
Does Trans-Stimulation of L-Tyrosine Lead to an Increase in Boron Uptake in Head and Neck Squamous Cell Carcinoma Cells?


Abstract
1. Introduction
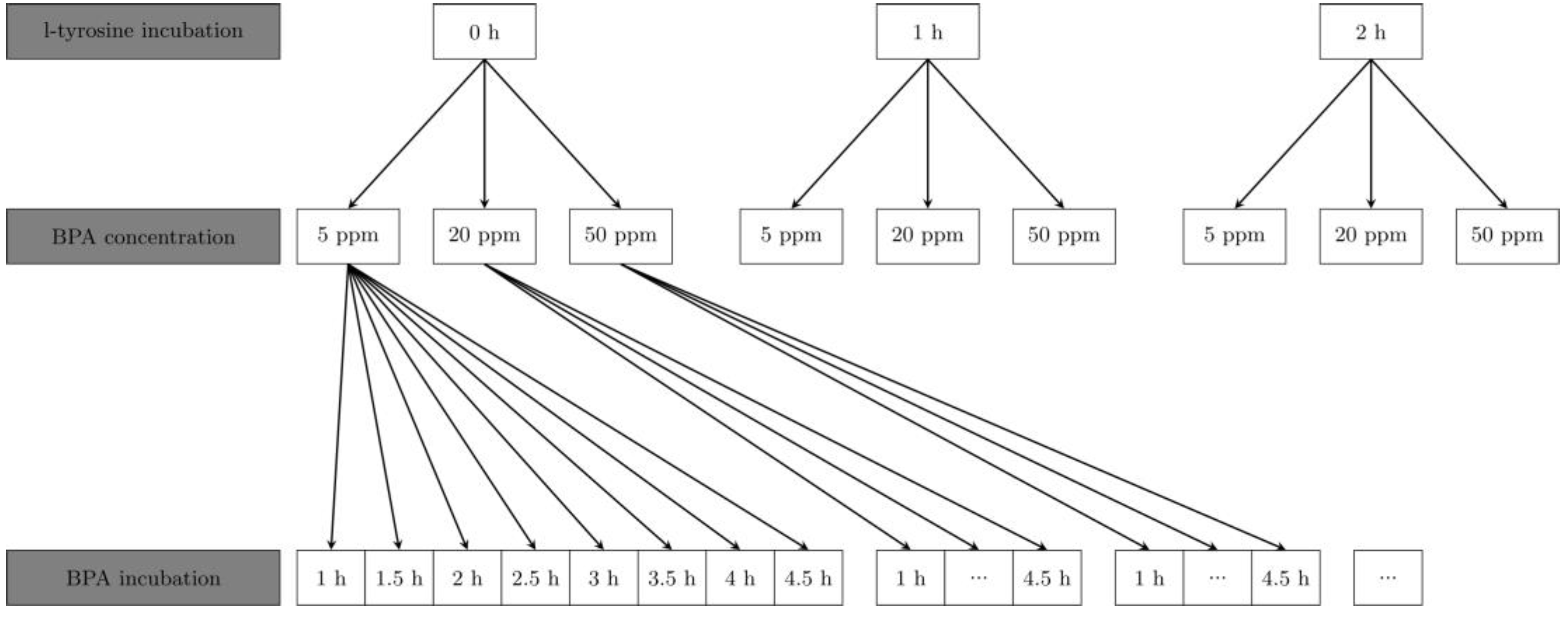
2. Materials and Methods
2.1. Cell Culture
2.2. Western Blot
2.3. Trans-Stimulation Experiments
2.4. Boron Analysis
2.5. Statistics
3. Results
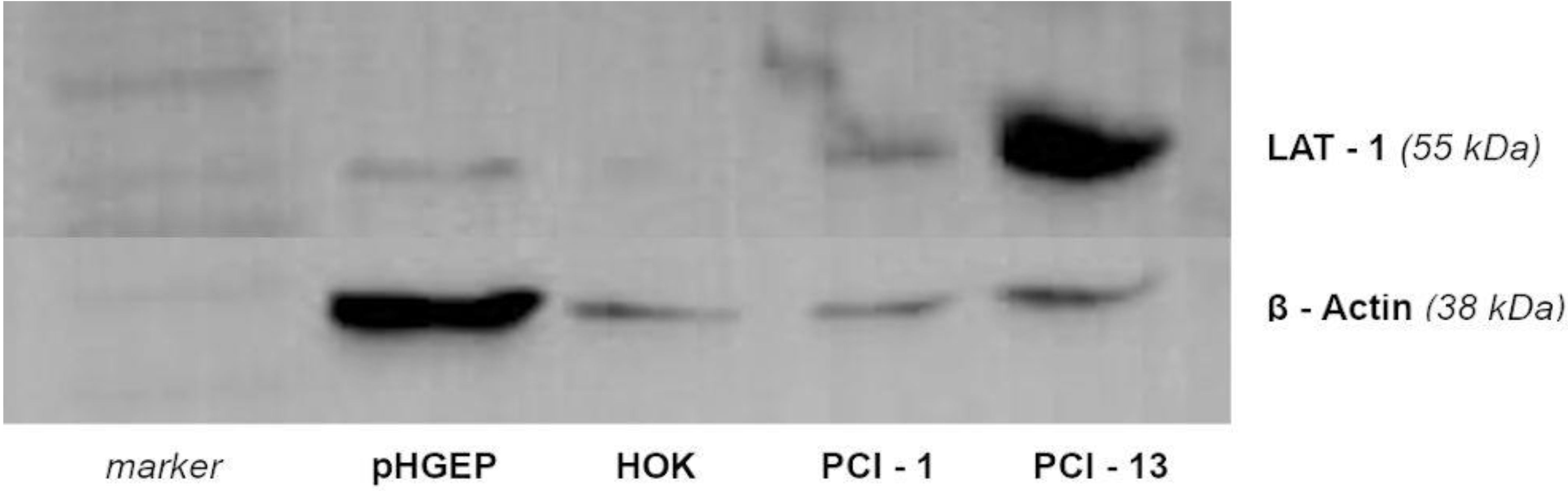
3.1. Western Blotting of LAT-1 in PCI 1 and PCI 13 Cell Lines
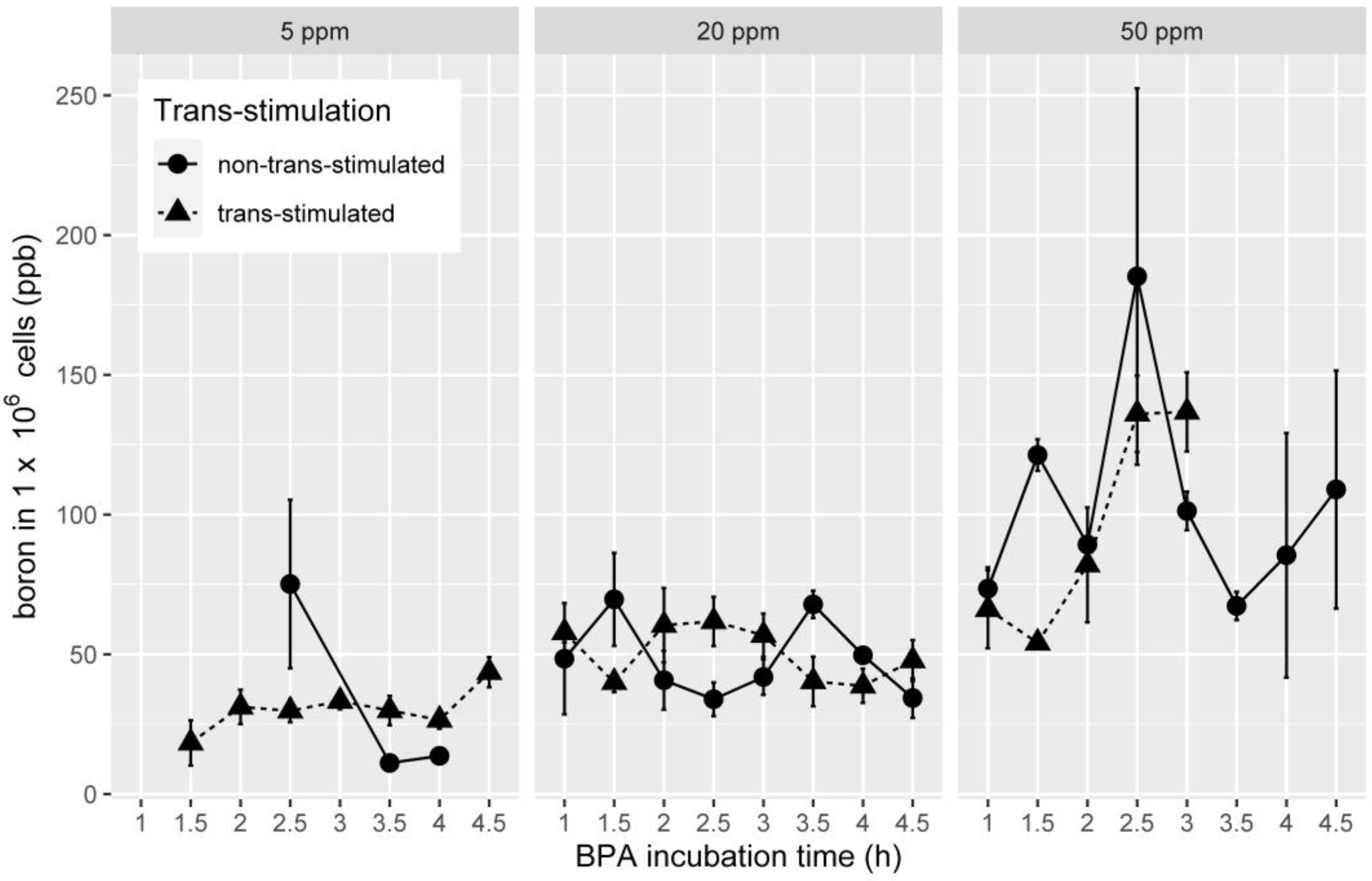
3.2. Boron Uptake in OSCC Cell Lines in Association with Trans-Stimulation
3.3. Boron Uptake in HNSCC Cell Lines in Association with BPA Concentration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Muzaffar, J.; Bari, S.; Kirtane, K.; Chung, C. Recent Advances and Future Directions in Clinical Management of Head and Neck Squamous Cell Carcinoma. Cancers 2021, 13, 338. [Google Scholar] [CrossRef] [PubMed]
- Barth, R.F.; Vicente, M.H.; Harling, O.K.; Kiger, W.S., 3rd; Riley, K.J.; Binns, P.J.; Wagner, F.M.; Suzuki, M.; Aihara, T.; Kato, I.; et al. Current status of boron neutron capture therapy of high grade gliomas and recurrent head and neck cancer. Radiat. Oncol. 2012, 7, 146. [Google Scholar] [CrossRef]
- Suzuki, M. Boron neutron capture therapy (BNCT): A unique role in radiotherapy with a view to entering the accelerator-based BNCT era. Int. J. Clin. Oncol. 2020, 25, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Christensen, H.N. Role of amino acid transport and countertransport in nutrition and metabolism. Physiol. Rev. 1990, 70, 43–77. [Google Scholar] [CrossRef]
- Koivunoro, H.; Kankaanranta, L.; Seppälä, T.; Haapaniemi, A.; Mäkitie, A.; Joensuu, H. Boron neutron capture therapy for locally recurrent head and neck squamous cell carcinoma: An analysis of dose response and survival. Radiother. Oncol. 2019, 137, 153–158. [Google Scholar] [CrossRef]
