
Dietary Soy Impact on Host Transcriptome Profile—A Review


Abstract
:1. Introduction
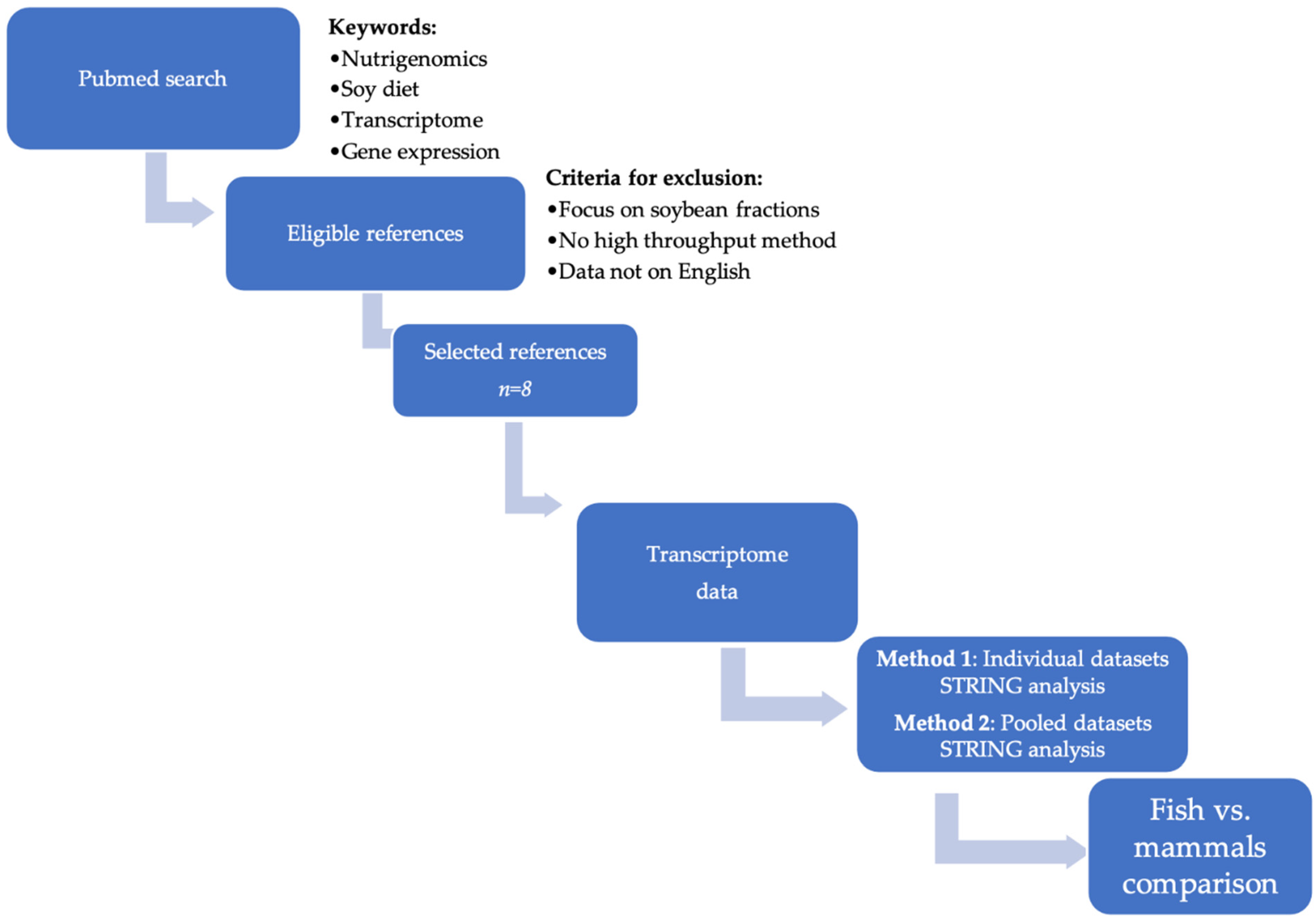
2. Materials and Methods
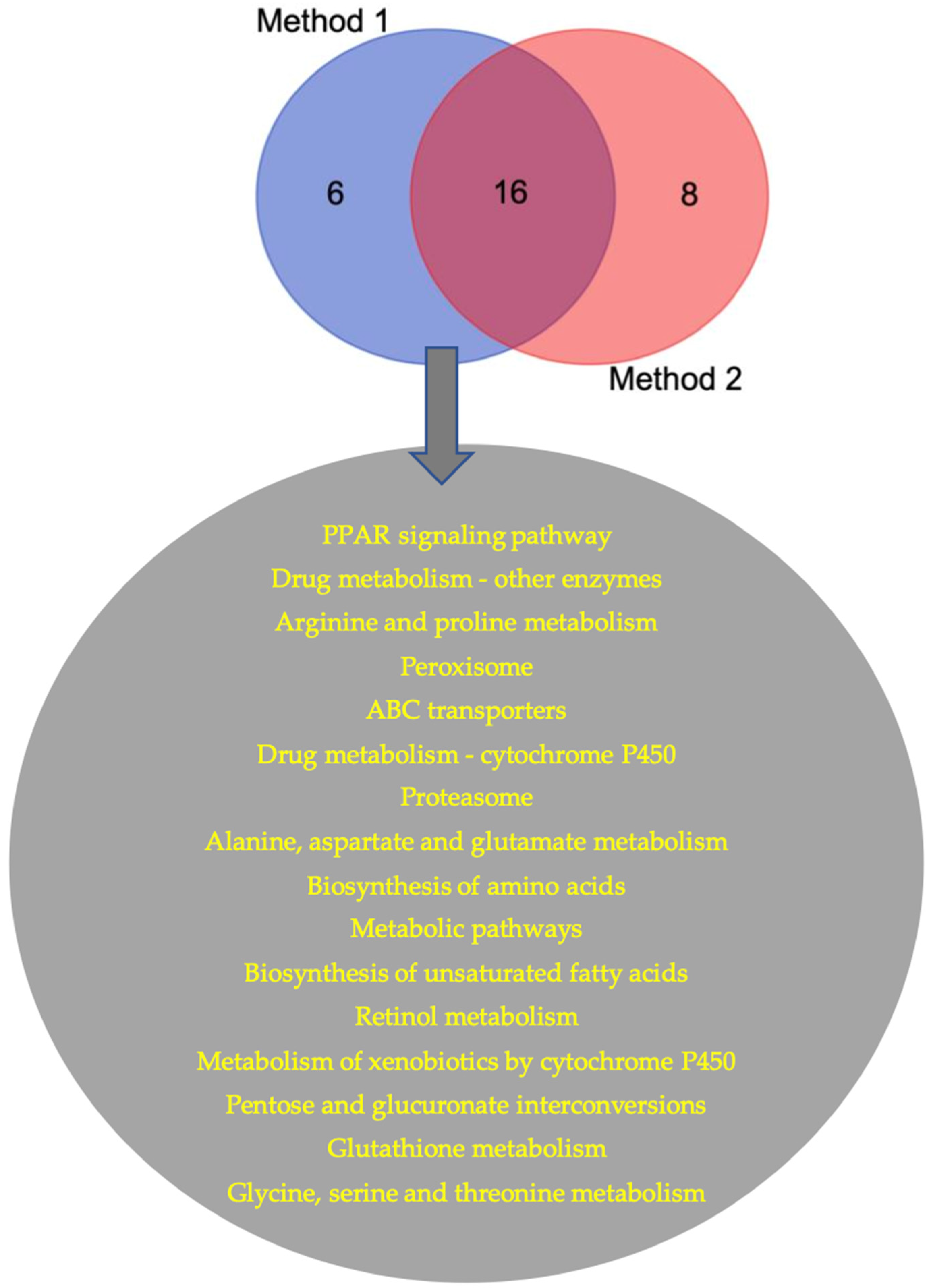
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Karr-Lilienthal, L.K.; Kadzere, C.T.; Grieshop, C.M.; Fahey, G.C. Chemical and nutritional properties of soybean carbohydrates as related to nonruminants. Livest. Prod. Sci. 2005, 97, 1–12. [Google Scholar] [CrossRef]
- Medic, J.; Atkinson, C.; Hurburgh, C.R. Current Knowledge in Soybean Composition. J. Am. Oil Chem. Soc. 2014, 91, 363–384. [Google Scholar] [CrossRef]
- Tucker, K.L.; Qiao, N.; Maras, J.E. Simulation with Soy Replacement Showed That Increased Soy Intake Could Contribute to Improved Nutrient Intake Profiles in the U.S. Population. J. Nutr. 2010, 140, 2296S–2301S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalaiselvan, V.; Kalaivani, M.; Vijayakumar, A.; Sureshkumar, K.; Venkateskumar, K. Current knowledge and future direction of research on soy isoflavones as a therapeutic agents. Pharmacogn. Rev. 2010, 4, 111–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCue, P.; Kalidas, S. Health benefits of soy isoflavonoids and strategies for enhancement: A review. Crit. Rev. Food Sci. Nutr. 2004, 44, 361–367. [Google Scholar] [CrossRef]
- Wu, A.H.; Ziegler, R.G.; Nomura, A.M.Y.; West, D.W.; Kolonel, L.N.; Horn-Ross, P.L.; Hoover, R.N.; Pike, M.C. Soy intake and risk of breast cancer in Asians and Asian Americans. Am. J. Clin. Nutr. 1998, 68, 1437S–1443S. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.O.; Jin, F.; Dai, Q.; Wen, W.; Potter, J.; Kushi, L.; Ruan, Z.; Gao, Y.T.; Zheng, W. Soyfood intake during adolescence and subsequent risk of breast cancer among Chinese women. Cancer Epidemiol. Biomark. Prev. 2001, 10, 483–488. [Google Scholar]
- Nagata, C. Factors to Consider in the Association Between Soy Isoflavone Intake and Breast Cancer Risk. J. Epidemiol. 2010, 20, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Patisaul, H.B.; Jefferson, W. The pros and cons of phytoestrogens. Front. Neuroendocrinol. 2010, 31, 400–419. [Google Scholar] [CrossRef] [Green Version]
- Ramdath, D.D.; Padhi, E.M.T.; Sarfaraz, S.; Renwick, S.; Duncan, A.M. Beyond the Cholesterol-Lowering Effect of Soy Protein: A Review of the Effects of Dietary Soy and Its Constituents on Risk Factors for Cardiovascular Disease. Nutrients 2017, 9, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirtori, C.R.; Even, R.; Lovati, M.R. Soybean Protein Diet and Plasma Cholesterol: From Therapy to Molecular Mechanisms. Ann. N. Y. Acad. Sci. 1993, 676, 188–201. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Murphy, P.A. Isoflavone Content in Commercial Soybean Foods. J. Agric. Food Chem. 1994, 42, 1666–1673. [Google Scholar] [CrossRef]
- Wang, H.; Murphy, P.A. Isoflavone Composition of American and Japanese Soybeans in Iowa: Effects of Variety, Crop Year, and Location. J. Agric. Food Chem. 1994, 42, 1674–1677. [Google Scholar] [CrossRef]
- Taku, K.; Umegaki, K.; Sato, Y.; Taki, Y.; Endoh, K.; Watanabe, S. Soy isoflavones lower serum total and LDL cholesterol in humans: A meta-analysis of 11 randomized controlled trials. Am. J. Clin. Nutr. 2007, 85, 1148–1156. [Google Scholar] [CrossRef] [PubMed]
- Hermansen, K.; Dinesen, B.; Hoie, L.H.; Morgenstern, E.; Gruenwald, J. Effects of soy and other natural products on LDL:HDL ratio and other lipid parameters: A literature review. Adv. Ther. 2003, 20, 50–78. [Google Scholar] [CrossRef]
- Lovati, M.R.; Manzoni, C.; Gianazza, E.; Arnoldi, A.; Kurowska, E.; Carroll, K.K.; Sirtori, C.R. Soy Protein Peptides Regulate Cholesterol Homeostasis in Hep G2 Cells. J. Nutr. 2000, 130, 2543–2549. [Google Scholar] [CrossRef] [Green Version]
