
Asynchrony Drives Plant and Animal Community Stability in Mediterranean Coastal Dunes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Collection Methods
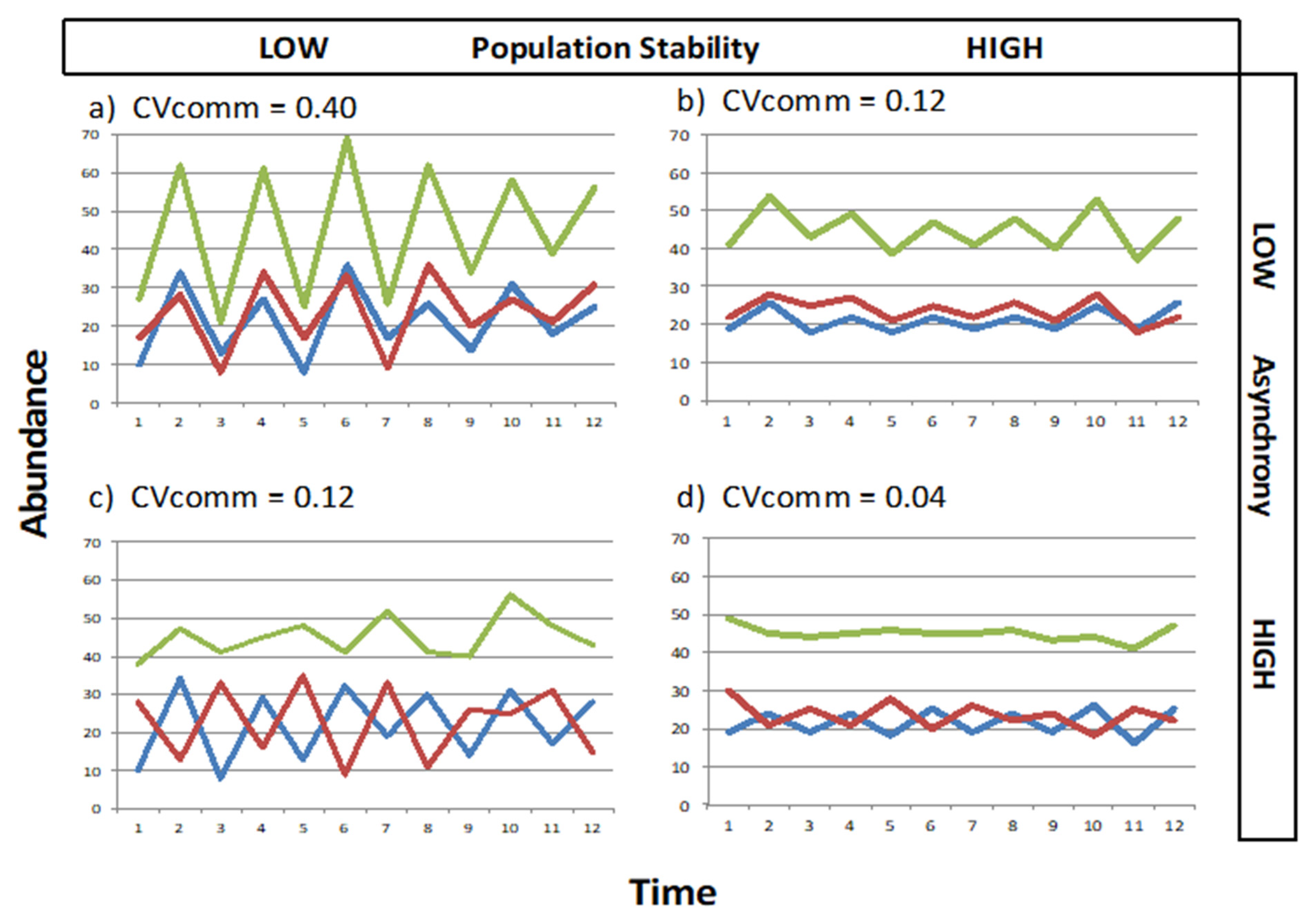
2.3. Measures of Stability
2.4. Statistical Analysis
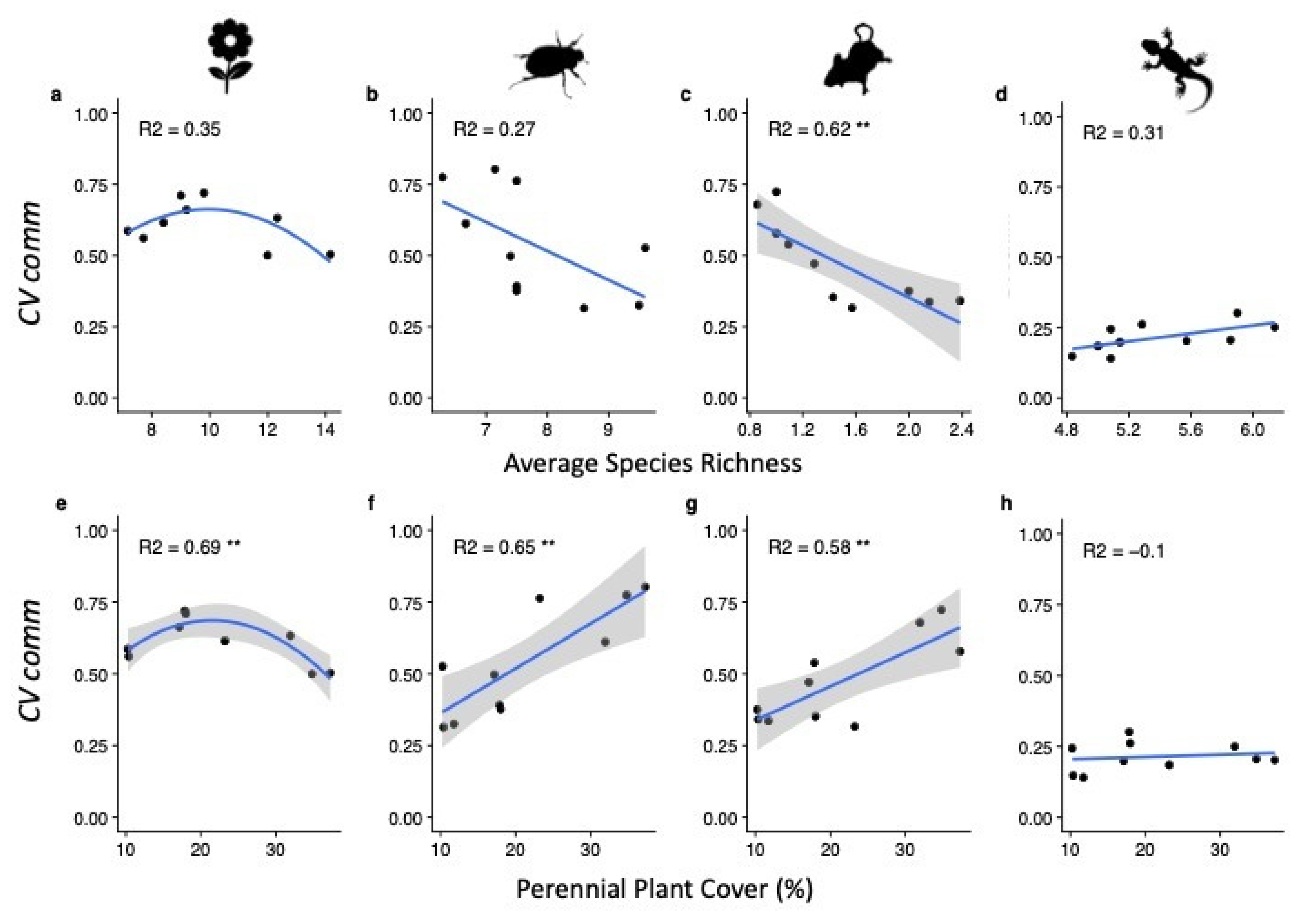
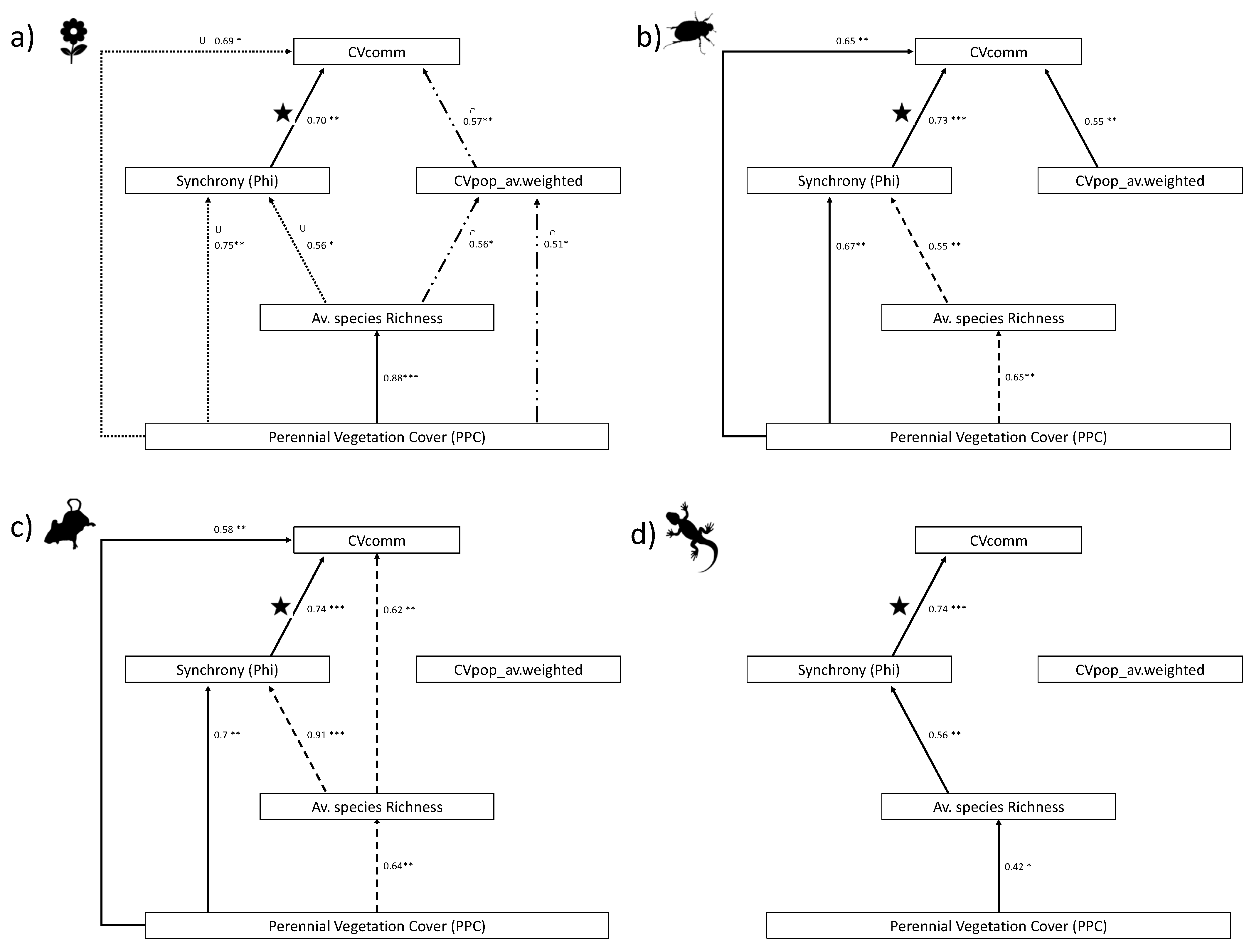
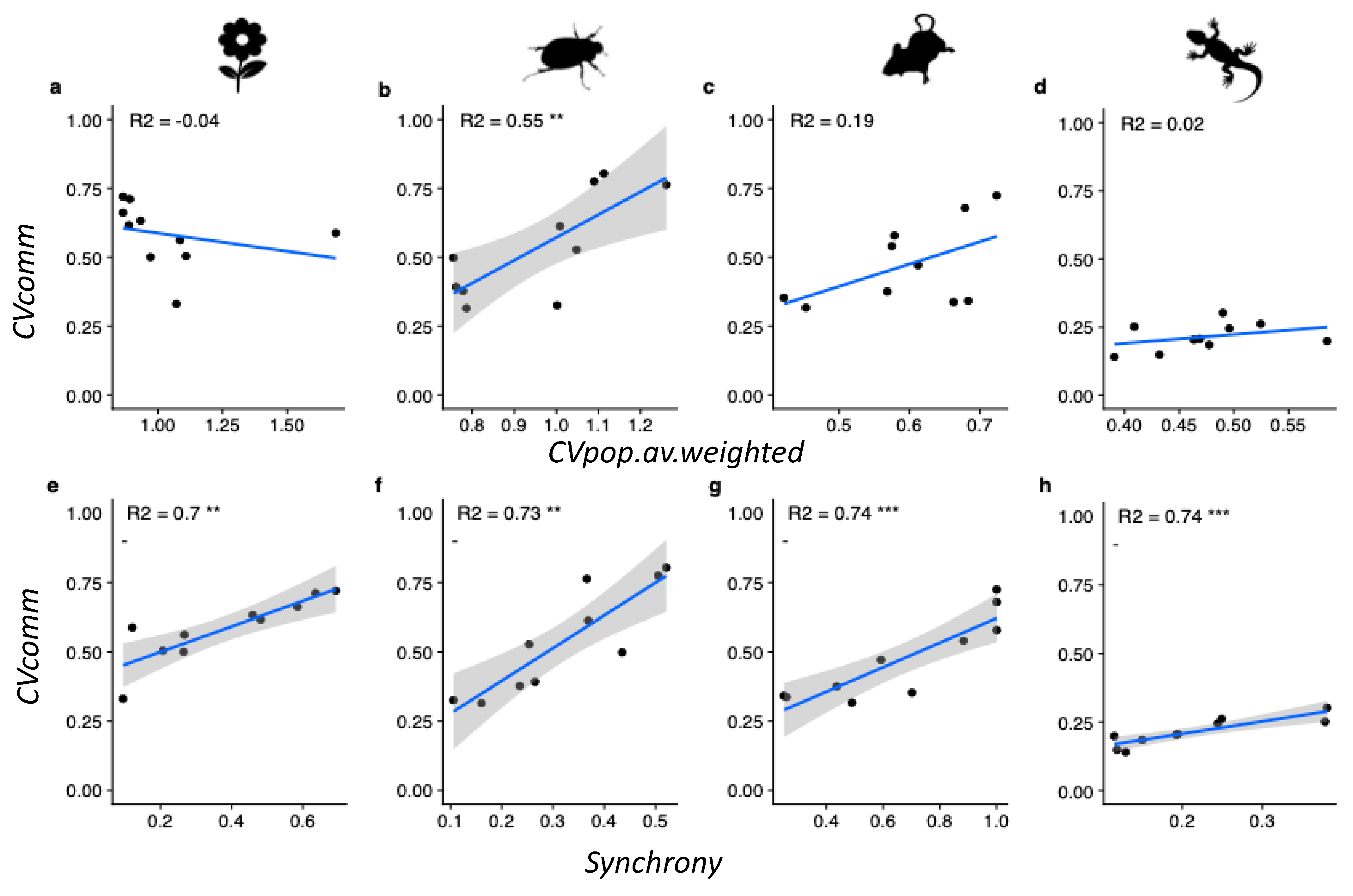
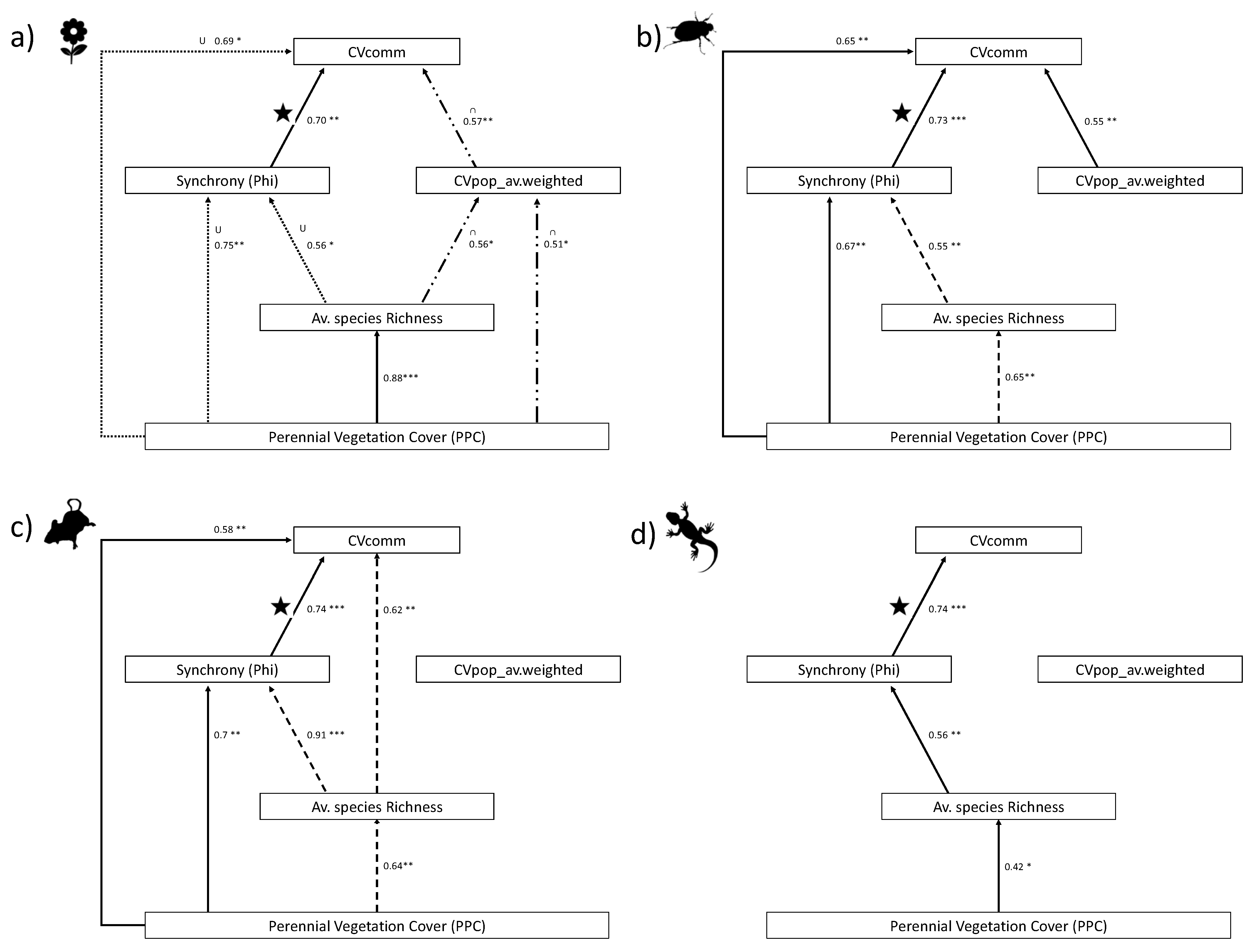