- Weissbach, L.; Handlogten, M.E.; Christensen, H.N.; Kilberg, M.S. Evidence for two Na+-independent neutral amino acid transport systems in primary cultures of rat hepatocytes. Time-dependent changes in activity. J. Biol. Chem. 1982, 257, 12006–12011. [Google Scholar] [CrossRef]
- Detta, A.; Cruickshank, G.S. l-Amino Acid Transporter-1 and Boronophenylalanine-Based Boron Neutron Capture Therapy of Human Brain Tumors. Cancer Res. 2009, 69, 2126–2132. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Kim, I.J.; Kim, H.; Kim, H.-J.; Jeong, M.J.; Ahn, S.G.; Kim, A.S.; Lee, C.H.; Choi, B.K.; Kim, J.-K.; et al. Amino acid transport system L is differently expressed in human normal oral keratinocytes and human oral cancer cells. Cancer Lett. 2005, 222, 237–245. [Google Scholar] [CrossRef]
- Kim, D.K.; Ahn, S.G.; Park, J.C.; Kanai, Y.; Endou, H.; Yoon, J.H. Expression of L-type amino acid transporter 1 (LAT1) and 4F2 heavy chain (4F2hc) in oral squamous cell carcinoma and its precusor lesions. Anticancer. Res. 2004, 24, 1671–1675. [Google Scholar]
- Obayashi, S.; Kato, I.; Ono, K.; Masunaga, S.-I.; Suzuki, M.; Nagata, K.; Sakurai, Y.; Yura, Y. Delivery of 10boron to oral squamous cell carcinoma using boronophenylalanine and borocaptate sodium for boron neutron capture therapy. Oral Oncol. 2004, 40, 474–482. [Google Scholar] [CrossRef]
- Lin, Y.C.; Wang, S.J.; Chung, H.P.; Liu, H.M.; Chou, F.I. Low dose of gamma irradiation enhanced boronophenylalanine uptake in head and neck carcinoma cells for boron neutron capture therapy. Appl. Radiat. Isot. 2011, 69, 1728–1731. [Google Scholar] [CrossRef]
