
Ectoine Production Using Novel Heterologous EctABCS. salarius from Marine Bacterium Salinicola salarius

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, Bacterial Strains, and Cultivation
2.2. Gene Cloning, and Expression for EctAS. salarius, EctBS. salarius, EctCS. salarius
2.3. Bioinformatic Analysis and Molecular Modeling of EctABCS. salarius Structure
2.4. Preparation of E. coli for Biocatalysis
2.5. Bioconversion Conditions
2.6. HPLC Analytical Methods
3. Results
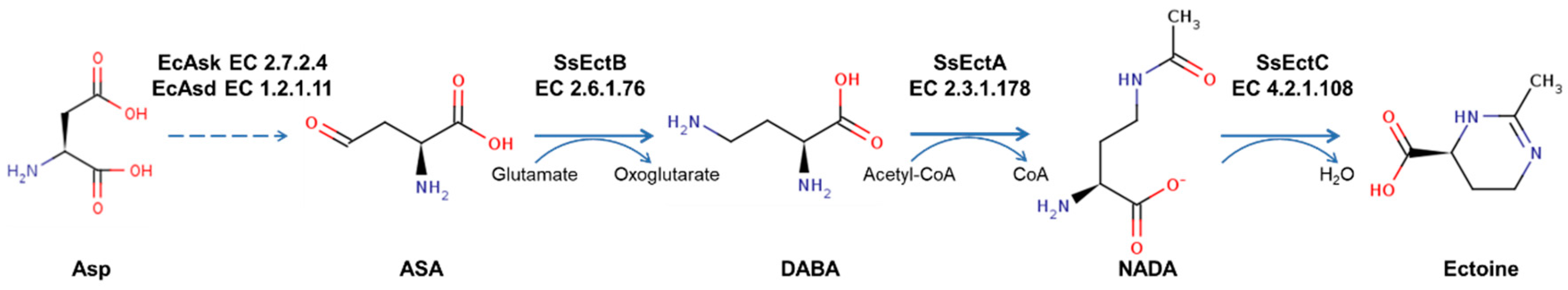
3.1. Gene Mining of Potential Ectoine Biosynthesis Cluster
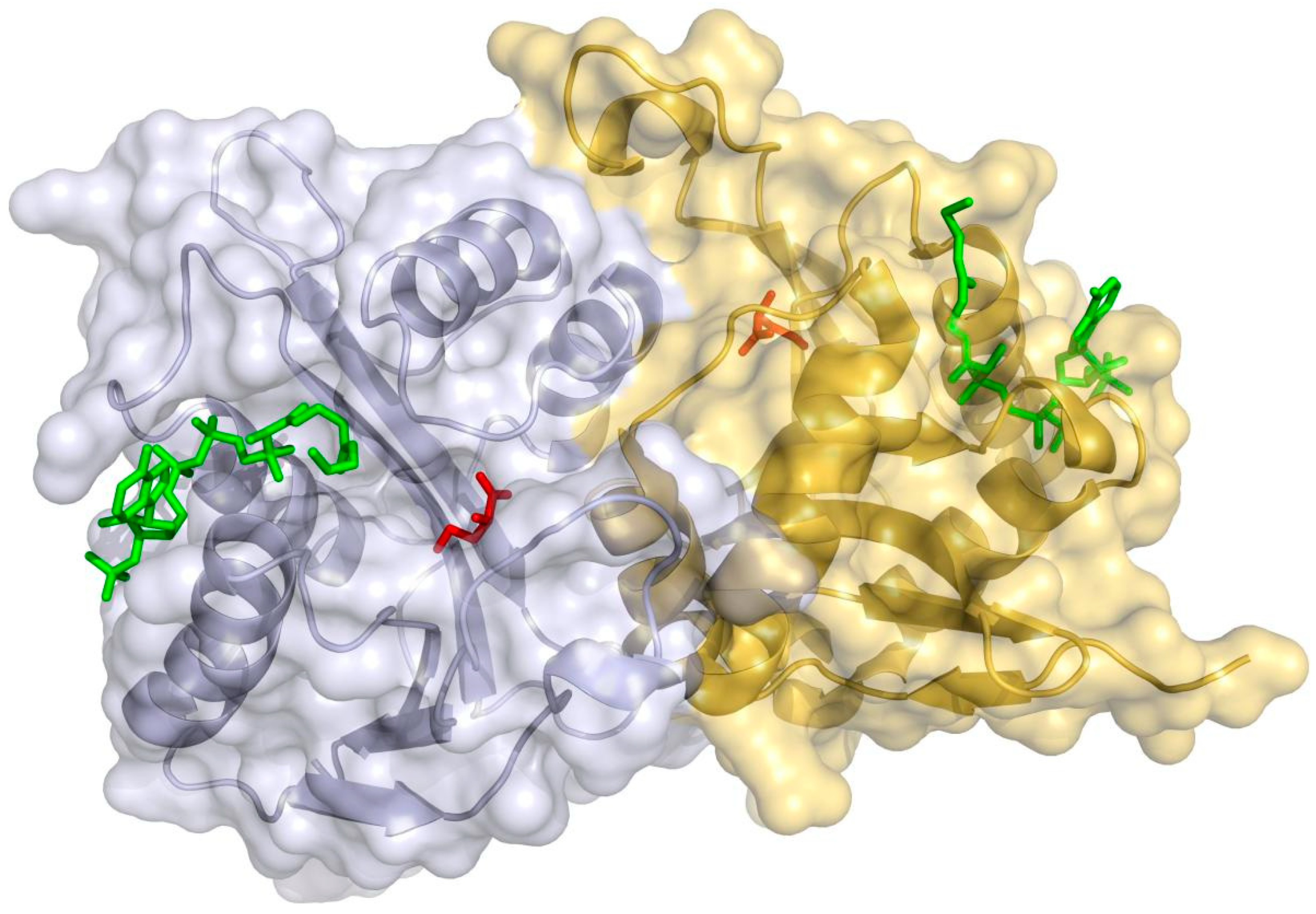
3.2. Bioinformatic Analysis and the Molecular Modeling of EctABCS. salarius
3.3. Production of Ectoine by the Recombinant E. coli
3.4. Optimization of Ectoine Production Conditions
3.5. Production of Ectoine Using High Density Cells in a Fermentor
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation or Terms | Meaning | Comments |
|---|---|---|
| Ask | L-aspartate-4-phosphotransferase | EC 2.7.2.4 |
| Asd | L-2, 4-diaminobutyrate:2-oxoglutarate-4-aminotransferase | EC 1.2.1.11 |
| EctB | Diaminobutyrate-2-oxoglutarate transaminase | EC 2.6.1.76 |
| EctA | L-2, 4-diaminobutyric acid acetyltransferase | EC 2.3.1.178 |
| EctC | L-ectoine synthase | EC 4.2.1.108 |
| ASA | L-aspartate-β-semialdehyde | Substrate of EctB |
| DABA | L-2, 4-diaminobutyric acid | Substrate of EctA |
| ADABA | Nγ-acetyl-L-2, 4-diaminobutyric acid | Substrate of EctC |
| CoA | Coenzyme A | |
| IPTG | Isopropyl-β-d-thiogalactoside | Inducer |
| Asp | Aspartate | Substrate of BL-SsEct |
| PLP | Pyridoxal-5-phosphate | Coenzyme for EctB |
| BL-SsEct | E. coli BL21(DE3)-pDK6- ectABCS. salarius | engineered strain |
| ectABCS. salarius | Ectoine biosynthetic cluster from Salinicola salarius 1A01339 | gene |
| ectABC | Ectoine biosynthetic gene cluster | gene |
References
- Oren, A.; Heldal, M.; Norland, S.; Galinski, E.A. Intracellular ion and organic solute concentrations of the extremely halophilic bacterium Salinibacter ruber. Extremophiles 2002, 6, 491–498. [Google Scholar] [CrossRef]
- Fenizia, S.; Thume, K.; Wirgenings, M.; Pohnert, G. Ectoine from Bacterial and Algal Origin is a Compatible Solute in Microalgae. Mar. Drugs 2020, 18, 42. [Google Scholar] [CrossRef] [Green Version]
- Galinski, E.A.; Pfeiffer, H.P.; Truper, H.G. 1,4,5,6-Tetrahydro-2-methyl-4-pyrimidinecarboxylic acid. A novel cyclic amino acid from halophilic phototrophic bacteria of the genus Ectothiorhodospira. Eur. J. Biochem. 1985, 149, 135–139. [Google Scholar] [CrossRef]
