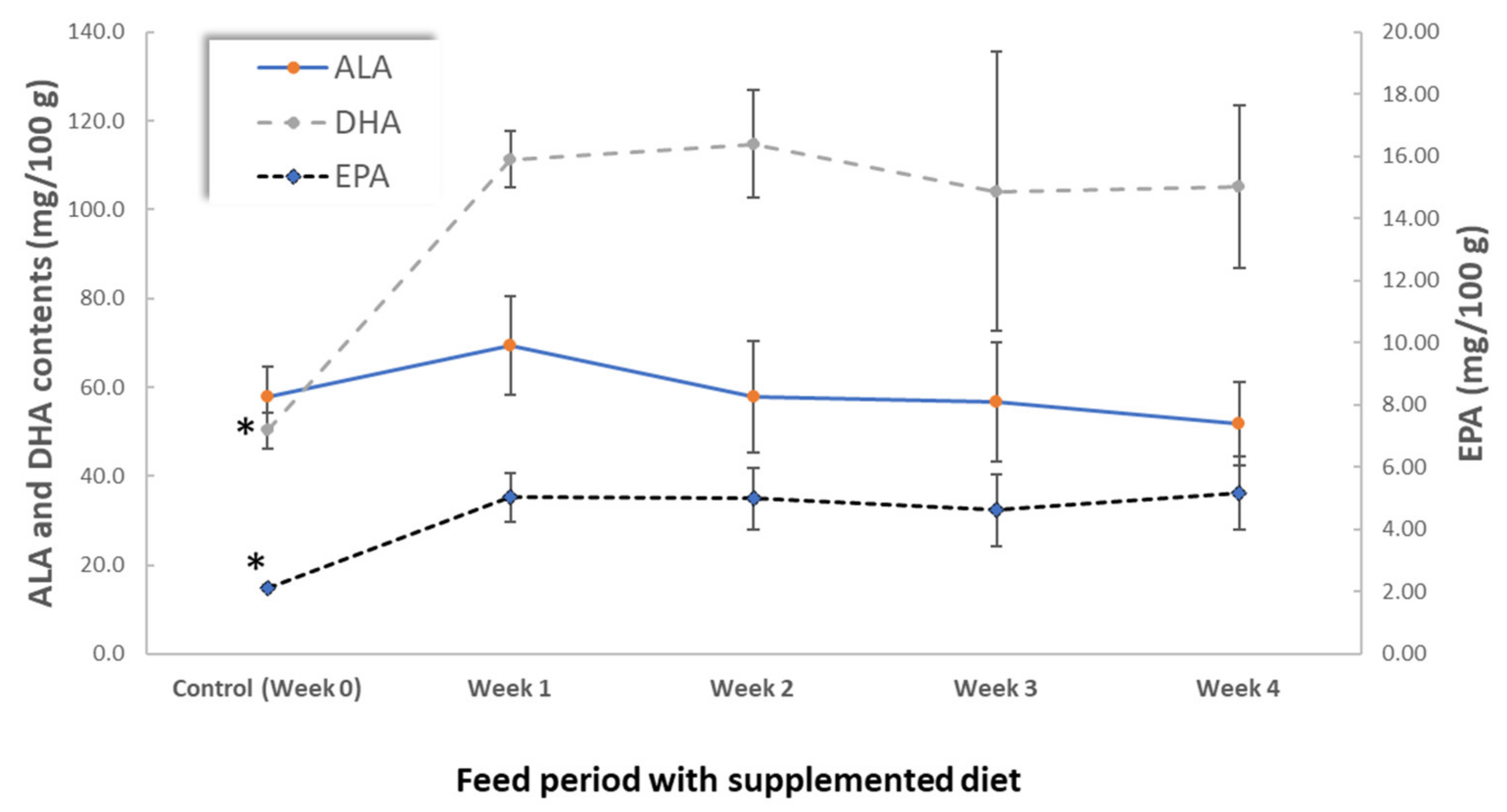
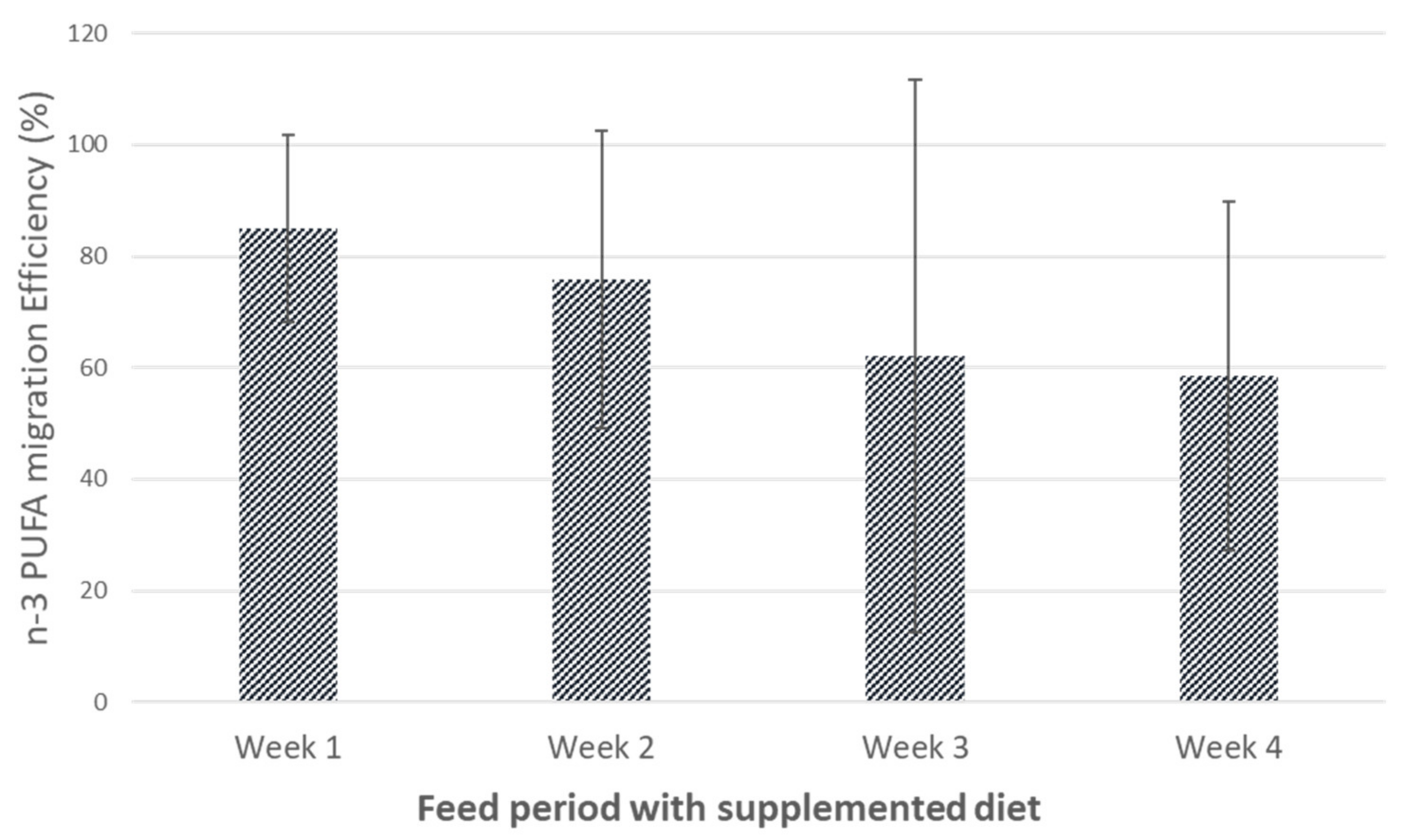
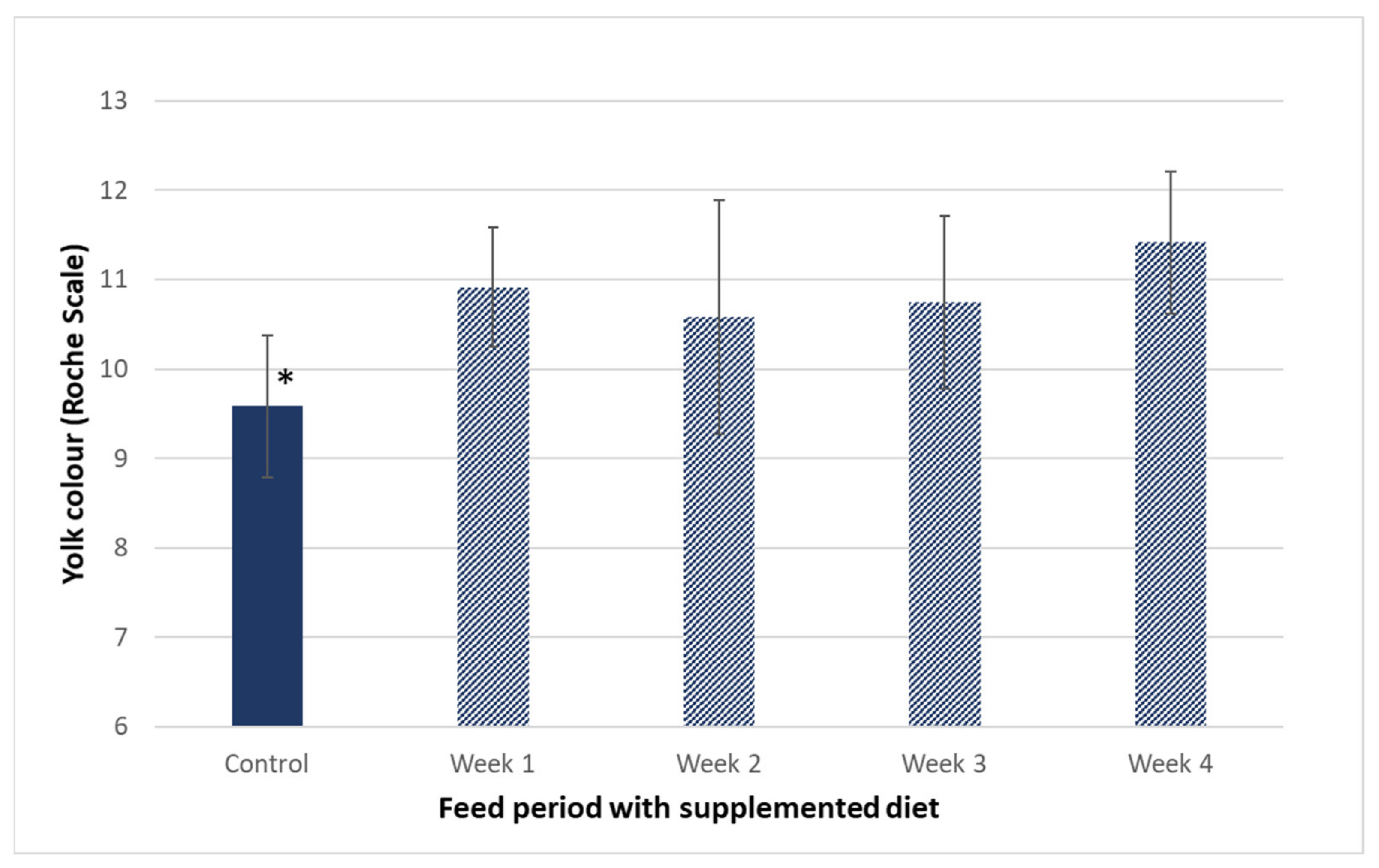
3.2. Influence of Hen Diet on Nutritional Profile and Physical Characteristics of Eggs
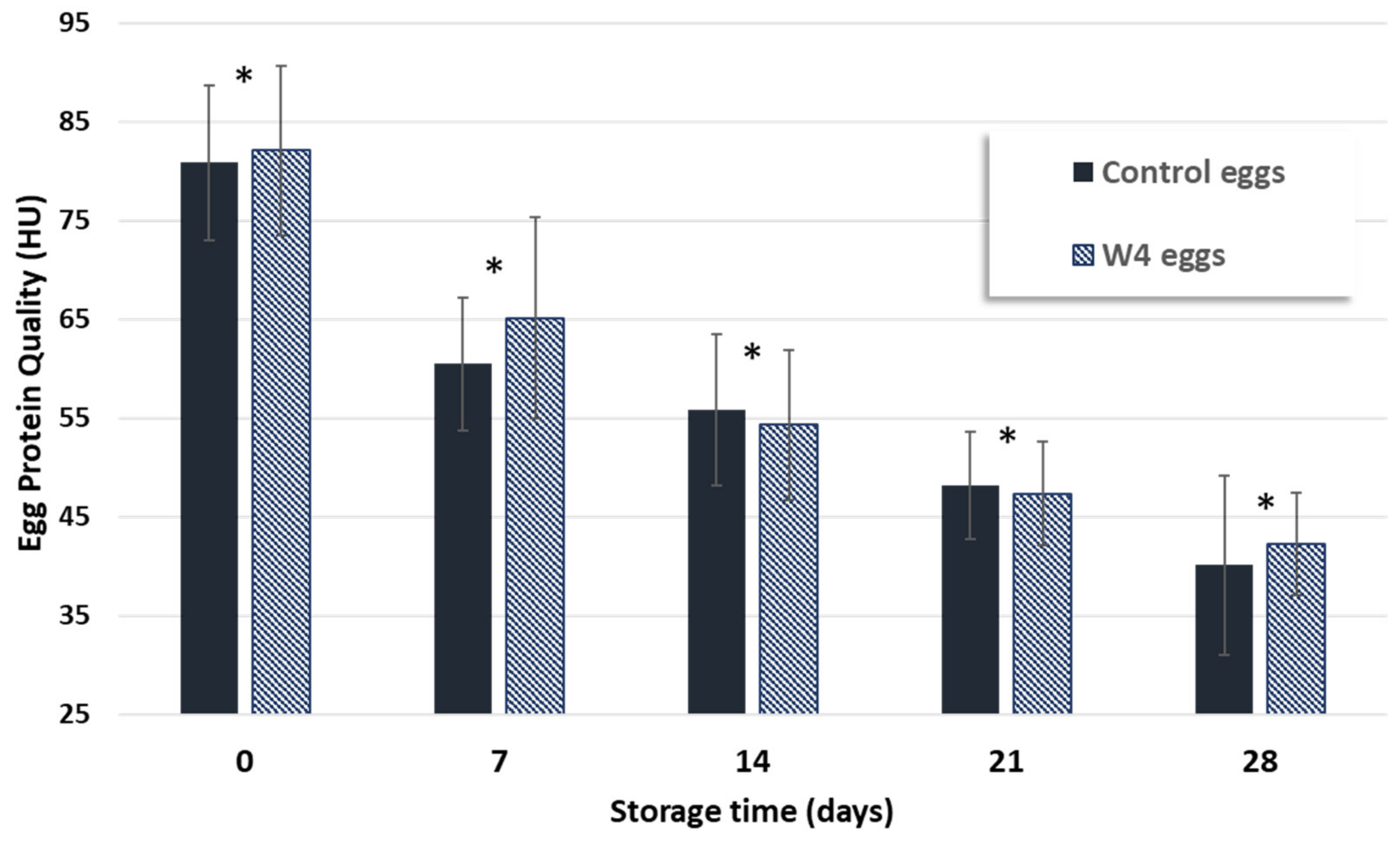
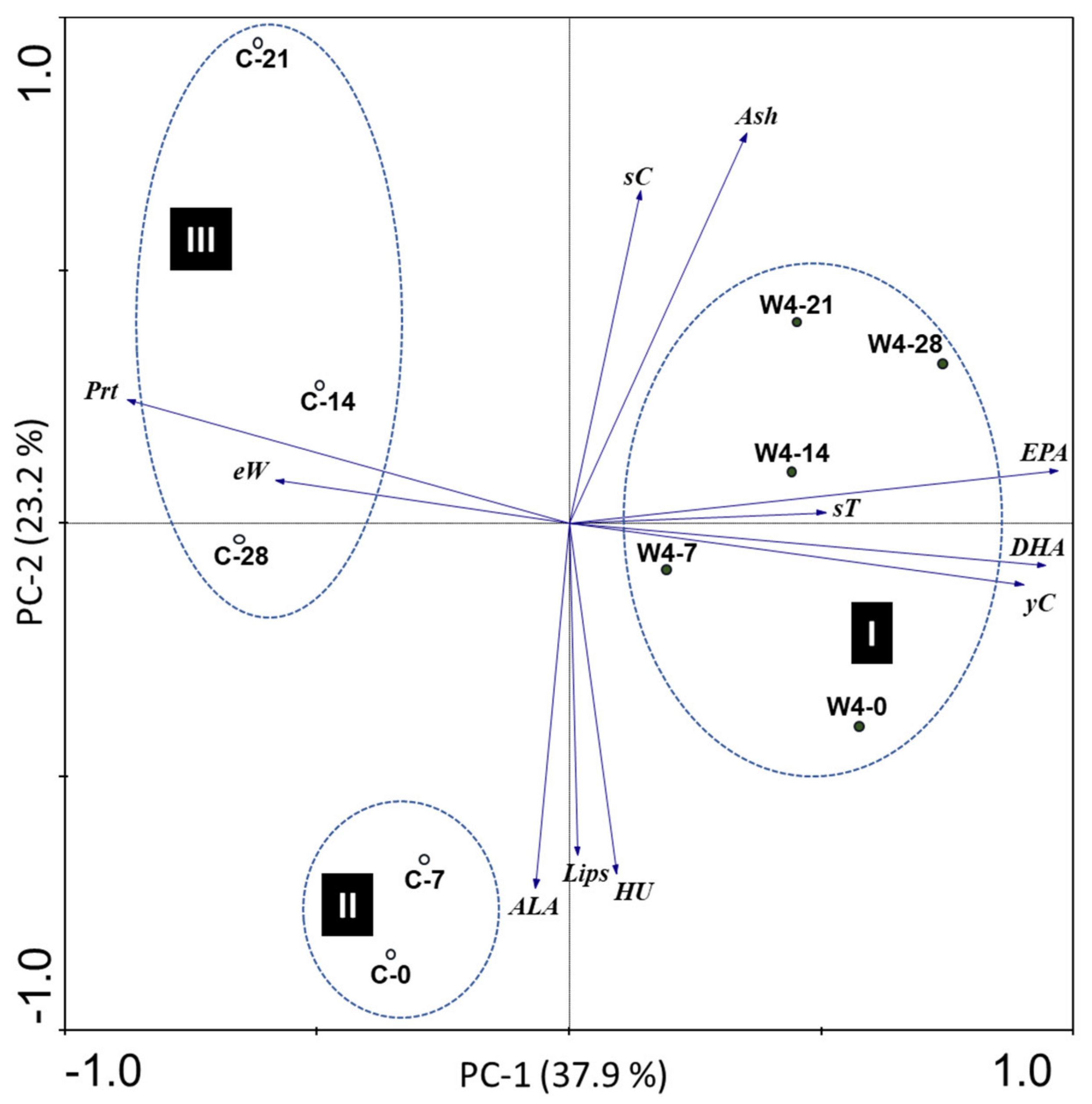
3.3. Assessment of Shelf Life of Eggs from Hens Fed with Supplemented Diet
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Watanabe, Y.; Tatsuno, I. Omega-3 Polyunsaturated Fatty Acids for Cardiovascular Diseases: Present, Past and Future. Expert Rev. Clin. Pharmacol. 2017, 10, 865–873. [Google Scholar] [CrossRef]
- Innes, J.K.; Calder, P.C. Marine Omega-3 (N-3) Fatty Acids for Cardiovascular Health: An Update for 2020. Int. J. Mol. Sci. 2020, 21, 1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, H.; Park, K. Omega-3 and Omega-6 Polyunsaturated Fatty Acids and Metabolic Syndrome: A Systematic Review and Meta-Analysis. Clin. Nutr. 2020, 39, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Albracht-Schulte, K.; Kalupahana, N.S.; Ramalingam, L.; Wang, S.; Rahman, S.M.; Robert-McComb, J.; Moustaid-Moussa, N. Omega-3 Fatty Acids in Obesity and Metabolic Syndrome: A Mechanistic Update. J. Nutr. Biochem. 2018, 58, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Nettleton, J.A.; Katz, R. N-3 Long-Chain Polyunsaturated Fatty Acids in Type 2 Diabetes: A Review. J. Am. Diet. Assoc. 2005, 105, 428–440. [Google Scholar] [CrossRef]
- Ciappolino, V.; Delvecchio, G.; Agostoni, C.; Mazzocchi, A.; Altamura, A.C.; Brambilla, P. The Role of N-3 Polyunsaturated Fatty Acids (n-3PUFAs) in Affective Disorders. J. Affect. Disord. 2017, 224, 32–47. [Google Scholar] [CrossRef] [PubMed]