- Wu, C.-Y.; Chan, P.-C.; Chou, L.-S.; Chang, C.-W.; Yang, F.-Y.; Liu, R.-S.; Chiou, S.-H.; Chen, Y.-W.; Yen, S.-H.; Wang, H.-E. Pulsed-Focused Ultrasound Enhances Boron Drug Accumulation in a Human Head and Neck Cancer Xenograft-Bearing Mouse Model. Mol. Imaging Biol. 2013, 16, 95–101. [Google Scholar] [CrossRef]
- Yang, W.; Barth, R.F.; Huo, T.; Kabalka, G.W.; Shaikh, A.L.; Haider, S.A.; Chandra, S. Effects of l-DOPA pre-loading on the uptake of boronophenylalanine using the F98 glioma and B16 melanoma models. Appl. Radiat. Isot. 2014, 88, 69–73. [Google Scholar] [CrossRef][Green Version]
- Rosenberg, T.; Wilbrandt, W. Uphill transport induced by counterflow. J. Gen. Physiol. 1957, 41, 289–296. [Google Scholar] [CrossRef]
- Heinz, E.; Walsh, P. Exchange Diffusion, Transport, and Intracellular Level of Amino Acids in Ehrlich Carcinoma Cells. J. Biol. Chem. 1958, 233, 1488–1493. [Google Scholar] [CrossRef]
- Page, C.M.; Inglis, M.S.; Wheatley, D.N. The intracellular acid-extractable (acid-soluble) amino acid pool in mammalian cells: 4 An hypothesis to explain the effects of preloading on the exchange of amino acids across the cell membrane. Cytobios 1982, 34, 105–123. [Google Scholar]
- Wittig, A.; Sauerwein, W.A.; Coderre, J.A. Mechanisms of transport of p-borono-phenylalanine through the cell membrane in vitro. Radiat. Res. 2000, 153, 173–180. [Google Scholar]
- Watanabe, T.; Tanaka, H.; Fukutani, S.; Suzuki, M.; Hiraoka, M.; Ono, K. L-phenylalanine preloading reduces the 10B(n, α)7Li dose to the normal brain by inhibiting the uptake of boronophenylalanine in boron neutron capture therapy for brain tumours. Cancer Lett. 2016, 370, 27–32. [Google Scholar] [CrossRef]
- Papaspyrou, M.; Feinendegen, L.E.; Müller-Gärtner, H.W. Preloading with L-tyrosine increases the uptake of boronophenylalanine in mouse melanoma cells. Cancer Res. 1994, 54, 6311–6314. [Google Scholar]