- Adams, M.R.; Golden, D.L.; Franke, A.A.; Potter, S.M.; Smith, H.S.; Anthony, M.S. Dietary Soy—Conglycinin (7S Globulin) inhibits atherosclerosis in mice. J. Nutr. 2004, 134, 511–516. [Google Scholar] [CrossRef] [Green Version]
- Wiseman, H.; O’Reilly, J.D.; Adlercreutz, H.; I Mallet, A.I.; Bowey, E.A.; Rowland, I.R.; Sanders, T.A. Isoflavone phytoestrogens consumed in soy decrease F(2)-isoprostane concentrations and increase resistance of low-density lipoprotein to oxidation in humans. Am. J. Clin. Nutr. 2000, 72, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Szkudelska, K.; Nogowski, L. Genistein—A dietary compound inducing hormonal and metabolic changes. J. Steroid Biochem. Mol. Biol. 2007, 105, 37–45. [Google Scholar] [CrossRef]
- Mann, G.E.; Bonacasa, B.; Ishii, T.; Siow, R.C. Targeting the redox sensitive Nrf2–Keap1 defense pathway in cardiovascular disease: Protection afforded by dietary isoflavones. Curr. Opin. Pharmacol. 2009, 9, 139–145. [Google Scholar] [CrossRef]
- Rimbach, G.; Boesch-Saadatmandi, C.; Frank, J.; Fuchs, D.; Wenzel, U.; Daniel, H.; Hall, W.L.; Weinberg, P.D. Dietary isoflavones in the prevention of cardiovascular disease—A molecular perspective. Food Chem. Toxicol. 2008, 46, 1308–1319. [Google Scholar] [CrossRef]
- Güçlü-Ustundağ, Ö.; Mazza, G. Saponins: Properties, applications and processing. Crit. Rev. Food Sci. Nutr. 2007, 47, 231–258. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Badger, T.; Ronis, M.; Wu, X. Non-isoflavone phytochemicals in soy and their health effects. J. Agric. Food Chem. 2010, 58, 8119–8813. [Google Scholar] [CrossRef] [PubMed]
- Høie, L.H.; Morgenstern, E.C.; Gruenwald, J.; Graubaum, H.J.; Busch, R.; Luder, W.; Zunft, H.J. A double-blind placebo- controlled clinical trial compares the cholesterol-lowering effects of two different soy protein preparations in hypercholesterolemic subjects. Eur. J. Nutr. 2005, 44, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Schreihofer, D.A.; Deutsch, C.; Lovekamp-Swan, T.; Sullivan, J.C.; Dorrance, A.M. Effect of high soy diet on the cerebrovasculature and endothelial nitric oxide synthase in the ovariectomized rat. Vasc. Pharmacol. 2010, 52, 236–242. [Google Scholar] [CrossRef] [Green Version]
- Scimone, C.; Donato, L.; Alibrandi, S.; Esposito, T.; Alafaci, C.; D’Angelo, R.; Sidoti, A. Transcriptome analysis provides new molecular signatures in sporadic Cerebral Cavernous Malformation endothelial cells. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165956. [Google Scholar] [CrossRef]
- Scimone, C.; Donato, L.; Katsarou, Z.; Bostantjopoulou, S.; D’Angelo, R.; Sidoti, A. Two Novel KRIT1 and CCM2 Mutations in Patients Affected by Cerebral Cavernous Malformations: New Information on CCM2 Penetrance. Front. Neurol. 2018, 9, 953. [Google Scholar] [CrossRef]
- Sak, K. Epidemiological Evidences on Dietary Flavonoids and Breast Cancer Risk: A Narrative Review. Asian Pac. J. Cancer Prev. 2017, 18, 2309–2328. [Google Scholar] [CrossRef]
- Messina, M. Soyfoods and soybean phyto-estrogens (isoflavones) as possible alternatives to hormone replacement therapy (HRT). Eur. J. Cancer 2000, 36, S71–S72. [Google Scholar] [CrossRef]
- Djuric, Z.; Chen, G.; Doerge, D.R.; Heilbrun, L.K.; Kucuk, O. Effect of soy isoflavone supplementation on oxidative stress in men and women. Cancer Lett. 2001, 172, 1–6. [Google Scholar] [CrossRef]
