
The Role of BiodentineTM on the Odontogenic/Osteogenic Differentiation of Human Dental Pulp Stem Cells

 ,
,
Abstract
:1. Introduction
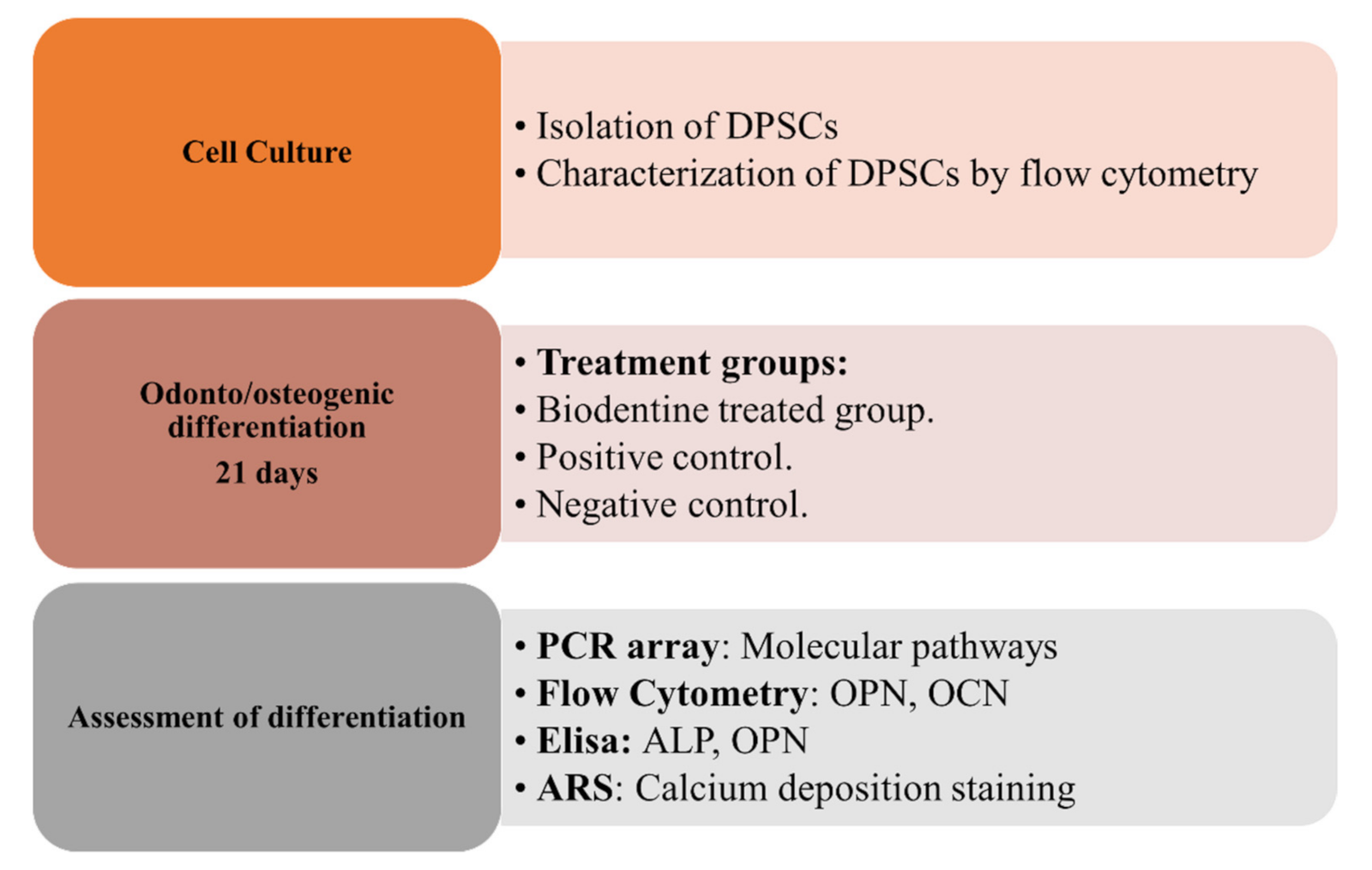
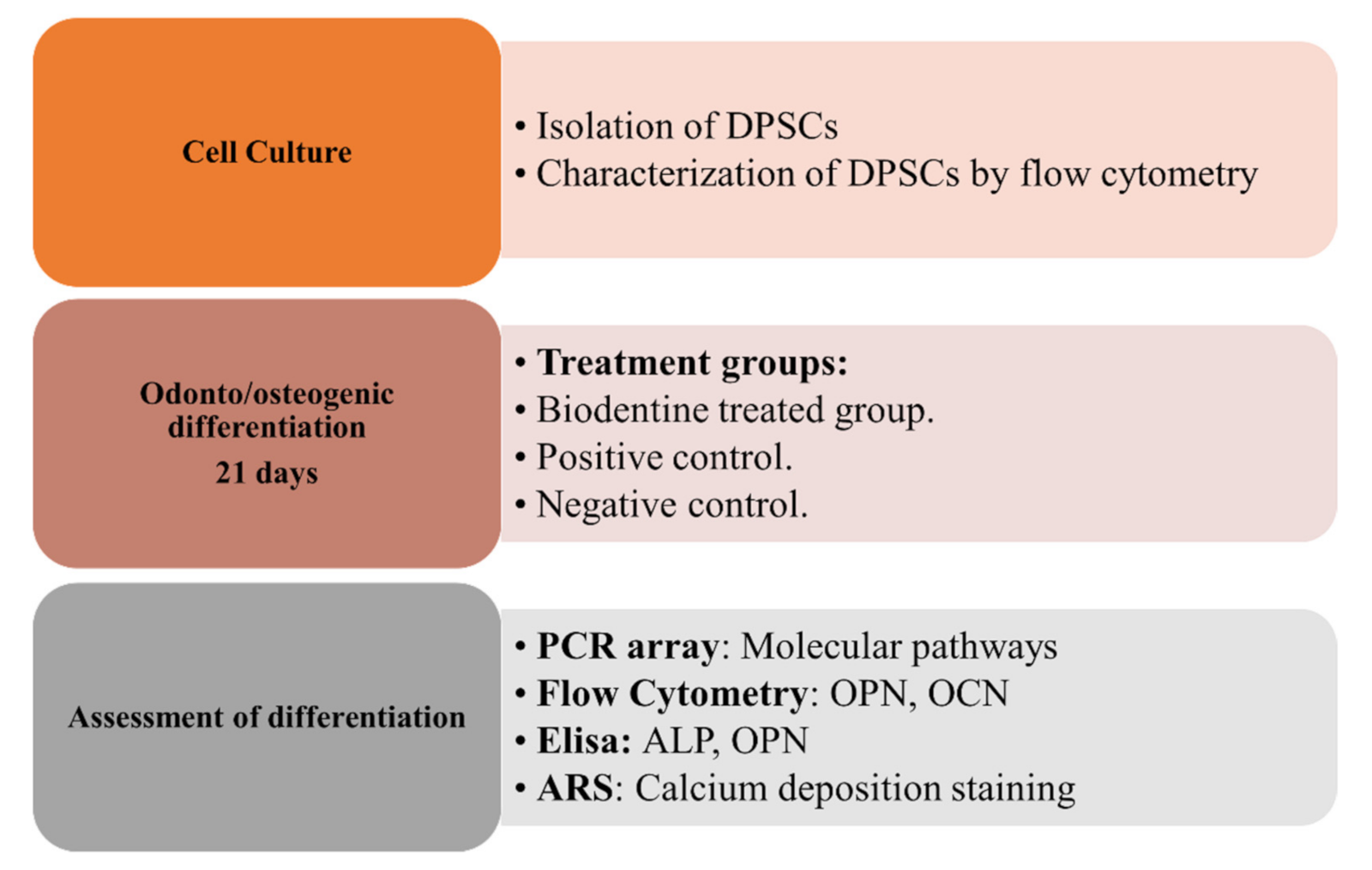
2. Materials and Methods
2.1. Preparation of Biodentine™
2.2. Sample Collection
2.3. Cell Culture of hDPSCs
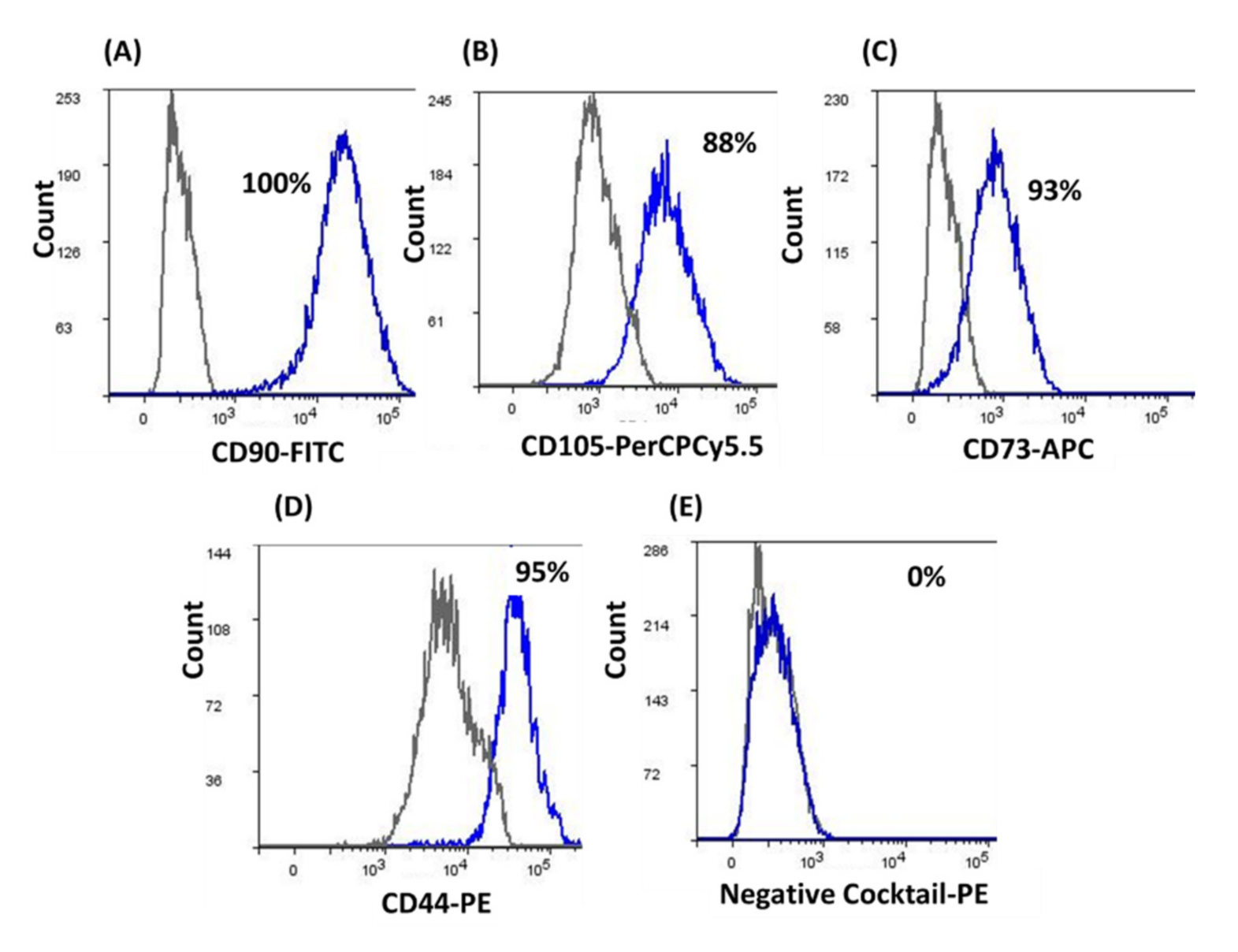
2.4. Characterization of hDPSCs by Flow Cytometry
2.5. Odonto/Osteogenic Differentiation of hDPSCs with BiodentineTM
2.6. PCR Array
2.7. Flow Cytometry for Odonto/Osteogenic Markers
2.8. ELISA for Odonto/Osteogenic Markers
2.9. Alizarin Red Stain ARS
2.10. Statistical Analysis
3. Results
3.1. Characterization of hDPSCs
3.2. Odonto/Osteogenic Differentiation of hDPSCs
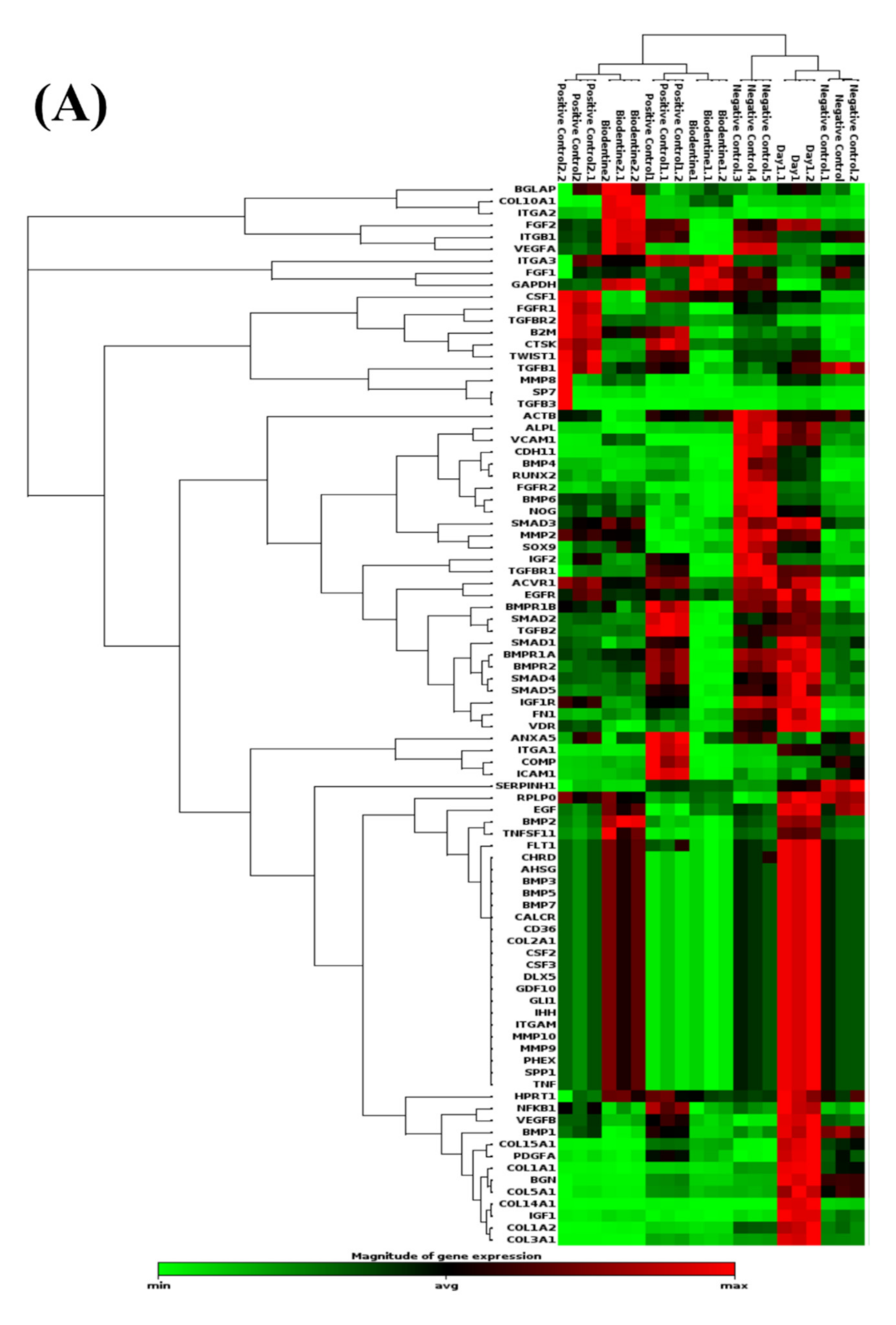
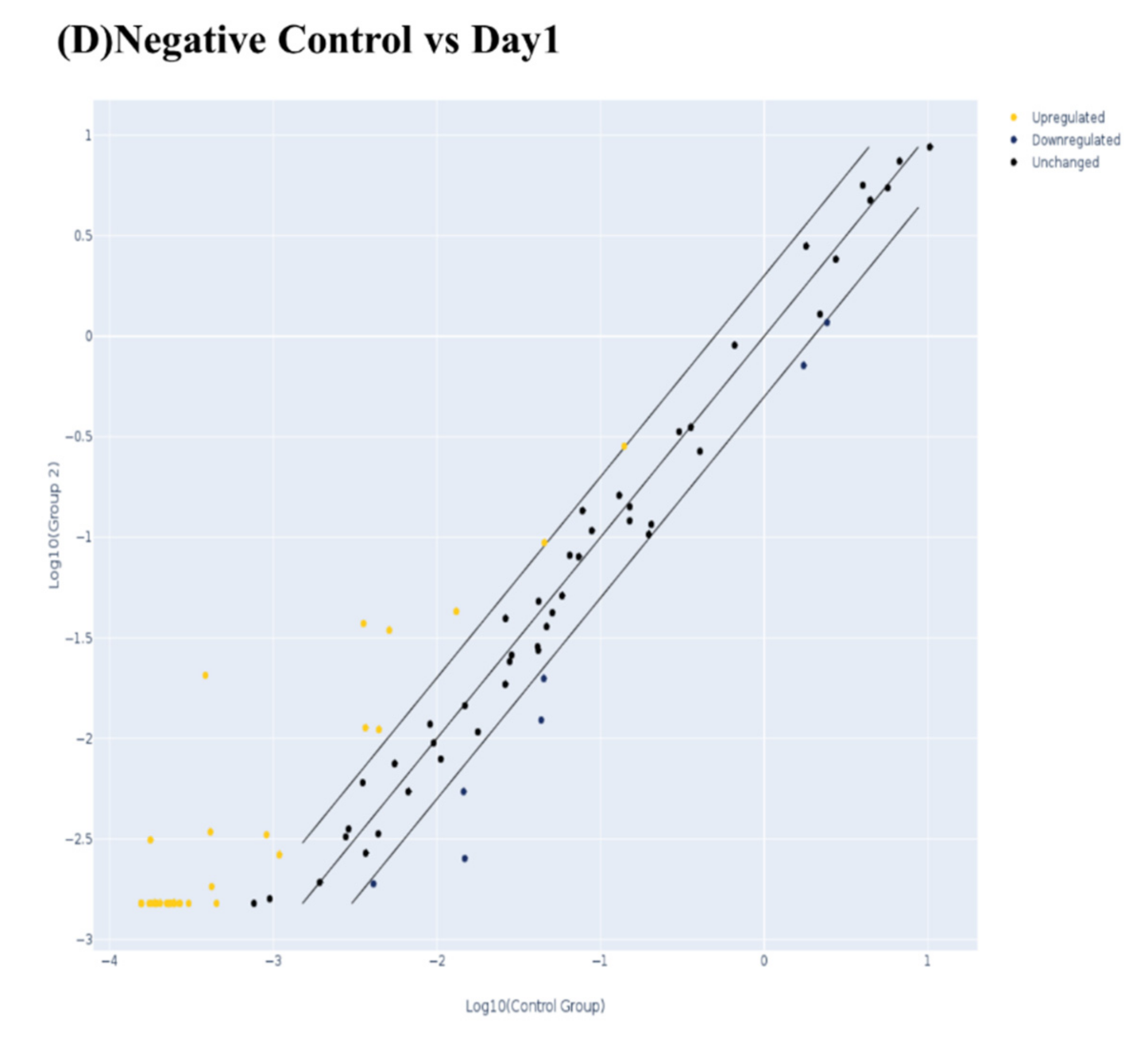
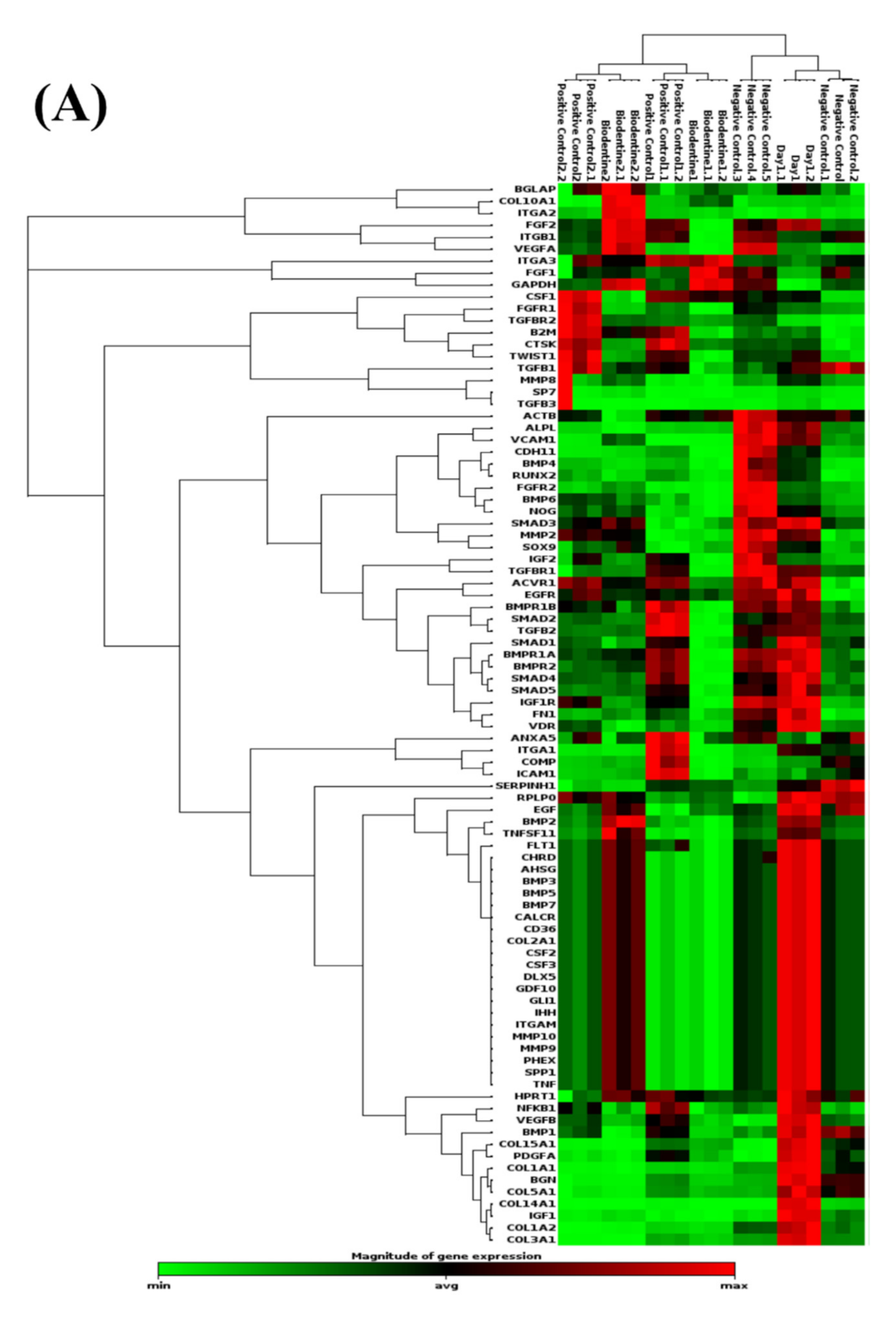
3.2.1. PCR Arrays for Odonto/Osteogenic Signaling Pathway
Odontogenic Differentiation and Ossification
Extracellular Matrix-Related Gene Clusters during Odonto/Osteogenic Differentiation (ECMG)
Ca2+ Signaling Pathway
3.2.2. Flow Cytometry
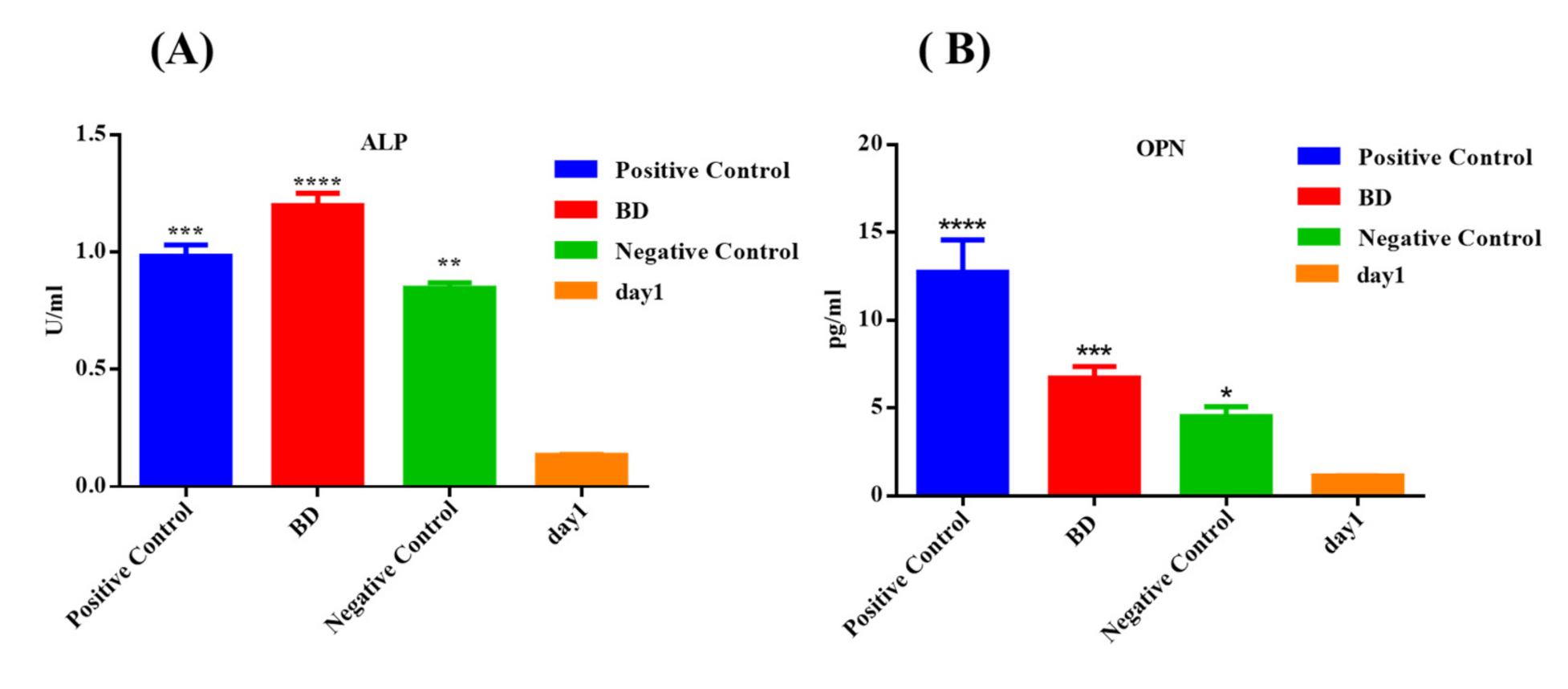
3.2.3. ELISA
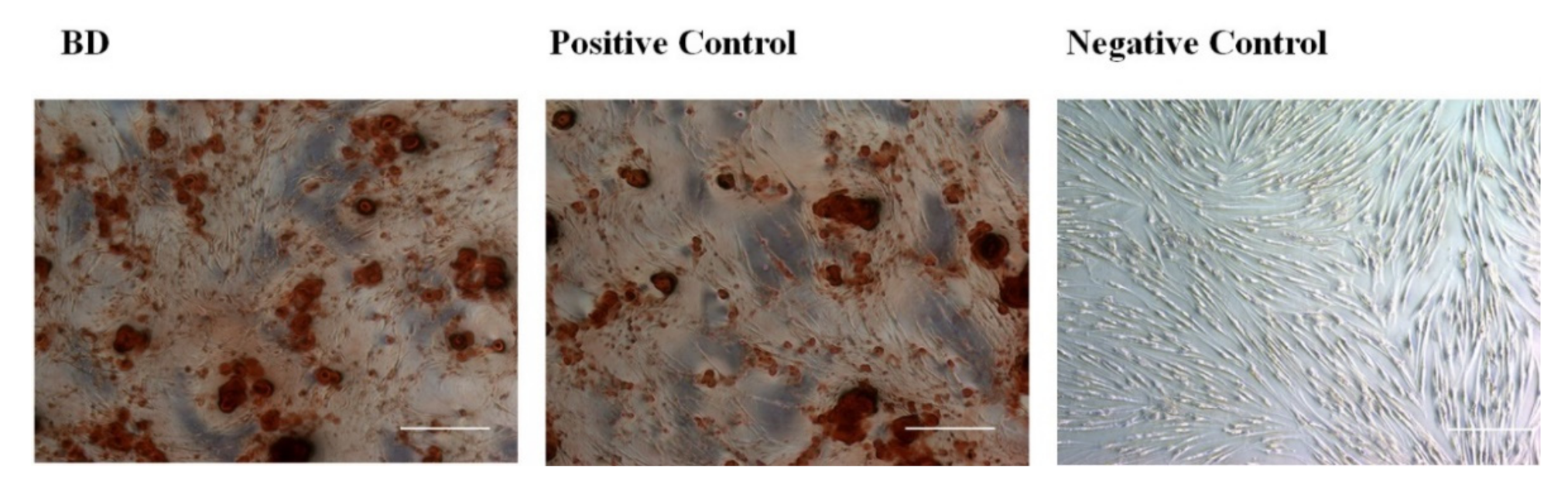
3.2.4. Alizarin Red Stain (Calcium Deposition Staining)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Mason, C.; Dunnill, P. A brief definition of regenerative medicine. Regen. Med. 2008, 3, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Abou Neel, E.A.; Chrzanowski, W.; Salih, V.M.; Kim, H.-W.; Knowles, J.C. Tissue engineering in dentistry. J. Dent. 2014, 42, 915–928. [Google Scholar] [CrossRef] [Green Version]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.G.; Shi, S. SHED: Stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. USA 2003, 100, 5807–5812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, B.-M.; Miura, M.; Gronthos, S.; Bartold, P.M.; Batouli, S.; Brahim, J.; Young, M.; Robey, P.G.; Wang, C.Y.; Shi, S. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef]
- Sonoyama, W.; Liu, Y.; Yamaza, T.; Tuan, R.S.; Wang, S.; Shi, S.; Huang, G.T.-J. Characterization of the apical papilla and its residing stem cells from human immature permanent teeth: A pilot study. J. Endod. 2008, 34, 166–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, G.-J.; Gronthos, S.; Shi, S. Mesenchymal stem cells derived from dental tissues vs. those from other sources: Their biology and role in regenerative medicine. J. Dent. Res. 2009, 88, 792–806. [Google Scholar] [CrossRef]