- Saccà, S.; Cutolo, C.; Ferrari, D.; Corazza, P.; Traverso, C. The Eye, Oxidative Damage and Polyunsaturated Fatty Acids. Nutrients 2018, 10, 668. [Google Scholar] [CrossRef] [Green Version]
- Molendi-Coste, O.; Legry, V.; Leclercq, I.A. Why and How Meet N-3 PUFA Dietary Recommendations? Gastroenterol. Res. Pract. 2011, 2011, 364040. [Google Scholar] [CrossRef] [Green Version]
- Yashodhara, B.M.; Umakanth, S.; Pappachan, J.M.; Bhat, S.K.; Kamath, R.; Choo, B.H. Omega-3 Fatty Acids: A Comprehensive Review of Their Role in Health and Disease. Postgrad. Med. J. 2009, 85, 84–90. [Google Scholar] [CrossRef]
- Saleh, A.A.; Gawish, E.; Mahmoud, S.F.; Amber, K.; Awad, W.; Alzawqari, M.H.; Shukry, M.; Abdel-Moneim, A.M.E. Effect of Natural and Chemical Colourant Supplementation on Performance, Egg-Quality Characteristics, Yolk Fatty-Acid Profile, and Blood Constituents in Laying Hens. Sustainability 2021, 13, 4503. [Google Scholar] [CrossRef]
- Lemahieu, C.; Bruneel, C.; Muylaert, K.; Buyse, J.; Foubert, I. Microalgal Feed Supplementation to Enrich Eggs with Omega-3 Fatty Acids. In Egg Innovations and Strategies for Improvements; Academic Press: Cambridge, MA, USA, 2017; pp. 383–391. ISBN 9780128011515. [Google Scholar]
- Fraeye, I.; Bruneel, C.; Lemahieu, C.; Buyse, J.; Muylaert, K.; Foubert, I. Dietary Enrichment of Eggs with Omega-3 Fatty Acids: A Review. Food Res. Int. 2012, 48, 961–969. [Google Scholar] [CrossRef]