- Tuli, H.S.; Tuorkey, M.J.; Thakral, F.; Sak, K.; Kumar, M.; Sharma, A.K.; Sharma, U.; Jain, A.; Aggarwal, V.; Bishayee, A. Molecular Mechanisms of Action of Genistein in Cancer: Recent Advances. Front. Pharmacol. 2019, 10, 1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, I.; Bilir, B.; Ali, S.; Sahin, K.; Kucuk, O. Soy Isoflavones in Integrative Oncology: Increased Efficacy and Decreased Toxicity of Cancer Therapy. Integr. Cancer Ther. 2019, 18, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuiper, G.G.; Carlsson, B.; Grandien, K.; Enmark, E.; Häggblad, J.; Nilsson, S.; Gustafsson, J.A. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology 1997, 138, 863–870. [Google Scholar] [CrossRef]
- Sivoňová, M.K.; Kaplán, P.; Tatarková, Z.; Lichardusová, L.; Dušenka, R.; Jurečeková, J. Androgen receptor and soy isoflavones in prostate cancer (Review). Mol. Clin. Oncol. 2018, 10, 191–204. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Adhami, V.M.; Mukhtar, H. Apoptosis by dietary agents for prevention and treatment of prostate cancer. Endocr. Relat. Cancer 2010, 17, 39–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shike, M.; Doane, A.S.; Russo, L.; Cabal, R.; Reis-Filo, J.; Gerald, W.; Cody, H.; Khanin, R.; Bromberg, J.; Norton, L. The Effects of Soy Supplementation on Gene Expression in Breast Cancer: A Randomized Placebo-Controlled Study. J. Natl. Cancer Inst. 2014, 106, dju189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messina, M. Impact of Soy Foods on the Development of Breast Cancer and the Prognosis of Breast Cancer Patients. Forsch Komplementmed. 2016, 23, 75–80. [Google Scholar] [CrossRef]
- Applegate, C.C.; Rowles, J.L.; Ranard, K.M.; Jeon, S.; Erdman, J.W. Soy Consumption and the Risk of Prostate Cancer: An Updated Systematic Review and Meta-Analysis. Nutrients 2018, 10, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.A.; Velasquez, M.T.; Hansen, C.T.; Mohamed, A.I.; Bhathena, S.J. Modulation of carbohydrate metabolism and peptide hormones by soybean isoflavones and probiotics in obesity and diabetes. J. Nutr. Biochem. 2005, 16, 693–699. [Google Scholar] [CrossRef]
- van Nielen, M.; Feskens, E.J.; Rietman, A.; Siebelink, E.; Mensink, M. Partly replacing meat protein with soy protein alters insulin resistance and blood lipids in postmenopausal women with abdominal obesity. J. Nutr. 2014, 144, 1423–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Ruan, W.; Peng, Y.; Wang, D. Soy and the risk of type 2 diabetes mellitus: A systematic review and meta-analysis of observational studies. Diabetes Res. Clin. Pract. 2018, 137, 190–199. [Google Scholar] [CrossRef]
- Konishi, K.; Wada, K.; Yamakawa, M.; Goto, Y.; Mizuta, F.; Koda, S.; Uji, T.; Tsuji, M.; Nagata, C. Dietary Soy Intake Is Inversely Associated with Risk of Type 2 Diabetes in Japanese Women but Not in Men. J. Nutr. 2019, 149, 1208–1214. [Google Scholar] [CrossRef]
- Pubmed. Available online: www.ncbi.nlm.nih.gov/ (accessed on 20 April 2021).
- Bédard, K.; Bédard, J.; Rocheleau, G.; Ferland, G.; Gaudreau, P. Aging and Diets Regulate the Rat Anterior Pituitary and Hypothalamic Transcriptome. Neuroendocrinology 2012, 97, 146–159. [Google Scholar] [CrossRef]
- Song, S.; Hooiveld, G.J.; Li, M.; Zhao, F.; Zhang, W.; Xu, X.; Müller, M.; Li, C.; Zhou, G. Dietary soy and meat proteins induce distinct physiological and gene expression changes in rats. Sci. Rep. 2016, 6, 20036. [Google Scholar] [CrossRef] [Green Version]
- Seclaman, E.; Balacescu, L.; Balacescu, O.; Bejinar, C.; Udrescu, M.; Marian, C.; Sirbu, I.O.; Anghel, A. MicroRNAs mediate liver transcriptome changes upon soy diet intervention in mice. J. Cell. Mol. Med. 2019, 23, 2263–2267. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Cai, Q.; Bao, P.; Wu, J.; Wen, W.; Ye, F.; Zheng, W.; Zheng, Y.; Shu, X.-O. Long-term soy consumption and tumor tissue MicroRNA and gene expression in triple-negative breast cancer. Cancer 2016, 122, 2544–2551. [Google Scholar] [CrossRef]