- Tatullo, M.; Marrelli, M.; Paduano, F. The regenerative medicine in oral and maxillofacial surgery: The most important innovations in the clinical application of mesenchymal stem cells. Int. J. Med Sci. 2015, 12, 72. [Google Scholar] [CrossRef] [Green Version]
- Mitsiadis, T.; Orsini, G.; Jimenez-Rojo, L. Stem cell-based approaches in dentistry. Eur. Cells Mater. 2015, 30, 248–257. [Google Scholar] [CrossRef]
- Pierdomenico, L.; Bonsi, L.; Calvitti, M.; Rondelli, D.; Arpinati, M.; Chirumbolo, G.; Becchetti, E.; Marchionni, C.; Alviano, F.; Fossati, V. Multipotent mesenchymal stem cells with immunosuppressive activity can be easily isolated from dental pulp. Transplantation 2005, 80, 836–842. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Walboomers, X.F.; Shi, S.; Fan, M.; Jansen, J.A. Multilineage differentiation potential of stem cells derived from human dental pulp after cryopreservation. Tissue Eng. 2006, 12, 2813–2823. [Google Scholar] [CrossRef]
- Huang, G.T.-J.; Yamaza, T.; Shea, L.D.; Djouad, F.; Kuhn, N.Z.; Tuan, R.S.; Shi, S. Stem/progenitor cell–mediated de novo regeneration of dental pulp with newly deposited continuous layer of dentin in an in vivo model. Tissue Eng. Part A 2010, 16, 605–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iohara, K.; Murakami, M.; Takeuchi, N.; Osako, Y.; Ito, M.; Ishizaka, R.; Utunomiya, S.; Nakamura, H.; Matsushita, K.; Nakashima, M. A novel combinatorial therapy with pulp stem cells and granulocyte colony—Stimulating factor for total pulp regeneration. Stem Cells Transl. Med. 2013, 2, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Davies, O.; Cooper, P.; Shelton, R.; Smith, A.; Scheven, B. A comparison of the in vitro mineralisation and dentinogenic potential of mesenchymal stem cells derived from adipose tissue, bone marrow and dental pulp. J. Bone Miner. Metab. 2015, 33, 371–382. [Google Scholar] [CrossRef]
- Henkel, J.; Woodruff, M.A.; Epari, D.R.; Steck, R.; Glatt, V.; Dickinson, I.C.; Choong, P.F.; Schuetz, M.A.; Hutmacher, D.W. Bone regeneration based on tissue engineering conceptions—A 21st century perspective. Bone Res. 2013, 1, 216–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, S.; Roy, M.; Bandyopadhyay, A. Recent advances in bone tissue engineering scaffolds. Trends Biotechnol. 2012, 30, 546–554. [Google Scholar] [CrossRef] [Green Version]
- Kinane, D.; Stathopoulou, P.; Papapanou, P. Periodontal Diseases. Nat. Rev. Dis. Primers 2017, 3, 17038. [Google Scholar] [CrossRef]
- Grayson, W.L.; Bunnell, B.A.; Martin, E.; Frazier, T.; Hung, B.P.; Gimble, J.M. Stromal cells and stem cells in clinical bone regeneration. Nat. Rev. Endocrinol. 2015, 11, 140. [Google Scholar] [CrossRef]