- Capuani, S.; Gili, T.; Bozzali, M.; Russo, S.; Porcari, P.; Cametti, C.; D’Amore, E.; Colasanti, M.; Venturini, G.; Maraviglia, B.; et al. L-DOPA Preloading Increases the Uptake of Borophenylalanine in C6 Glioma Rat Model: A New Strategy to Improve BNCT Efficacy. Int. J. Radiat. Oncol. 2008, 72, 562–567. [Google Scholar] [CrossRef]
- Heo, D.S.; Snyderman, C.; Gollin, S.M.; Pan, S.; Walker, E.; Deka, R.; Barnes, E.L.; Johnson, J.T.; Herberman, R.B.; Whiteside, T.L. Biology, cytogenetics, and sensitivity to immunological effector cells of new head and neck squamous cell carcinoma lines. Cancer Res. 1989, 49, 5167–5175. [Google Scholar] [PubMed]
- Dirix, P.; Nuyts, S.; van der Bogaert, W. Radiation-induced xerostomia in patients with head and neck cancer. Cancer 2006, 107, 2525–2534. [Google Scholar] [CrossRef]
- Wang, L.-W.; Chen, Y.-W.; Ho, C.-Y.; Liu, Y.-W.H.; Chou, F.-I.; Liu, Y.-H.; Liu, H.-M.; Peir, J.-J.; Jiang, S.-H.; Chang, C.-W.; et al. Fractionated Boron Neutron Capture Therapy in Locally Recurrent Head and Neck Cancer: A Prospective Phase I/II Trial. Int. J. Radiat. Oncol. 2016, 95, 396–403. [Google Scholar] [CrossRef]
- Shotwell, M.A.; Oxender, D.L. The regulation of neutral amino acid transport by amino acid availability in animal cells. Trends Biochem. Sci. 1983, 8, 314–316. [Google Scholar] [CrossRef]
- Moreno, A.; Lobatón, C.D.; Oxender, D.L. Regulation of amino acid transport system L by amino acid availability in CHO-K1 cells. A special role for leucine. Biochim. Biophys. Acta Biomembr. 1985, 819, 271–274. [Google Scholar] [CrossRef]
- Campbell, W.A.; Thompson, N.L. Overexpression of LAT1/CD98 Light Chain Is Sufficient to Increase System l-Amino Acid Transport Activity in Mouse Hepatocytes but Not Fibroblasts. J. Biol. Chem. 2001, 276, 16877–16884. [Google Scholar] [CrossRef]