- Ono, H.; Sawada, K.; Khunajakr, N.; Tao, T.; Yamamoto, M.; Hiramoto, M.; Shinmyo, A.; Takano, M.; Murooka, Y. Characterization of biosynthetic enzymes for ectoine as a compatible solute in a moderately halophilic eubacterium, Halomonas elongata. J. Bacteriol. 1999, 181, 91–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, P.; Galinski, E.A. Characterization of genes for the biosynthesis of the compatible solute ectoine from Marinococcus halophilus and osmoregulated expression in Escherichia coli. Microbiology 1997, 143, 1141–1149. [Google Scholar] [CrossRef] [Green Version]
- Gregory, G.J.; Boyd, E.F. Stressed out: Bacterial response to high salinity using compatible solute biosynthesis and uptake systems, lessons from Vibrionaceae. Comput. Struct. Biotechnol. 2021, 19, 1014–1027. [Google Scholar] [CrossRef]
- Chen, J.; Liu, P.; Chu, X.; Chen, J.; Zhang, H.; Rowley, D.C.; Wang, H. Metabolic Pathway Construction and Optimization of Escherichia coli for High-Level Ectoine Production. Curr. Microbiol. 2020, 77, 1412–1418. [Google Scholar] [CrossRef] [PubMed]
- Bilstein, A.; Sonnemann, U. Nasal spray and eye drops containing ectoine, a novel natural, non-drug anti-allergic substance are not less effective than azelastine nasal spray and eye drops in improving the symptoms of allergic rhinitis and conjunctivitis. Allergy 2011, 66, 132. [Google Scholar] [CrossRef]
- Liu, M.; Liu, H.; Shi, M.; Jiang, M.; Li, L.; Zheng, Y. Microbial production of ectoine and hydroxyectoine as high-value chemicals. Microb. Cell. Fact. 2021, 20, 33771157. [Google Scholar] [CrossRef]
- Peters, P.; Galinski, E.; Trüper, H.G. The Biosynthesis of Ectoine. FEMS Microbiol. Lett. 1990, 71, 157–162. [Google Scholar] [CrossRef]
- Bursy, J.; Pierik, A.J.; Pica, N.; Bremer, E. Osmotically induced synthesis of the compatible solute hydroxyectoine is mediated by an evolutionarily conserved ectoine hydroxylase. J. Biol. Chem. 2007, 282, 31147–31155. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Zhao, Y.; Huang, W.; Zhang, L.; Wu, F.; Ye, J.; Chen, G.Q. Rational flux-tuning of Halomonas bluephagenesis for co-production of bioplastic PHB and ectoine. Nat. Commun. 2020, 11, 32620759. [Google Scholar] [CrossRef]
- Li, Y.; Wei, H.; Wang, T.; Xu, Q.; Zhang, C.; Fan, X.; Ma, Q.; Chen, N.; Xie, X. Current status on metabolic engineering for the production of l-aspartate family amino acids and derivatives. Bioresour. Technol. 2017, 245, 1588–1602. [Google Scholar] [CrossRef] [PubMed]