- Kiron, V.; Park, Y.; Siriyappagouder, P.; Dahle, D.; Vasanth, G.K.; Dias, J.; Fernandes, J.; Sørensen, M.; Trichet, V.V. Intestinal Transcriptome Analysis Reveals Soy Derivative-Linked Changes in Atlantic Salmon. Front. Immunol. 2020, 11, 596514. [Google Scholar] [CrossRef]
- Król, E.; Douglas, A.; Tocher, D.R.; Crampton, V.O.; Speakman, J.R.; Secombes, C.J.; Martin, S.A. Differential responses of the gut transcriptome to plant protein diets in farmed Atlantic salmon. BMC Genom. 2016, 17, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemski, M.M.; Rappleye, C.A.; Dabrowski, K.; Bruno, R.S.; Wick, M. Transcriptomic response to soybean meal-based diets as the first formulated feed in juvenile yellow perch (Perca flavescens). Sci. Rep. 2020, 10, 3998. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, L.; Pacheco, S.; Rincón, G.; Pavez, L.; Lam, N.; Hernández, A.; Dantagnan, P.; González, F.; Jilberto, F.; Ravanal, M.; et al. Intestinal Transcriptome Analysis Reveals Enrichment of Genes Associated with Immune and Lipid Mechanisms, Favoring Soybean Meal Tolerance in High-Growth Zebrafish (Danio Rerio). Genes (Basel) 2021, 12, 700. [Google Scholar] [CrossRef]
- Osada, J. The Use of Transcriptomics to Unveil the Role of Nutrients in Mammalian Liver. ISRN Nutr. 2013, 2013, 403792. [Google Scholar] [CrossRef] [Green Version]
- Piironen, V.; Lampi, A. Occurrence and Levels of Phytosterols in Foods. In Phytosterols as Functional Foods Components and Nutraceuticals; Dutta, P., Ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 1–32. ISBN 9780824747503. [Google Scholar]
- Omoni, A.O.; Aluko, R.E. Soybean Foods and Their Benefits: Potential Mechanisms of Action. Nutr. Rev. 2005, 63, 272–283. [Google Scholar] [CrossRef]
- Corbett, J.W.; Harwood, J.H. Inhibitors of mammalian acetyl-CoA carboxylase. Recent Pat. Cardiovasc. Drug Discov. 2007, 2, 162–180. [Google Scholar] [CrossRef]
- Soni, K.G.; Lehner, R.; Metalnikov, P.; O’Donnell, P.; Semache, M.; Gao, W.; Ashman, K.; Pshezhetsky, A.V.; Mitchell, G.A. Carboxylesterase 3 (EC 3.1.1.1) is a major adipocyte lipase. J. Biol. Chem. 2004, 279, 40683–40689. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, T.; Emi, Y.; Kawabata, S.-I.; Omura, T. Purification and Characterization of Three Male-Specific and One Female-Specific Forms of Cytochrome P-450 from Rat Liver Microsomes. J. Biochem. 1986, 100, 1359–1371. [Google Scholar] [CrossRef]
- Tilly-Kiesi, M.; Schaefer, E.J.; Knudsen, P.; Welty, F.K.; Dolnikowski, G.G.; Taskinen, M.R.; Lichtenstein, A.H. Lipoprotein me-tabolism in subjects with hepatic lipase deficiency. Metabolism 2004, 53, 520–525. [Google Scholar] [CrossRef]
- Karackattu, S.L.; Trigatti, B.; Krieger, M. Hepatic lipase deficiency delays atherosclerosis, myocardial infarction, and cardiac dysfunction and extends lifespan in SR-BI/apolipoprotein E double knockout mice. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 548–554. [Google Scholar] [CrossRef] [Green Version]
- Lepesheva, G.I.; Waterman, M.R. Sterol 14alpha-demethylase cytochrome P450 (CYP51), a P450 in all biological kingdoms. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2007, 1770, 467–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, T.; Corraze, G.; Plagnes-Juan, E.; Skiba-Cassy, S. Cholesterol metabolism regulation mediated by SREBP-2, LXRα and miR-33a in rainbow trout (Oncorhynchus mykiss) both in vivo and in vitro. PLoS ONE 2020, 15, e0223813. [Google Scholar] [CrossRef] [PubMed]