- Loeffler, J.; Duda, G.N.; Sass, F.A.; Dienelt, A. The metabolic microenvironment steers bone tissue regeneration. Trends Endocrinol. Metab. 2018, 29, 99–110. [Google Scholar] [CrossRef]
- Gao, C.; Peng, S.; Feng, P.; Shuai, C. Bone biomaterials and interactions with stem cells. Bone Res. 2017, 5, 17059. [Google Scholar] [CrossRef]
- Moioli, E.K.; Clark, P.A.; Xin, X.; Lal, S.; Mao, J.J. Matrices and scaffolds for drug delivery in dental, oral and craniofacial tissue engineering. Adv. Drug Deliv. Rev. 2007, 59, 308–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Li, D.; Kohli, M.R.; Yu, Q.; Kim, S.; He, W.-x. Effect of Biodentine™ on the proliferation, migration and adhesion of human dental pulp stem cells. J. Dent. 2014, 42, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Amano, K.; Iohara, K.; Ito, M.; Imabayashi, K.; Into, T.; Matsushita, K.; Nakamura, H.; Nakashima, M. Matrix metalloproteinase-3 accelerates wound healing following dental pulp injury. Am. J. Pathol. 2009, 175, 1905–1914. [Google Scholar] [CrossRef] [Green Version]
- Palma, P.J.; Marques, J.A.; Santos, J.; Falacho, R.I.; Sequeira, D.; Diogo, P.; Caramelo, F.; Ramos, J.C.; Santos, J.M. Tooth discoloration after regenerative endodontic procedures with calcium silicate-based cements—An ex vivo study. Appl. Sci. 2020, 10, 5793. [Google Scholar] [CrossRef]
- Sequeira, D.B.; Oliveira, A.R.; Seabra, C.M.; Palma, P.J.; Ramos, C.; Figueiredo, M.H.; Santos, A.C.; Cardoso, A.L.; Peça, J.; Santos, J.M. Regeneration of pulp-dentin complex using human stem cells of the apical papilla: In vivo interaction with two bioactive materials. Clin. Oral Investig. 2021, 1–13. [Google Scholar] [CrossRef]
- Luo, Z.; Kohli, M.R.; Yu, Q.; Kim, S.; Qu, T.; He, W.-X. Biodentine induces human dental pulp stem cell differentiation through mitogen-activated protein kinase and calcium-/calmodulin-dependent protein kinase II pathways. J. Endod. 2014, 40, 937–942. [Google Scholar] [CrossRef]
- Laurent, P.; Camps, J.; About, I. BiodentineTM induces TGF-β1 release from human pulp cells and early dental pulp mineralization. Int. Endod. J. 2012, 45, 439–448. [Google Scholar] [CrossRef]
- Loison-Robert, L.S.; Tassin, M.; Bonte, E.; Berbar, T.; Isaac, J.; Berdal, A.; Simon, S.; Fournier, B.P. In vitro effects of two silicate-based materials, Biodentine and BioRoot RCS, on dental pulp stem cells in models of reactionary and reparative dentinogenesis. PLoS ONE 2018, 13, e0190014. [Google Scholar] [CrossRef] [Green Version]