- Hermann, L.; Mais, C.-N.; Czech, L.; Smits, S.; Bange, G.; Bremer, E. The ups and downs of ectoine: Structural enzymology of a major microbial stress protectant and versatile nutrient. Biol. Chem. 2020, 401, 1443–1468. [Google Scholar] [CrossRef]
- Mais, C.N.; Hermann, L.; Altegoer, F.; Seubert, A.; Richter, A.A.; Wernersbach, I.; Czech, L.; Bremer, E.; Bange, G. Degradation of the microbial stress protectants and chemical chaperones ectoine and hydroxyectoine by a bacterial hydrolase-deacetylase complex. J. Biol. Chem. 2020, 295, 9087–9104. [Google Scholar] [CrossRef]
- Parwata, I.P.; Wahyuningrum, D.; Suhandono, S.; Hertadi, R. Heterologous Ectoine Production in Escherichia coli: Optimization Using Response Surface Methodology. Int. J. Microbiol. 2019, 2019, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hans Jorg, K.; Georg, L.; Erwin, A.G. Industrial Production of the Cell Protectant Ectoine: Protection Mechanisms, Processes, and Products. Curr. Biotech. 2014, 3, 10–25. [Google Scholar] [CrossRef] [Green Version]
- Vandrich, J.; Pfeiffer, F.; Alfaro-Espinoza, G.; Kunte, H.J. Contribution of mechanosensitive channels to osmoadaptation and ectoine excretion in Halomonas elongata. Extremophiles 2020, 24, 421–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.Z.; Gong, J.; Yu, H.Y.; Tao, Y.; Zhang, S.; Dong, Z.Y. High production of ectoine from aspartate and glycerol by use of whole-cell biocatalysis in recombinant Escherichia coli. Microb. Cell Fact. 2015, 14, 55. [Google Scholar] [CrossRef] [Green Version]
- Grammann, K.; Volke, A.; Kunte, H.J. New type of osmoregulated solute transporter identified in halophilic members of the Bacteria domain: TRAP transporter TeaABC mediates uptake of ectoine and hydroxyectoine in Halomonas elongata DSM 2581(T). J. Bacteriol. 2002, 184, 3078–3085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwibbert, K.; Marin-Sanguino, A.; Bagyan, I.; Heidrich, G.; Lentzen, G.; Seitz, H.; Rampp, M.; Schuster, S.C.; Klenk, H.P.; Pfeiffer, F.; et al. A blueprint of ectoine metabolism from the genome of the industrial producer Halomonas elongata DSM 2581(T). Environ. Microbiol. 2011, 13, 1973–1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauer, T.; Galinski, E.A. Bacterial milking: A novel bioprocess for production of compatible solutes. Biotechnol. Bioeng. 1998, 57, 306–313. [Google Scholar] [CrossRef]