- Harris, W.S.; Mozaffarian, D.; Lefevre, M.; Toner, C.D.; Colombo, J.; Cunnane, S.C.; Holden, J.M.; Klurfeld, D.M.; Morris, M.C.; Whelan, J. Towards Establishing Dietary Reference Intakes for Eicosapentaenoic and Docosahexaenoic Acids. J. Nutr. 2009, 139, 804S–819S. [Google Scholar] [CrossRef] [Green Version]
- Baucells, M.D.; Crespo, N.; Barroeta, A.C.; López-Ferrer, S.; Grashorn, M.A. Incorporation of Different Polyunsaturated Fatty Acids into Eggs. Poult. Sci. 2000, 79, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Cachaldora, P.; García-Rebollar, P.; Alvarez, C.; de Blas, J.C.; Méndez, J. Effect of Type and Level of Fish Oil Supplementation on Yolk Fat Composition and N-3 Fatty Acids Retention Efficiency in Laying Hens. Br. Poult. Sci. 2006, 47, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Bovet, P.; Faeh, D.; Madeleine, G.; Viswanathan, B.; Paccaud, F. Decrease in Blood Triglycerides Associated with the Consumption of Eggs of Hens Fed with Food Supplemented with Fish Oil. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Elswyk, M.E.; Dawson, P.L.; Sams, A. Dietary Menhaden Oil Influences Sensory Characteristics and Headspace Volatiles of Shell Eggs. J. Food Sci. 1995, 60, 85–89. [Google Scholar] [CrossRef]
- Farrell, D.J. Enrichment of Hen Eggs with N-3 Long-Chain Fatty Acids and Evaluation of Enriched Eggs in Humans. Am. J. Clin. Nutr. 1998, 68, 538–544. [Google Scholar] [CrossRef]
- Cachaldora, P.; García-Rebollar, P.; Alvarez, C.; de Blas, J.C.; Méndez, J. Effect of Type and Level of Basal Fat and Level of Fish Oil Supplementation on Yolk Fat Composition and N-3 Fatty Acids Deposition Efficiency in Laying Hens. Anim. Feed Sci. Technol. 2008, 141, 104–114. [Google Scholar] [CrossRef]