- Abuarqoub, D.; Aslam, N.; Jafar, H.; Abu Harfil, Z.; Awidi, A. Biocompatibility of biodentine™® with periodontal ligament stem cells: In vitro study. Dent. J. 2020, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Hasweh, N.; Awidi, A.; Rajab, L.; Hiyasat, A.; Jafar, H.; Islam, N.; Hasan, M.; Abuarqoub, D. Characterization of the biological effect of BiodentineTM on primary dental pulp stem cells. Indian J. Dent. Res. 2018, 29, 787. [Google Scholar] [PubMed]
- Chiang, Y.-C.; Chang, H.-H.; Wong, C.-C.; Wang, Y.-P.; Wang, Y.-L.; Huang, W.-H.; Lin, C.-P. Nanocrystalline calcium sulfate/hydroxyapatite biphasic compound as a TGF-β1/VEGF reservoir for vital pulp therapy. Dent. Mater. 2016, 32, 1197–1208. [Google Scholar] [CrossRef]
- Saberi, E.; Farhad-Mollashahi, N.; Aval, F.S.; Saberi, M. Proliferation, odontogenic/osteogenic differentiation, and cytokine production by human stem cells of the apical papilla induced by biomaterials: A comparative study. Clin. Cosmet. Investig. Dent. 2019, 11, 181. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Xu, C.; Du, R.; Wen, Y.; Chang, J.; Huan, Z.; Zhu, Y. Effects of silicate-based composite material on the proliferation and mineralization behaviors of human dental pulp cells: An in vitro assessment. Dent. Mater. J. 2018, 37, 889–896. [Google Scholar] [CrossRef] [Green Version]
- Luu, H.H.; Song, W.X.; Luo, X.; Manning, D.; Luo, J.; Deng, Z.L.; Sharff, K.A.; Montag, A.G.; Haydon, R.C.; He, T.C. Distinct roles of bone morphogenetic proteins in osteogenic differentiation of mesenchymal stem cells. J. Orthop. Res. 2007, 25, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Wei, A.; Whittaker, S.; Williams, L.A.; Tao, H.; Ma, D.D.; Diwan, A.D. The role of BMP-7 in chondrogenic and osteogenic differentiation of human bone marrow multipotent mesenchymal stromal cells in vitro. J. Cell. Biochem. 2010, 109, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Chen, G.; Li, Y.-P. TGF-β and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef] [PubMed]
- Bigdeli, N.; de Peppo, G.M.; Lennerås, M.; Sjövall, P.; Lindahl, A.; Hyllner, J.; Karlsson, C. Superior osteogenic capacity of human embryonic stem cells adapted to matrix-free growth compared to human mesenchymal stem cells. Tissue Eng. Part A 2010, 16, 3427–3440. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, A.; Ishizuya, T.; Kintou, N.; Wada, Y.; Katagiri, T.; Wozney, J.M.; Rosen, V.; Yoshiki, S. Effects of BMP-2, BMP-4, and BMP-6 on osteoblastic differentiation of bone marrow-derived stromal cell lines, ST2 and MC3T3-G2/PA6. Biochem. Biophys. Res. Commun. 1996, 220, 366–371. [Google Scholar] [CrossRef]