- Grunewald, C.; Sauberer, M.; Filip, T.; Wanek, T.; Stanek, J.; Mairinger, S.; Rollet, S.; Kudejova, P.; Langer, O.; Schütz, C.; et al. On the applicability of [18F]FBPA to predict L-BPA concentration after amino acid preloading in HuH-7 liver tumor model and the implication for liver boron neutron capture therapy. Nucl. Med. Biol. 2017, 44, 83–89. [Google Scholar] [CrossRef]
- Wingelhofer, B.; Kreis, K.; Mairinger, S.; Muchitsch, V.; Stanek, J.; Wanek, T.; Langer, O.; Kuntner, C. Preloading with L-BPA, L-tyrosine and L-DOPA enhances the uptake of [18F]FBPA in human and mouse tumour cell lines. Appl. Radiat. Isot. 2016, 118, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Kreimann, E.L.; Itoiz, M.E.; Dagrosa, A.; Garavaglia, R.; Farías, S.; Batistoni, D.; Schwint, A.E. The hamster cheek pouch as a model of oral cancer for boron neutron capture therapy studies: Selective delivery of boron by boronophenylalanine. Cancer Res. 2001, 61, 8775–8781. [Google Scholar]
- Trivillin, V.A.; Garabalino, M.A.; Colombo, L.L.; Gonzalez, S.J.; Farias, R.O.; Monti Hughes, A.; Pozzi, E.C.; Bortolussi, S.; Altieri, S.; Itoiz, M.E.; et al. Biodistribution of the boron carriers boronophenylalanine (BPA) and/or decahydrodecaborate (GB-10) for Boron Neutron Capture Therapy (BNCT) in an experimental model of lung metastases. Appl. Radiat. Isot. 2014, 88, 94–98. [Google Scholar] [CrossRef]
- Ring, K.; Gross, W.; Heinz, E. Negative feedback regulation of amino acid transport in Streptomyces hydrogenans. Arch. Biochem. Biophys. 1970, 137, 243–252. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| BPA Concentration (ppm) | Trans-Stimulation | Mean Boron Concentration (ppb per One Million Cells) | SD | p |
|---|---|---|---|---|