- Hubacek, J.A.; Bobkova, D. Role of Cholesterol 7alpha-Hydroxylase (CYP7A1) in Nutrigenetics and Pharmacogenetics of Cholesterol Lowering. Mol. Diagn. Ther. 2006, 10, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.C.; Cali, J.J.; Jelinek, D.F.; Mehrabian, M.; Sparkes, R.S.; Lusis, A.J.; Russell, D.W.; Hobbs, H.H. Cloning of the human cholesterol 7alpha-hydroxylase gene (CYP7) and localization to chromosome 8q11–q. Genomics 1992, 14, 153–161. [Google Scholar] [CrossRef]
- Li, H.; Huang, W.; Wang, M.; Chen, P.; Chen, L.; Zhang, X. Tandem Mass Tag-based quantitative proteomics analysis of metabolic associated fatty liver disease induced by high fat diet in mice. Nutr. Metab. (Lond.) 2020, 17, 97. [Google Scholar] [CrossRef]
- Berthiaume, L.; Deichaite, I.; Peseckis, S.; Resh, M.D. Regulation of enzymatic activity by active site fatty acylation. A new role for long chain fatty acid acylation of proteins. J. Biol. Chem. 1994, 269, 6498–6505. [Google Scholar] [CrossRef]
- He, Z.; Zhang, R.; Jiang, F.; Zhang, H.; Zhao, A.; Xu, B.; Jin, L.; Wang, T.; Jia, W.; Jia, W.; et al. FADS1-FADS2 genetic polymorphisms are associated with fatty acid metabolism through changes in DNA methylation and gene expression. Clin. Epigenet. 2018, 10, 113. [Google Scholar] [CrossRef] [PubMed]
- Oey, N.A.; den Boer, M.E.; Wijburg, F.A.; Vekemans, M.; Augé, J.; Steiner, C.; Wanders, R.J.A.; Waterham, H.R.; Ruiter, J.P.; Attié-Bitach, T. Long-Chain Fatty Acid Oxidation during Early Human Development. Pediatr. Res. 2005, 57, 755–759. [Google Scholar] [CrossRef] [Green Version]
- Torres, N.; Torre-Villalvazo, I.; Tovar, A.R. Regulation of lipid metabolism by soy protein and its implication in diseases me-diated by lipid disorders. J. Nutr. Biochem. 2006, 17, 365–373. [Google Scholar] [CrossRef]
- Takahashi, Y.; Ide, T. Effects of soy protein and isoflavone on hepatic fatty acid synthesis and oxidation and mRNA expression of uncoupling proteins and peroxisome proliferator-activated receptor gamma in adipose tissues of rats. J. Nutr. Biochem. 2008, 19, 682–693. [Google Scholar] [CrossRef]
- Santos e Silva, L.; Cavalcanti, J.V.F.L.; Magalhães, A.L.R.; Santoro, K.R.; Gonçalves, G.D.; Vasconcelos Santana, L.P.; Barbosa da Silva, J.K.; Calvacanti de Almeida, O. Soybean oil modulates the fatty acid synthesis in the mammary gland, improving nutritional quality of the goat milk. Small Rumin. Res. 2020, 183, 106041. [Google Scholar] [CrossRef]
- Zhang, Y.; Klaassen, C.D. Hormonal regulation of Cyp4a isoforms in mouse liver and kidney. Xenobiotica 2013, 43, 1055–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, M.R.; Italiano, L.; Wilson, S.G.; Mullin, B.H.; Mead, R.; Dudbridge, F.; Watts, G.F.; Stuckey, B.G. Polymorphism in HSD17B6 is associated with key features of polycystic ovary syndrome. Fertil. Steril. 2006, 86, 1438–1446. [Google Scholar] [CrossRef] [PubMed]
- Strauss, J.F.; Barbieri, R.L. Yen and Jaffe’s Reproductive Endocrinology; Elsevier Health Sciences: Philadelphia, PA, USA, 2013; p. 82. [Google Scholar]
- Russo, I.H.; Russo, J. Role of Hormones in Mammary Cancer Initiation and Progression. J. Mammary Gland. Biol. Neoplasia 1998, 3, 49–61. [Google Scholar] [CrossRef]
- Wang, X.; Gérard, C.; Thériault, J.-F.; Poirier, D.; Doillon, C.J.; Lin, S.-X. Synergistic control of sex hormones by 17β-HSD type 7: A novel target for estrogen-dependent breast cancer. J. Mol. Cell Biol. 2015, 7, 568–579. [Google Scholar] [CrossRef] [Green Version]