- Gonzalez-Esquerra, R.; Leeson, S. Effect of Feeding Hens Regular or Deodorized Menhaden Oil on Production Parameters, Yolk Fatty Acid Profile, and Sensory Quality of Eggs. Poult. Sci. 2000, 79, 1597–1602. [Google Scholar] [CrossRef] [PubMed]
- Becker, W. Microalgae in Human and Animal Nutrition. In Handbook of Microalgal Culture; Richmond, A., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2003; ISBN 9780470995280. [Google Scholar]
- Chen, C.-Y.; Yeh, K.-L.; Aisyah, R.; Lee, D.-J.; Chang, J.-S. Cultivation, Photobioreactor Design and Harvesting of Microalgae for Biodiesel Production: A Critical Review. Bioresour. Technol. 2011, 102, 71–81. [Google Scholar] [CrossRef]
- Haoujar, I.; Cacciola, F.; Abrini, J.; Mangraviti, D.; Giuffrida, D.; el Majdoub, Y.O.; Kounnoun, A.; Miceli, N.; Taviano, M.F.; Mondello, L.; et al. The Contribution of Carotenoids, Phenolic Compounds, and Flavonoids to the Antioxidative Properties of Marine Microalgae Isolated from Mediterranean Morocco. Molecules 2019, 24, 4037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.-N.; Chen, T.-P.; Yang, B.; Liu, J.; Chen, F. Lipid Production from Nannochloropsis. Mar. Drugs 2016, 14, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-Y.; Chen, Y.-C.; Huang, H.-C.; Huang, C.-C.; Lee, W.-L.; Chang, J.-S. Engineering Strategies for Enhancing the Production of Eicosapentaenoic Acid (EPA) from an Isolated Microalga Nannochloropsis Oceanica CY2. Bioresour. Technol. 2013, 147, 160–167. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Chen, Y.-C.; Huang, H.-C.; Ho, S.-H.; Chang, J.