| 5 | no | 50.31600 | 63.72422 | 0.893 |
| yes | 29.25684 | 13.29931 | ||
| 20 | no | 48.92545 | 30.77410 | 0.998 |
| yes | 51.18492 | 26.31162 | ||
| 50 | no | 98.63308 | 49.07148 | 0.615 |
| yes | 82.42652 | 43.24766 |
| Trans-Stimulation | BPA Concentration (ppm) | Mean Boron Concentration (ppb per One Million Cells) | SD | p | ||
|---|---|---|---|---|---|---|
| 5 ppm vs. 20 ppm | 5 ppm vs. 50 ppm | 20 ppm vs. 50 ppm | ||||
| no | 5 | 50.31600 | 63.72422 | 1.000 | 0.208 | |
| 20 | 29.25684 | 13.29931 | 0.012 | |||
| 50 | 48.92545 | 30.77410 | ||||
| yes | 5 | 51.18492 | 26.31162 | 0.000 | 0.000 | |
| 20 | 98.63308 | 49.07148 | 0.032 | |||
| 50 | 82.42652 | 43.24766 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gielisch, M.; Moergel, M.; Al-Nawas, B.; Kämmerer, P.W. Does Trans-Stimulation of L-Tyrosine Lead to an Increase in Boron Uptake in Head and Neck Squamous Cell Carcinoma Cells? Appl. Sci. 2021, 11, 7286. https://doi.org/10.3390/app11167286
Gielisch M, Moergel M, Al-Nawas B, Kämmerer PW. Does Trans-Stimulation of L-Tyrosine Lead to an Increase in Boron Uptake in Head and Neck Squamous Cell Carcinoma Cells? Applied Sciences. 2021; 11(16):7286. https://doi.org/10.3390/app11167286
Chicago/Turabian StyleGielisch, Matthias, Maximilian Moergel, Bilal Al-Nawas, and Peer W. Kämmerer. 2021. "Does Trans-Stimulation of L-Tyrosine Lead to an Increase in Boron Uptake in Head and Neck Squamous Cell Carcinoma Cells?" Applied Sciences 11, no. 16: 7286. https://doi.org/10.3390/app11167286
APA StyleGielisch, M., Moergel, M., Al-Nawas, B., & Kämmerer, P. W. (2021). Does Trans-Stimulation of L-Tyrosine Lead to an Increase in Boron Uptake in Head and Neck Squamous Cell Carcinoma Cells? Applied Sciences, 11(16), 7286. https://doi.org/10.3390/app11167286