- Seppen, J. A diet containing the soy phytoestrogen genistein causes infertility in female rats partially deficient in UDP glucuronyltransferase. Toxicol. Appl. Pharmacol. 2015, 264, 335–342. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, D.N.; Clugston, R.D.; Blaner, W.S. Vitamin A Metabolism: An Update. Nutrients 2011, 3, 63–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Standeven, A.M.; Beard, R.L.; Johnson, A.T.; Boehm, M.F.; Escobar, M.; Heyman, R.A.; Chandraratna, R.A. Retinoid-induced hypertriglyceridemia in rats is mediated by retinoic acid receptors. Fundam. Appl. Toxicol. 1996, 33, 264–271. [Google Scholar] [CrossRef]
- Radcliffe, J.D.; Czajka-Narins, D.M. Partial replacement of dietary casein with soy protein isolate can reduce the severity of retinoid-induced hypertriglyceridemia. Plant Foods Hum. Nutr. 1998, 52, 97–108. [Google Scholar] [CrossRef]
- Schupp, M.; Lefterova, M.I.; Janke, J.; Leitner, K.; Cristancho, A.G.; Mullican, S.E.; Qatanani, M.; Szwergold, N.; Steger, D.J.; Curtin, J.C. Retinol saturase promotes adipogenesis and is downregulated in obesity. Proc. Natl. Acad. Sci. USA 2009, 106, 1105–1110, corrected in Proc. Natl. Acad. Sci. USA 2009, 106, 4571. [Google Scholar] [CrossRef] [Green Version]
- Hodges, R.E.; Minich, D.M. Modulation of Metabolic Detoxification Pathways Using Foods and Food- Derived Components: A Scientific Review with Clinical Application. J. Nutr. Metabol. 2015, 2015, 760689. [Google Scholar] [CrossRef]
- Senafi, S.B.; Clarke, D.J.; Burchell, B. Investigation of the substrate specificity of a cloned expressed human bilirubin UDP- glu-ronosyltransferase: UDP-sugar specificity and involvement in steroid and xenobiotic glucuronidation. Biochem J. 1994, 303, 233–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allain, E.P.; Rouleau, M.; Lévesque, E.; Guillemette, C. Emerging roles for UDP-glucuronosyltransferases in drug resistance and cancer progression. Br. J. Cancer 2020, 122, 1277–1287. [Google Scholar] [CrossRef] [Green Version]
- Fay, M.J.; Nguyen, M.T.; Snouwaert, J.N.; Dye, R.; Grant, D.J.; Bodnar, W.M.; Koller, B.H. Xenobiotic Metabolism in Mice Lacking the UDP-Glucuronosyltransferase 2 Family. Drug Metab. Dispos. 2015, 43, 1838–1846. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Huang, H.; Wu, Q. Characterization of the Zebrafish Ugt Repertoire Reveals a New Class of Drug-Metabolizing UDP Glucuronosyltransferases. Mol. Pharmacol. 2014, 86, 62–75. [Google Scholar] [CrossRef]
- Li, D.; Li, Y. The interaction between ferroptosis and lipid metabolism in cancer. Signal Transduct. Target. Ther. 2020, 5, 108. [Google Scholar] [CrossRef]
- Magtanong, L.; Ko, P.-J.; To, M.; Cao, J.Y.; Forcina, G.C.; Tarangelo, A.; Ward, C.C.; Cho, K.; Patti, G.J.; Nomura, D.K.; et al. Exogenous Monounsaturated Fatty Acids Promote a Ferroptosis-Resistant Cell State. Cell Chem. Biol. 2019, 26, 420–432.e9. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Hou, W.; Song, X.; Yu, Y.; Huang, J.; Sun, X.; Kang, R.; Tang, D. Ferroptosis: Process and function. Cell Death Differ. 2016, 23, 369–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koppula, P.; Zhuang, L.; Gan, B. Cystine transporter SLC7A11/xCT in cancer: Ferroptosis, nutrient dependency, and cancer therapy. Protein Cell 2020, 12, 599–620. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, J. Iron and the sex difference in heart disease risk. Lancet 1981, 317, 1293–1294. [Google Scholar] [CrossRef]
- Liu, F.; Di Wang, X. miR-150-5p represses TP53 tumor suppressor gene to promote proliferation of colon adenocarcinoma. Sci. Rep. 2019, 9, 6740. [Google Scholar] [CrossRef]