- Kokabu, S.; Gamer, L.; Cox, K.; Lowery, J.; Tsuji, K.; Raz, R.; Economides, A.; Katagiri, T.; Rosen, V. BMP3 suppresses osteoblast differentiation of bone marrow stromal cells via interaction with Acvr2b. Mol. Endocrinol. 2012, 26, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Rathinam, E.; Govindarajan, S.; Rajasekharan, S.; Declercq, H.; Elewaut, D.; De Coster, P.; Martens, L. Transcriptomic profiling of human dental pulp cells treated with tricalcium silicate–based cements by RNA sequencing. Clin. Oral Investig. 2020, 25, 3181–3195. [Google Scholar] [CrossRef] [PubMed]
- Doshi, R.; Kulkarni, U.; Shinde, S.; Sabane, A.; Patil, A. Role of genes in odontogenesis. J. Adv. Med. Med. Res. 2016, 14, 1–9. [Google Scholar] [CrossRef]
- Ducy, P.; Zhang, R.; Geoffroy, V.; Ridall, A.L.; Karsenty, G. Osf2/Cbfa1: A transcriptional activator of osteoblast differentiation. Cell 1997, 89, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Lian, J.B.; Stein, G.S. Concepts of osteoblast growth and differentiation: Basis for modulation of bone cell development and tissue formation. Crit. Rev. Oral Biol. Med. 1992, 3, 269–305. [Google Scholar] [CrossRef] [Green Version]
- Castano-Izquierdo, H.; Álvarez-Barreto, J.; Dolder, J.V.D.; Jansen, J.A.; Mikos, A.G.; Sikavitsas, V.I. Preculture period of mesenchymal stem cells in osteogenic media influences their in vivo bone forming potential. J. Biomed. Mater. Res. Part A 2007, 82, 129–138. [Google Scholar] [CrossRef]
- Colnot, C.I.; Helms, J.A. A molecular analysis of matrix remodeling and angiogenesis during long bone development. Mech. Dev. 2001, 100, 245–250. [Google Scholar] [CrossRef]
- Knuth, C.; Andres Sastre, E.; Fahy, N.; Witte-Bouma, J.; Ridwan, Y.; Strabbing, E.; Koudstaal, M.; van de Peppel, J.; Wolvius, E.; Narcisi, R. Collagen type X is essential for successful mesenchymal stem cell-mediated cartilage formation and subsequent endochondral ossification. Eur. Cells Mater. 2019, 38, 106–122. [Google Scholar] [CrossRef] [PubMed]
- Bautista, C.M.; Mohan, S.; Baylink, D.J. Insulin-like growth factors I and II are present in the skeletal tissues of ten vertebrates. Metabolism 1990, 39, 96–100. [Google Scholar] [CrossRef]
- Gelse, K.; Pöschl, E.; Aigner, T. Collagens—structure, function, and biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef] [Green Version]
- Kirkham, G.; Cartmell, S. Genes and proteins involved in the regulation of osteogenesis. Top. Tissue Eng. 2007, 3, 1–22. [Google Scholar]
- Sandhu, S.V.; Gupta, S.; Bansal, H.; Singla, K. Collagen in health and disease. J. Orofac. Res. 2012, 2, 153–159. [Google Scholar] [CrossRef]