-S. Enhancing the Production of Eicosapentaenoic Acid (EPA) from Nannochloropsis Oceanica CY2 Using Innovative Photobioreactors with Optimal Light Source Arrangements. Bioresour. Technol. 2015, 191, 407–413. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Jiang, J.; Wang, H.; Cao, X.; Xue, S.; Yang, Q.; Wang, W. The Characteristics of TAG and EPA Accumulation in Nannochloropsis Oceanica IMET1 under Different Nitrogen Supply Regimes. Bioresour. Technol. 2015, 179, 483–489. [Google Scholar] [CrossRef] [PubMed]
- AOAC. AOAC: Official Methods of Analysis, 15th ed.; AOAC: Rockville, MD, USA, 1990; Volume 1. [Google Scholar]
- Iverson, S.J.; Lang, S.L.C.; Cooper, M.H. Comparison of the Bligh and Dyer and Folch Methods for Total Lipid Determination in a Broad Range of Marine Tissue. Lipids 2001, 36, 1283–1287. [Google Scholar] [CrossRef]
- Fernández, A.; Grienke, U.; Soler-vila, A.; Guihéneuf, F.; Stengel, D.B.; Tasdemir, D. Seasonal and Geographical Variations in the Biochemical Composition of the Blue Mussel (Mytilus Edulis L.) from Ireland. Food Chem. 2015, 177, 43–52. [Google Scholar] [CrossRef]
- Li-Chan, E.C.Y.; Kim, H.-O. Structure and Chemical Compositions of Eggs. In Egg Bioscience and Biotechnology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2008; pp. 1–97. [Google Scholar]
- Lemahieu, C.; Bruneel, C.; Termote-Verhalle, R.; Muylaert, K.; Buyse, J.; Foubert, I. Impact of Feed Supplementation with Different Omega-3 Rich Microalgae Species on Enrichment of Eggs of Laying Hens. Food Chem. 2013, 141, 4051–4059. [Google Scholar] [CrossRef]
- TSS-York QCM+. Available online: http://www.tss-york.com/products/qcmrange#QCC (accessed on 19 November 2018).