- Li, T.; Xie, J.; Shen, C.; Cheng, D.; Shi, Y.; Wu, Z.; Zhan, Q.; Deng, X.; Chen, H.; Shen, B.; et al. miR-150-5p Inhibits Hepatoma Cell Migration and Invasion by Targeting MMP. PLoS ONE 2014, 9, e115577. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Brown, M.S.; Goldstein, J.L. Bifurcation of insulin signaling pathway in rat liver: mTORC1 required for stimulation of lipogenesis, but not inhibition of gluconeogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 3441–3446. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Lawrence, J.C., Jr.; Jung, D.Y.; Ko, H.J.; Keller, S.R.; Kim, J.K.; Magnuson, M.A.; Harris, T.E. Fat Cell-Specific Ablation of Rictor in Mice Impairs Insulin-Regulated Fat Cell and Whole-Body Glucose and Lipid Metabolism. Diabetes 2010, 59, 1397–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chango, A.; Pogribny, I.P. Considering Maternal Dietary Modulators for Epigenetic Regulation and Programming of the Fetal Epigenome. Nutrients 2015, 7, 2748–2770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolinoy, D.C.; Weidman, J.R.; Waterland, R.A.; Jirtle, R.L. Maternal genistein alters coat color and protects Avy mouse offspring from obesity by modifying the fetal epigenome. Environ. Health Perspect. 2006, 114, 567–572. [Google Scholar] [CrossRef] [Green Version]
- Howard, T.D.; Ho, S.-M.; Zhang, L.; Chen, J.; Cui, W.; Slager, R.; Gray, S.; Hawkins, G.A.; Medvedovic, M.; Wagner, J.D. Epigenetic Changes with Dietary Soy in Cynomolgus Monkeys. PLoS ONE 2011, 6, e26791. [Google Scholar] [CrossRef] [Green Version]
- Wagner, J.D.; Jorgensen, M.J.; Cline, J.M.; Lees, C.J.; Franke, A.A.; Zhang, L.; Ayers, M.R.; Schultz, C.; Kaplan, J.R. Effects of soy vs. casein protein on body weight and glycemic control in female monkeys and their offspring. Am. J. Primatol. 2009, 71, 802–811. [Google Scholar] [CrossRef]
- Paço, A.; de Bessa Garcia, S.A.; Freitas, R. Methylation in HOX Clusters and Its Applications in Cancer Therapy. Cells 2020, 9, 1613. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Species | Soy Diet | Duration of Feeding | Tissue | Analytical Platform | Ref. |
|---|---|---|---|---|---|
| Male rats (Rattus norvegicus) | Regular chow diet containing 20% soybean meal | 18 months | Anterior pituitary and hypothalamus | Microarray | [44] |
| Male rats (Rattus norvegicus) | Regular chow containing 20% soy protein + 7% soybean oil | 7 days | Liver | RNA-seq | [45] |
| Male mice (Mus musculus) | Regular chow containing 25% soybean meal | 28 days | Liver | RNA-seq/Microarray | [46] |
| Human (Homo sapiens) breast cancer patients | Different soy foods | 12 months | Tumor Tissue | NanoString nCounter | [47] |
| Atlantic salmon (Salmo salar) | Regular chow containing 20% soy protein + 10% soybean meal + 2% soy saponin | 36 days | Distal intestine | RNA-seq | [48] |
| Atlantic salmon (Salmo salar) | Regular chow containing 45% soy protein/Regular chow containing 36% soybean meal | 8 weeks | Distal intestine | Microarray | [49] |
| Yellow perch (Perca flavescens) | Regular chow containing 75% soybean meal | 61 days | Mid intestine | RNA-seq | [50] |
| Zebrafish (Danio Rerio) | Regular chow containing 50% soybean meal + 2% soy saponin | 53 days | Intestine | RNA-seq | [51] |
| Common Pathways for Downregulated Genes | Common Pathways for Upregulated Genes | Common Pathways in Fish vs. Mammals (Up and Down DEGs) |
|---|---|---|
| Drug metabolism-other enzymes | Biosynthesis of unsaturated fatty acids | PPAR signaling pathway |
| Peroxisome | Phagosome | Steroid biosynthesis |
| Retinol metabolism | Proteasome | Drug metabolism-other enzymes |
| Biosynthesis of amino acids | Metabolic pathways | Arginine and proline metabolism |
| Metabolism of xenobiotics by cytochrome P450 | Peroxisome | |
| Drug metabolism-cytochrome P450 | Metabolic pathways | |
| PPAR signaling pathway | Biosynthesis of unsaturated fatty acids | |
| ABC transporters | Retinol metabolism | |