- Leiendecker, A.P.; Qi, Y.-P.; Sawyer, A.N.; Niu, L.-N.; Agee, K.A.; Loushine, R.J.; Weller, R.N.; Pashley, D.H.; Tay, F.R. Effects of calcium silicate–based materials on collagen matrix integrity of mineralized dentin. J. Endod. 2012, 38, 829–833. [Google Scholar] [CrossRef]
- Widbiller, M.; Lindner, S.; Buchalla, W.; Eidt, A.; Hiller, K.-A.; Schmalz, G.; Galler, K. Three-dimensional culture of dental pulp stem cells in direct contact to tricalcium silicate cements. Clin. Oral Investig. 2016, 20, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Egusa, H.; Iida, K.; Kobayashi, M.; Lin, T.Y.; Zhu, M.; Zuk, P.A.; Wang, C.J.; Thakor, D.K.; Hedrick, M.H.; Nishimura, I. Downregulation of extracellular matrix-related gene clusters during osteogenic differentiation of human bone marrow-and adipose tissue-derived stromal cells. Tissue Eng. 2007, 13, 2589–2600. [Google Scholar] [CrossRef] [PubMed]
- Thesleff, I. Epithelial-mesenchymal signalling regulating tooth morphogenesis. J. Cell Sci. 2003, 116, 1647–1648. [Google Scholar] [CrossRef] [Green Version]
- Hayrapetyan, A.; Jansen, J.A.; van den Beucken, J.J. Signaling pathways involved in osteogenesis and their application for bone regenerative medicine. Tissue Eng. Part B Rev. 2015, 21, 75–87. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Up−Down Regulation (Comparing to Control Group) | |||||
|---|---|---|---|---|---|---|
| BD | Positive Control | Negative Control | ||||
| Fold Regulation | p Value | Fold Regulation | p Value | Fold Regulation | p Value | |
| BGLAP | −1.30 | 0.356363 | −2.17 | 0.000020 | −1.88 | 0.023708 |
| BMP2 | 6.94 | 0.063191 | 6.76 | 0.000001 | 5.24 | 0.000075 |
| BMP3 | 4.70 | 0.060118 | 5.60 | 0.000256 | 3.82 | 0.020257 |
| BMP4 | −10.33 | 0.000000 | −2.68 | 0.511734 | −3.95 | 0.000000 |
| BMP5 | 5.12 | 0.046668 | 6.11 | 0.000051 | 4.16 | 0.009006 |
| BMP6 | −2.04 | 0.093800 | 1.17 | 0.440566 | −1.31 | 0.388649 |
| BMP7 | 6.64 | 0.028578 | 7.91 | 0.000001 | 5.39 | 0.001793 |
| BMPR1A | −2.79 | 0.071691 | −1.15 | 0.938805 | −1.03 | 0.897948 |
| BMPR1B | −2.44 | 0.020348 | −1.31 | 0.563774 | 1.11 | 0.496108 |
| BMPR2 | −4.41 | 0.008592 | −1.30 | 0.483172 | −1.32 | 0.542680 |
| CDH11 | −6.00 | 0.000008 | 1.11 | 0.450717 | −2.85 | 0.005088 |
| DLX5 | 8.12 | 0.024341 | 9.67 | 0.000000 | 6.59 | 0.001195 |
| GDF10 | 5.57 | 0.037773 | 6.64 | 0.000010 | 4.52 | 0.004392 |
| GLI1 | 8.12 | 0.024341 | 9.67 | 0.000000 | 6.59 | 0.001195 |
| ICAM1 | 1.42 | 0.112959 | 2.50 | 0.024031 | 3.22 | 0.102427 |