- Zhu, D.; Liu, J.; Han, R.; Shen, G.; Long, Q.; Wei, X.; Liu, D. Identification and characterization of ectoine biosynthesis genes and heterologous expression of the ectABC gene cluster from Halomonas sp. QHL1, a moderately halophilic bacterium isolated from Qinghai Lake. J. Microbiol. 2014, 52, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Giesselmann, G.; Dietrich, D.; Jungmann, L.; Kohlstedt, M.; Jeon, E.J.; Yim, S.S.; Sommer, F.; Zimmer, D.; Muhlhaus, T.; Schroda, M.; et al. Metabolic Engineering of Corynebacterium glutamicum for High-Level Ectoine Production: Design, Combinatorial Assembly, and Implementation of a Transcriptionally Balanced Heterologous Ectoine Pathway. Biotechnol. J. 2019, 14, 1800417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Qin, N.; Guan, L.Y. A Novel Cold-adapted Endoglucanase (M6A) from Microbacterium kitamiense S12 Isolated from Qinghai-Tibetan Plateau. Biotechnol. Bioprocess Eng. 2019, 24, 544–551. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Richter, A.A.; Kobus, S.; Czech, L.; Hoeppner, A.; Zarzycki, J.; Erb, T.J.; Lauterbach, L.; Dickschat, J.S.; Bremer, E.; Smits, S.H.J. The architecture of the diaminobutyrate acetyltransferase active site provides mechanistic insight into the biosynthesis of the chemical chaperone ectoine. J. Biol. Chem. 2020, 295, 2822–2838. [Google Scholar] [CrossRef]
- Hillier, H.T.; Altermark, B.; Leiros, I. The crystal structure of the tetrameric DABA-aminotransferase EctB, a rate-limiting enzyme in the ectoine biosynthesis pathway. FEBS J. 2020, 287, 4641–4658. [Google Scholar] [CrossRef]
- Czech, L.; Hoppner, A.; Kobus, S.; Seubert, A.; Riclea, R.; Dickschat, J.S.; Heider, J.; Smits, S.H.J.; Bremer, E. Illuminating the catalytic core of ectoine synthase through structural and biochemical analysis. Sci. Rep. 2019, 9, 364. [Google Scholar] [CrossRef] [Green Version]
- Ning, Y.; Wu, X.; Zhang, C.; Xu, Q.; Chen, N.; Xie, X. Pathway construction and metabolic engineering for fermentative production of ectoine in Escherichia coli. Metab. Eng. 2016, 36, 10–18. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, 320–324. [Google Scholar] [CrossRef] [Green Version]
- Neuwald, A.F.; Landsman, D. GCN5-related histone N-acetyltransferases belong to a diverse superfamily that includes the yeast SPT10 protein. Trends Biochem. Sci. 1997, 22, 154–155. [Google Scholar] [CrossRef]