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice-Hall/Pearson: Hoboken, NJ, USA, 2010. [Google Scholar]
- Legendre, L.; Legendre, P. Écologie Numérique. Tome 1: Le Traitement Multiple Des Données Écologiques. Collection d’Écologie. Hydrobiology 1981, 66, 775–776. [Google Scholar]
- Patil, V.; Källqvist, T.; Olsen, E.; Vogt, G.; Gislerød, H.R. Fatty Acid Composition of 12 Microalgae for Possible Use in Aquaculture Feed. Aquac. Int. 2007, 15, 1–9. [Google Scholar] [CrossRef]
- Ashour, M.; Elshobary, M.E.; El-Shenody, R.; Kamil, A.W.; Abomohra, A.E.F. Evaluation of a Native Oleaginous Marine Microalga Nannochloropsis Oceanica for Dual Use in Biodiesel Production and Aquaculture Feed. Biomass Bioenergy 2019, 120, 439–447. [Google Scholar] [CrossRef]
- Guerra, I.; Pereira, H.; Costa, M.; Silva, J.T.; Santos, T.; Varela, J.; Mateus, M.; Silva, J. Operation Regimes: A Comparison Based on Nannochloropsis Oceanica Biomass and Lipid Productivity. Energies 2021, 14, 1542. [Google Scholar] [CrossRef]
- Alves, S.P.; Mendonça, S.H.; Silva, J.L.; Bessa, R.J.B. Nannochloropsis Oceanica, a Novel Natural Source of Rumen-Protected Eicosapentaenoic Acid (EPA) for Ruminants. Sci. Rep. 2018, 8, 2–11. [Google Scholar] [CrossRef]
- Sukenik, A.; Beardall, J.; Kromkamp, J.C.; Kopecký, J.; Masojídek, J.; van Bergeijk, S.; Gabai, S.; Shaham, E.; Yamshon, A. Photosynthetic Performance of Outdoor Nannochloropsis Mass Cultures under a Wide Range of Environmental Conditions. Aquat. Microb. Ecol. 2009, 56, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Bruneel, C.; Lemahieu, C.; Fraeye, I.; Ryckebosch, E.; Muylaert, K.; Buyse, J.; Foubert, I. Impact of Microalgal Feed Supplementation on Omega-3 Fatty Acid Enrichment of Hen Eggs. J. Funct. Foods 2013, 5, 897–904. [Google Scholar] [CrossRef]
- Fredriksson, S.; Elwinger, K.; Pickova, J. Fatty Acid and Carotenoid Composition of Egg Yolk as an Effect of Microalgae Addition to Feed Formula for Laying Hens. Food Chem. 2006, 99, 530–537. [Google Scholar] [CrossRef]
- Lemahieu, C.; Bruneel, C.; Ryckebosch, E.; Muylaert, K.; Buyse, J.; Foubert, I. Impact of Different Omega-3 Polyunsaturated Fatty Acid (n-3 PUFA) Sources (Flaxseed, Isochrysis Galbana, Fish Oil and DHA Gold) on n-3 LC-PUFA Enrichment (Efficiency) in the Egg Yolk. J. Funct. Foods 2015, 19, 821–827. [Google Scholar] [CrossRef]