| Arginine biosynthesis | Metabolism of xenobiotics by cytochrome P450 | |
| Metabolic pathways | Fatty acid elongation | |
| Steroid hormone biosynthesis | Pentose and glucuronate interconversions | |
| Arginine and proline metabolism | Glutathione metabolism | |
| Alanine, aspartate and glutamate metabolism | Phagosome | |
| Pentose and glucuronate interconversions | Apoptosis | |
| Glutathione metabolism | Carbon metabolism | |
| Carbon metabolism | Glycine, serine and threonine metabolism | |
| Ascorbate and aldarate metabolism | ABC transporters | |
| Drug metabolism-cytochrome P450 | ||
| Proteasome | ||
| Arginine biosynthesis | ||
| Alanine, aspartate and glutamate metabolism | ||
| Biosynthesis of amino acids |
| Common Pathways for Downregulated Genes | Common Pathways for Upregulated Genes | Common Pathways in Fish vs. Mammals (Up and Down DEGs) |
|---|---|---|
| Drug metabolism-other enzymes | Biosynthesis of unsaturated fatty acids | PPAR signaling pathway |
| Peroxisome | Protein export | Steroid biosynthesis |
| Retinol metabolism | Proteasome | Drug metabolism-other enzymes |
| Biosynthesis of amino acids | Metabolic pathways | Metabolic pathways |
| Metabolism of xenobiotics by cytochrome P450 | Biosynthesis of unsaturated fatty acids | |
| Drug metabolism-cytochrome P450 | Retinol metabolism | |
| PPAR signaling pathway | Metabolism of xenobiotics by cytochrome P450 | |
| ABC transporters | Fatty acid metabolism | |
| Porphyrin and chlorophyll metabolism | ABC transporters | |
| Metabolic pathways | Drug metabolism-cytochrome P450 | |
| Steroid hormone biosynthesis | Pentose and glucuronate interconversions | |
| Arginine and proline metabolism | Glutathione metabolism | |
| Alanine, aspartate and glutamate metabolism | Peroxisome | |
| Pentose and glucuronate interconversions | Glycine, serine and threonine metabolism | |
| Glutathione metabolism | Proteasome | |
| Carbon metabolism | Alanine, aspartate and glutamate metabolism | |
| Ascorbate and aldarate metabolism | Ascorbate and aldarate metabolism | |
| Glycolysis/Gluconeogenesis | Glycolysis/Gluconeogenesis | |
| Ferroptosis | Ferroptosis | |
| Protein export | ||
| Biosynthesis of amino acids | ||
| Porphyrin and chlorophyll metabolism | ||
| Arginine and proline metabolism | ||
| Up vs. Down Common Pathways in Mammals | Up vs. Down Common Pathways in Fish |
|---|---|
| Metabolic pathways | Metabolic pathways |
| Autophagy-animal | Protein processing in endoplasmic reticulum |
| Hepatocellular carcinoma | Carbon metabolism |
| Citrate cycle (TCA cycle) | |
| Amino sugar and nucleotide sugar metabolism | |
| Fatty acid metabolism | |
| Glyoxylate and dicarboxylate metabolism | |
| Starch and sucrose metabolism | |
| Glycosaminoglycan degradation | |
| PPAR signaling pathway |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anghel, A.; Sala-Cirtog, M.; Marian, C.; Samoila, C.; Sirbu, I.O. Dietary Soy Impact on Host Transcriptome Profile—A Review. Appl. Sci. 2021, 11, 7905. https://doi.org/10.3390/app11177905
Anghel A, Sala-Cirtog M, Marian C, Samoila C, Sirbu IO. Dietary Soy Impact on Host Transcriptome Profile—A Review. Applied Sciences. 2021; 11(17):7905. https://doi.org/10.3390/app11177905
Chicago/Turabian StyleAnghel, Andrei, Maria Sala-Cirtog, Catalin Marian, Corina Samoila, and Ioan Ovidiu Sirbu. 2021. "Dietary Soy Impact on Host Transcriptome Profile—A Review" Applied Sciences 11, no. 17: 7905. https://doi.org/10.3390/app11177905
APA StyleAnghel, A., Sala-Cirtog, M., Marian, C., Samoila, C., & Sirbu, I. O. (2021). Dietary Soy Impact on Host Transcriptome Profile—A Review. Applied Sciences, 11(17), 7905. https://doi.org/10.3390/app11177905