- Shen, B.W.; Hennig, M.; Hohenester, E.; Jansonius, J.N.; Schirmer, T. Crystal structure of human recombinant ornithine aminotransferase. J. Mol. Biol. 1998, 277, 81–102. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, A.U.; Bremer, E. Osmotically regulated synthesis of the compatible solute ectoine in Bacillus pasteurii and related Bacillus spp. Appl. Environ. Microbiol. 2002, 68, 772–783. [Google Scholar] [CrossRef] [Green Version]
- Czech, L.; Hermann, L.; Stoveken, N.; Richter, A.A.; Hoppner, A.; Smits, S.H.J.; Heider, J.; Bremer, E. Role of the Extremolytes Ectoine and Hydroxyectoine as Stress Protectants and Nutrients: Genetics, Phylogenomics, Biochemistry, and Structural Analysis. Genes 2018, 9, 177. [Google Scholar] [CrossRef] [Green Version]
- Schubert, T.; Maskow, T.; Benndorf, D.; Harms, H.; Breuer, U. Continuous synthesis and excretion of the compatible solute ectoine by a transgenic, nonhalophilic bacterium. Appl. Environ. Microbiol. 2007, 73, 3343–3347. [Google Scholar] [CrossRef] [Green Version]
- Margesin, R.; Schinner, F.; Marx, J.C.; Gerday, C. Psychrophiles: From Biodiversity to Biotechnology; Margesin, R., Ed.; Springer: Cham, Switzerland, 2017; pp. 285–303. [Google Scholar] [CrossRef]
- Dou, S.H.; Chi, N.Y.; Zhou, X.S.; Zhang, Q.F.; Pang, F.; Xiu, Z.L. Molecular cloning, expression, and biochemical characterization of a novel cold-active alpha-amylase from Bacillus sp. dsh19-1. Extremophiles 2018, 22, 739–749. [Google Scholar] [CrossRef]
- Merin, M.G.; de Ambrosini, V.I.M. Highly cold-active pectinases under wine-like conditions from non-Saccharomyces yeasts for enzymatic production during winemaking. Lett. Appl. Microbiol. 2015, 60, 467–474. [Google Scholar] [CrossRef]
- Bethlehem, L.; Moritz, K.D. Boosting Escherichia coli’s heterologous production rate of ectoines by exploiting the non-halophilic gene cluster from Acidiphilium cryptum. Extremophiles 2020, 24, 733–747. [Google Scholar] [CrossRef] [PubMed]
- Czech, L.; Wilcken, S.; Czech, O.; Linne, U.; Brauner, J.; Smits, S.H.J.; Galinski, E.A.; Bremer, E. Exploiting Substrate Promiscuity of Ectoine Hydroxylase for Regio- and Stereoselective Modification of Homoectoine. Front. Microbiol. 2019, 10, 2745–2762. [Google Scholar] [CrossRef] [PubMed]
- Richter, A.A.; Mais, C.N.; Czech, L.; Geyer, K.; Hoeppner, A.; Smits, S.H.J.; Erb, T.J.; Bange, G.; Bremer, E. Biosynthesis of the Stress-Protectant and Chemical Chaperon Ectoine: Biochemistry of the Transaminase EctB. Front. Microbiol. 2019, 10, 2811–2830. [Google Scholar] [CrossRef] [PubMed]









| Strain/Plasmid | Relevant Characteristics | Source |
|---|---|---|
| Gene | ||
| ectABCS. salarius | Ectoine biosynthesis cluster from Salinicola salarius 1A01339 | This study |