- Li, I.C.; Wu, S.Y.; Liou, J.F.; Liu, H.H.; Chen, J.H.; Chen, C.C. Physiology and Reproduction: Effects of Deinococcus Spp. Supplement on Egg Quality Traits in Laying Hens. Poult. Sci. 2018, 97, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Lubián, L.M.; Montero, O.; Moreno-Garrido, I.; Huertas, I.E.; Sobrino, C.; González-Del Valle, M.; Parés, G. Nannochloropsis (Eustigmatophyceae) as Source of Commercially Valuable Pigments. J. Appl. Phycol. 2000, 12, 249–255. [Google Scholar] [CrossRef]
- Grčević, M.; Kralik, Z.; Kralik, G.; Galović, D.; Radišić, Ž.; Hanžek, D. Quality and Oxidative Stability of Eggs Laid by Hens Fed Marigold Extract Supplemented Diet. Poult. Sci. 2019, 98, 3338–3344. [Google Scholar] [CrossRef]
- Ahmadi, F.; Rahimi, F. Factors Affecting Quality and Quantity of Egg Production in Laying Hens: A Review. World Appl. Sci. J. 2011, 12, 372–384. [Google Scholar]
- Scott, T.A.; Silversides, F.G. The Effect of Storage and Strain of Hen on Egg Quality. Poult. Sci. 2000, 79, 1725–1729. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, V.C.; Gaspar, A.; Calixto, L.F.L.; Agostinho, T.S.P. Stability of the Pigmentation of Egg Yolks Enriched with Omega-3 and Carophyll Stored at Room Temperature and under Refrigeration. Rev. Bras. De Zootec. 2011, 40, 1540–1544. [Google Scholar] [CrossRef] [Green Version]
- Perić, L.; Đukić Stojčić, M.; Bjedov, S. The Effect of Storage and Age of Hens on the Quality of Table Eggs. Adv. Res. Life Sci. 2017, 1, 64–67. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Standard Feed | Supplemented Feed | N. oceanica | |
|---|---|---|---|
| Protein (%) | 14.7 ± 2.6 | 16.6 ± 1.9 | 44.0 ± 0.7 |
| Lipids (%) | 3.96 ± 0.37 | 3.96 ± 0.39 | 20.9 ± 2.7 |
| Ash (%) | 11.44 ± 0.89 | 10.70 ± 0.32 | 8.77 ± 0.20 |
| Fatty acids (µg/mg) | |||
| C16:0 | 4.10 ± 0.34 | 4.89 ± 0.15 | 22.38 ± 0.53 |
| C18:0 | 0.74 ± 0.08 | 0.63 ± 0.07 * | n. d. |
| C16:1 n-7 | n. d. | 1.67 ± 0.32 | 37.15 ± 0.99 |
| C18:1 n-9 | 11.99 ± 0.65 | 10.06 ± 0.10 * | n. d. |
| C18:2 n-6 | 29.2 ± 1.1 | 26.1 ± 1.1 * | n. d. |
| C18:3 n-3 (ALA) | 1.21 ± 0.05 | 1.15 ± 0.43 | n. d. |
| C20:4 n-6 | n. d. | n. d. | 10.35 ± 0.58 |
| C20:5 n-3 (EPA) | n. d. | 0.95 ± 0.21 | 21.40 ± 0.64 |
| Control | Week 1 | Week 2 | Week 3 | Week 4 | ||
|---|---|---|---|---|---|---|
| Fatty acids | EPA + DHA (mg/100 g) | 52 ± 4 * | 116 ± 7 | 120 ± 13 | 109 ± 33 | 110 ± 19 |
| EPA + DHA (mg/100 Kcal) | 37 ± 3 * | 84 ± 4 | 85 ± 9 | 77 ± 22 | 80 ± 12 | |
| Nutritional profile | Lipids (%) | 9.90 ± 0.26 | 9.64 ± 0.27 | 9.78 ± 0.12 | 9.73 ± 0.46 | 9.63 ± 0.39 |
| Protein (%) | 11.9 ± 0.50 | 12.0 ± 0.11 | 12.1 ± 0.36 | 12.2 ± 0.69 | 11.8 ± 0.30 | |
| Ash (%) | 0.83 ± 0.01 | 0.85 ± 0.06 | 0.86 ± 0.05 | 0.88 ± 0.04 | 0.84 ± 0.02 | |