| IGF1 | 3.07 | 0.073945 | 8.28 | 0.040665 | 2.72 | 0.010881 |
| IGF1R | −2.93 | 0.000207 | −1.20 | 0.953262 | 1.07 | 0.263772 |
| IGF2 | 1.40 | 0.278557 | 3.64 | 0.117069 | 4.22 | 0.006362 |
| IHH | 8.12 | 0.024341 | 9.67 | 0.000000 | 6.59 | 0.001195 |
| MMP2 | −2.99 | 0.000058 | −2.43 | 0.010297 | −2.95 | 0.000422 |
| MMP9 | 4.77 | 0.059368 | 5.68 | 0.000245 | 3.88 | 0.019653 |
| NOG | 1.17 | 0.488601 | 2.41 | 0.102990 | 1.22 | 0.402647 |
| SMAD1 | −2.30 | 0.000404 | −1.01 | 0.988875 | 1.03 | 0.715177 |
| SP7 | 6.55 | 0.047188 | 7.43 | 0.000002 | 8.67 | 0.447590 |
| SPP1 | 5.48 | 0.039164 | 6.53 | 0.000014 | 4.45 | 0.004967 |
| TGFB2 | −4.13 | 0.069790 | 1.22 | 0.363399 | 1.51 | 0.260642 |
| TNF | 6.73 | 0.028147 | 8.02 | 0.000001 | 5.47 | 0.001717 |
| TNFSF11 | 3.37 | 0.133685 | 3.36 | 0.000807 | 2.29 | 0.059883 |
| TWIST1 | −2.09 | 0.000312 | −1.52 | 0.074148 | 1.49 | 0.013689 |
| FGFR2 | −2.27 | 0.016590 | 1.23 | 0.401179 | −2.80 | 0.000057 |
| AHSG | 8.12 | 0.024341 | 9.67 | 0.000000 | 6.59 | 0.001195 |
| Symbol | Up−Down Regulation (Comparing to Control Group) | |||||
|---|---|---|---|---|---|---|
| BD | Positive Control | Negative Control | ||||
| Fold Regulation | p Value | Fold Regulation | p Value | Fold Regulation | p Value | |
| COL10A1 | −1.48 | 0.328066 | −5.86 | 0.000000 | −6.94 | 0.000001 |
| COL14A1 | 6.03 | 0.002053 | 53.47 | 0.155544 | 15.37 | 0.148270 |
| COL15A1 | 5.63 | 0.136740 | 10.45 | 0.147288 | 10.22 | 0.106341 |
| COL1A1 | −13.35 | 0.000021 | 1.41 | 0.241557 | −5.37 | 0.000659 |
| COL1A2 | −4.40 | 0.000004 | 1.11 | 0.289958 | −4.63 | 0.000110 |
| COL2A1 | 7.06 | 0.026841 | 8.42 | 0.000001 | 5.74 | 0.001508 |
| COL3A1 | −4.54 | 0.000280 | −1.14 | 0.064240 | −3.38 | 0.021133 |
| COL5A1 | −2.85 | 0.041857 | 1.75 | 0.211971 | −2.52 | 0.539793 |
| FGFR1 | −2.77 | 0.000008 | −1.25 | 0.402002 | 1.18 | 0.435306 |
| TGFBR1 | −10.70 | 0.000006 | −1.76 | 0.199559 | −3.70 | 0.007490 |
| EGFR | −1.81 | 0.000008 | −2.27 | 0.138319 | −1.39 | 0.010765 |
| VCAM1 | −3.37 | 0.087270 | 1.24 | 0.393835 | −8.41 | 0.000000 |
| FLT1 | 5.35 | 0.041403 | 6.38 | 0.000021 | 5.94 | 0.000475 |
| ITGA1 | −2.32 | 0.034715 | 3.26 | 0.089661 | 3.77 | 0.173366 |
| ITGA2 | 3.77 | 0.153415 | −1.37 | 0.176973 | 1.61 | 0.001199 |
| ITGA3 | 11.69 | 0.001542 | 6.73 | 0.000101 | 7.76 | 0.015945 |
| ITGAM | 5.43 | 0.040022 | 6.47 | 0.000016 | 4.41 | 0.005348 |
| ITGB1 | −3.51 | 0.064370 | −1.70 | 0.000253 | −2.13 | 0.000332 |
| PHEX | 5.13 | 0.038577 | 6.11 | 0.000008 | 4.17 | 0.004322 |
| SERPINH1 | −4.45 | 0.001103 | −1.51 | 0.753602 | −5.49 | 0.009375 |