| Plasmid | ||
| pDK-6 | Kmr, tac promoter, cloning vector | Sangon Biotech |
| pDK6-ectABCS. salarius | Kmr, tac promoter, EctABCS. salarius expression plasmid | This study |
| Strain | ||
| E. coli DH5α | Cloning strain | Sangon Biotech |
| E. coli BL21(DE3) | Expression strain | Sangon Biotech |
| E. coli BL21(DE3)-pDK6 | Recombinant bacteria for control experiment | This study |
| E. coli BL21(DE3)-pDK6- ectABCS. salarius (BL-SsEct) | Engineered strain for producing ectoine | This study |
| Microorganism | Source of Genes | Ectoine Titer (g/L) | Optimal Temperature (°C) | Optimal pH | Reactor System | Reference |
|---|---|---|---|---|---|---|
| Corynebacterium glutamicum ectABCopt | Pseudomonas stutzeri | 65.3 | 30 | 7.0 | Fed-batch | [24] |
| Escherichia coli W3110 | Halomonas elongata | 25.1 | 36 | 7.0 | Fed-batch | [30] |
| Escherichia coli K12/BW25113 | Halomonas elongata DSM 2581 | 25.1 | 30 | 7.0 | Whole-cell catalysis | [19] |
| Escherichia coli BL21 (DE3) | Salinicola salarius 1A01339 | 22.5 | 25 | 6.5 | Fed-batch | This study |
| Escherichia coli MG1655 | Halomonas elongata | 12.7 | 40 | 6.5 | Batch | [7] |
| Escherichia coli DH5α | Chromohalobacter salexigens DSM 3034 | 6.0 | 30 | 7.0 | Batch | [37] |
| Escherichia coli DH5α | Acidiphilium cryptum DSM 2389 | 1.7 | 37 | 7.0 | Batch | [41] |
| E. coli BL21 (DE3) | Halomonas elongata BK-AG25 | 0.75 | 25 | 7.0 | Batch | [16] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Y.; Peng, W.; Wang, T.; Li, Y.; Zhao, L.; Wang, X.; Li, Y.; Lin, L. Ectoine Production Using Novel Heterologous EctABCS. salarius from Marine Bacterium Salinicola salarius. Appl. Sci. 2021, 11, 6873. https://doi.org/10.3390/app11156873
Su Y, Peng W, Wang T, Li Y, Zhao L, Wang X, Li Y, Lin L. Ectoine Production Using Novel Heterologous EctABCS. salarius from Marine Bacterium Salinicola salarius. Applied Sciences. 2021; 11(15):6873. https://doi.org/10.3390/app11156873
Chicago/Turabian StyleSu, Yue, Wenting Peng, Tong Wang, Yanhui Li, Luyu Zhao, Xinyu Wang, Ying Li, and Ling Lin. 2021. "Ectoine Production Using Novel Heterologous EctABCS. salarius from Marine Bacterium Salinicola salarius" Applied Sciences 11, no. 15: 6873. https://doi.org/10.3390/app11156873
APA StyleSu, Y., Peng, W., Wang, T., Li, Y., Zhao, L., Wang, X., Li, Y., & Lin, L. (2021). Ectoine Production Using Novel Heterologous EctABCS. salarius from Marine Bacterium Salinicola salarius. Applied Sciences, 11(15), 6873. https://doi.org/10.3390/app11156873