| Physical Parameters | Egg weight (g) | 65.2 ± 4.0 | 64.8 ± 4.1 | 61.5 ± 2.1 | 66.0 ± 5.2 | 63.1 ± 5.3 |
| Shell colour (%) | 28.4 ± 7.2 | 26.2 ± 8.5 | 28.1 ± 5.0 | 26.4 ± 4.1 | 31.7 ± 4.4 | |
| Quality (UH) | 80.8 ± 7.9 * | 84.7 ± 8.9 *,# | 88.3 ± 6.3 *,# | 88.8 ± 5.5 # | 82.1 ± 8.6 *,# | |
| Shell thickness (µm) | 372 ± 19 | 372 ± 10 | 378 ± 29 | 361 ± 24 | 371 ± 27 |
| Control-0 | Control-28 | W4-0 | W4-28 | ||
|---|---|---|---|---|---|
| Fatty acids | ALA (mg/100 g) | 57.7 ± 6.9 | 58.2 ± 8.0 | 51.9 ± 9.4 | 49.3 ± 3.3 |
| EPA (mg/100 g) | 2.1 ± 0.1 | 2.2 ± 0.1 | 5.2 ± 1.2 * | 4.7 ± 0.7 * | |
| DHA (mg/100 g) | 50.3 ± 4.0 | 51.7 ± 6.5 | 105.2 ± 18.2 * | 87.1 ± 9.0 * | |
| Nutritional profile | Lipids (%) | 9.90 ± 0.26 | 9.79 ± 0.40 | 9.62 ± 0.39 | 9.63 ± 0.12 |
| Protein (%) | 11.95 ± 0.50 | 12.28 ± 0.48 | 11.81 ± 0.30 | 11.73 ± 0.36 | |
| Ash (%) | 0.83 ± 0.01 * | 0.86 ± 0.04 *,# | 0.84 ± 0.02 *,# | 0.89 ± 0.02 # | |
| Physical parameters | Eggs weight (g) | 65.2 ± 4.0 | 61.5 ± 6.0 | 63.1 ± 5.3 | 61.8 ± 2.1 |
| Yolk colour (Roche scale) | 9.6 ± 0.8 | 8.5 ± 1.0 | 11.4 ± 0.8 * | 11.7 ± 1.0 * | |
| Shell colour (%) | 28.4 ± 7.2 | 30.0 ± 4.6 | 31.7 ± 4.4 | 30.0 ± 4.6 | |
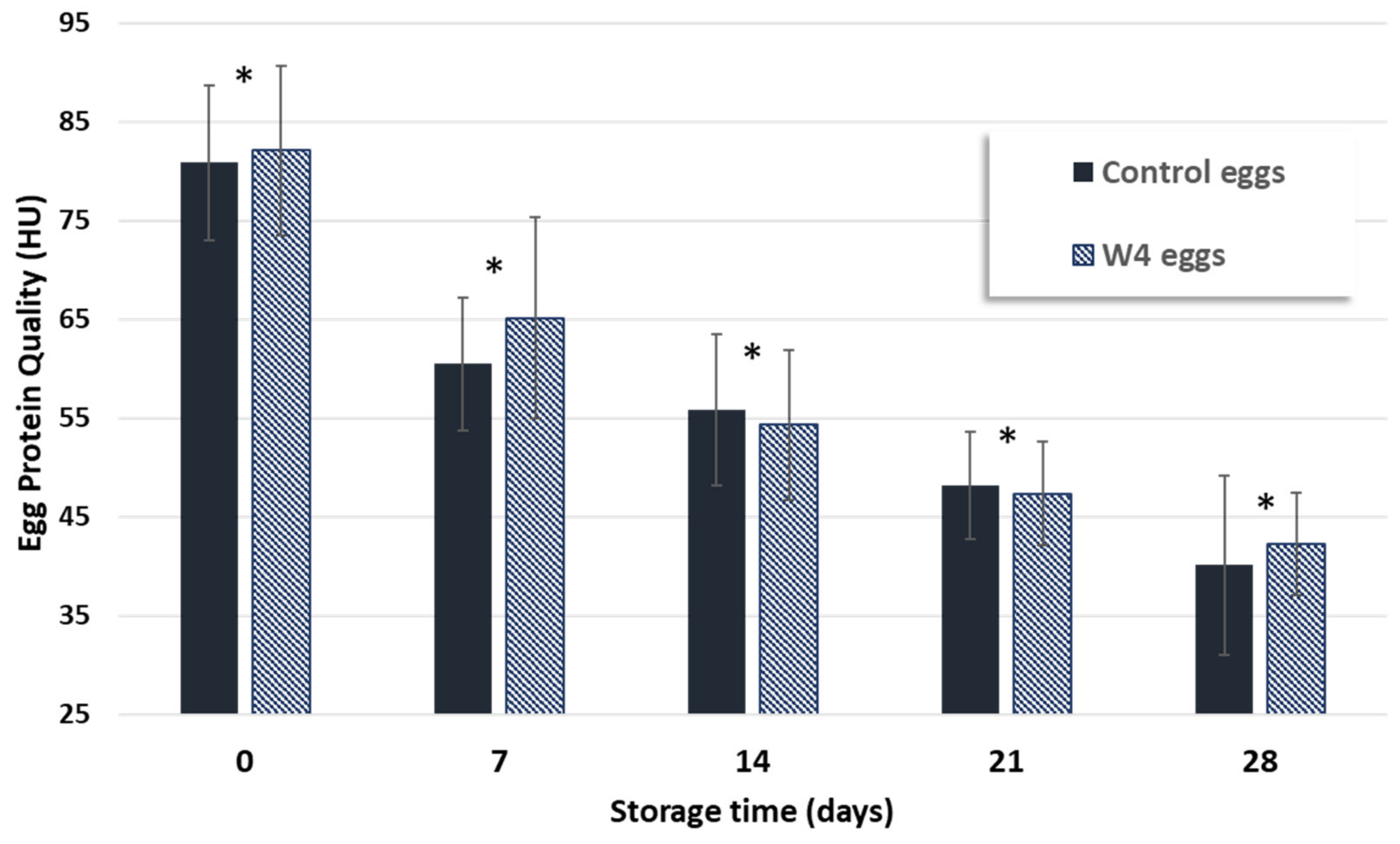
| Protein quality (UH) | 80.8 ± 7.9 | 40.2 ± 9.0 * | 82.1 ± 8.6 | 42.3 ± 5.2 * | |
| Shell thickness (µm) | 372 ± 19 | 349 ± 25 | 371 ± 27 | 387 ± 24 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neves, M.; Ferreira, A.; Antunes, M.; Laranjeira Silva, J.; Mendes, S.; Gil, M.M.; Tecelão, C. Nannochloropsis oceanica as a Sustainable Source of n-3 Polyunsaturated Fatty Acids for Enrichment of Hen Eggs. Appl. Sci. 2021, 11, 8747. https://doi.org/10.3390/app11188747
Neves M, Ferreira A, Antunes M, Laranjeira Silva J, Mendes S, Gil MM, Tecelão C. Nannochloropsis oceanica as a Sustainable Source of n-3 Polyunsaturated Fatty Acids for Enrichment of Hen Eggs. Applied Sciences. 2021; 11(18):8747. https://doi.org/10.3390/app11188747
Chicago/Turabian StyleNeves, Marta, Ana Ferreira, Madalena Antunes, Joana Laranjeira Silva, Susana Mendes, Maria M. Gil, and Carla Tecelão. 2021. "Nannochloropsis oceanica as a Sustainable Source of n-3 Polyunsaturated Fatty Acids for Enrichment of Hen Eggs" Applied Sciences 11, no. 18: 8747. https://doi.org/10.3390/app11188747
APA StyleNeves, M., Ferreira, A., Antunes, M., Laranjeira Silva, J., Mendes, S., Gil, M. M., & Tecelão, C. (2021). Nannochloropsis oceanica as a Sustainable Source of n-3 Polyunsaturated Fatty Acids for Enrichment of Hen Eggs. Applied Sciences, 11(18), 8747. https://doi.org/10.3390/app11188747