| ALPL | −5.31 | 0.000026 | 1.25 | 0.388258 | −3.19 | 0.000000 |
| BGN | −2.70 | 0.005630 | 2.03 | 0.160352 | −1.86 | 0.792866 |
| CD36 | 7.26 | 0.026215 | 8.65 | 0.000001 | 5.90 | 0.001419 |
| MMP10 | 5.62 | 0.037035 | 6.70 | 0.000009 | 4.57 | 0.004108 |
| Symbol | Up−Down Regulation (Comparing to Control Group) | |||||
|---|---|---|---|---|---|---|
| BD | Positive Control | Negative Control | ||||
| Fold Regulation | p Value | Fold Regulation | p Value | Fold Regulation | p Value | |
| EGF | 3.90 | 0.005593 | 4.35 | 0.002983 | 2.45 | 0.025906 |
| VDR | −2.85 | 0.000005 | −1.02 | 0.864395 | −1.19 | 0.192859 |
| CALCR | 6.55 | 0.029038 | 7.80 | 0.000001 | 5.32 | 0.001879 |
| COMP | 7.15 | 0.026571 | 17.59 | 0.052347 | 23.30 | 0.075397 |
| EGF | 3.90 | 0.005593 | 4.35 | 0.002983 | 2.45 | 0.025906 |
| VDR | −2.85 | 0.000005 | −1.02 | 0.864395 | −1.19 | 0.192859 |
| CALCR | 6.55 | 0.029038 | 7.80 | 0.000001 | 5.32 | 0.001879 |
| COMP | 7.15 | 0.026571 | 17.59 | 0.052347 | 23.30 | 0.075397 |
| RUNX2 | −8.04 | 0.000000 | −3.52 | 0.062884 | −4.19 | 0.000000 |
| SMAD4 | −2.87 | 0.001775 | −1.41 | 0.141474 | −1.12 | 0.789034 |
| SMAD5 | −3.62 | 0.000977 | −1.44 | 0.124030 | −1.53 | 0.086751 |
| CSF2 | 5.12 | 0.046668 | 6.11 | 0.000051 | 4.16 | 0.009006 |
| CSF3 | 4.17 | 0.041875 | 4.97 | 0.000003 | 3.39 | 0.004201 |
| VEGFB | 1.95 | 0.000019 | 2.07 | 0.032152 | 3.61 | 0.000213 |
| PDGFA | 2.32 | 0.062215 | 3.09 | 0.167867 | 4.63 | 0.098655 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abuarqoub, D.; Zaza, R.; Aslam, N.; Jafar, H.; Zalloum, S.; Atoom, R.; Awidi, A. The Role of BiodentineTM on the Odontogenic/Osteogenic Differentiation of Human Dental Pulp Stem Cells. Appl. Sci. 2021, 11, 7563. https://doi.org/10.3390/app11167563
Abuarqoub D, Zaza R, Aslam N, Jafar H, Zalloum S, Atoom R, Awidi A. The Role of BiodentineTM on the Odontogenic/Osteogenic Differentiation of Human Dental Pulp Stem Cells. Applied Sciences. 2021; 11(16):7563. https://doi.org/10.3390/app11167563
Chicago/Turabian StyleAbuarqoub, Duaa, Rand Zaza, Nazneen Aslam, Hanan Jafar, Suzan Zalloum, Renata Atoom, and Abdalla Awidi. 2021. "The Role of BiodentineTM on the Odontogenic/Osteogenic Differentiation of Human Dental Pulp Stem Cells" Applied Sciences 11, no. 16: 7563. https://doi.org/10.3390/app11167563
APA StyleAbuarqoub, D., Zaza, R., Aslam, N., Jafar, H., Zalloum, S., Atoom, R., & Awidi, A. (2021). The Role of BiodentineTM on the Odontogenic/Osteogenic Differentiation of Human Dental Pulp Stem Cells. Applied Sciences, 11(16), 7563. https://doi.org/10.3390/app11167563