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pimm, S.L. The complexity and stability of ecosystems. Nature 1984, 307, 321–326. [Google Scholar] [CrossRef]
- Tilman, D.; Downing, J.A. Biodiversity and stability in grasslands. Nature 1994, 367, 363–365. [Google Scholar] [CrossRef]
- McCann, K.S. The diversity–stability debate. Nature 2000, 405, 228–233. [Google Scholar] [CrossRef]
- Loreau, M.; de Mazancourt, C. Biodiversity and ecosystem stability: A synthesis of underlying mechanisms. Ecol. Lett. 2013, 16, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Kéfi, S.; Domínguez-García, V.; Donohue, I.; Fontaine, C.; Thébault, E.; Dakos, V. Advancing our understanding of ecological stability. Ecol. Lett. 2019, 22, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Schindler, D.E.; Hilborn, R.; Chasco, B.; Boatright, C.P.; Quinn, T.P.; Rogers, L.; Webster, M.S. Population diversity and the portfolio effect in an exploited species. Nature 2010, 465, 609–612. [Google Scholar] [CrossRef] [PubMed]
- May, R.M. Stability and Complexity in Model Ecosystems; Princeton University Press: Princeton, NJ, USA, 2001; ISBN 9780691088617. [Google Scholar]
- Grman, E.; Lau, J.A.; Schoolmaster, D.R.; Gross, K.L. Mechanisms contributing to stability in ecosystem function depend on the environmental context. Ecol. Lett. 2010, 13, 1400–1410. [Google Scholar] [CrossRef] [PubMed]
- Naeem, S. Ecosystem consequences of biodiversity loss: The evolution of a paradigm. Ecology 2002, 83, 1537–1552. [Google Scholar] [CrossRef]
- Domínguez-García, V.; Dakos, V.; Kéfi, S. Unveiling dimensions of stability in complex ecological networks. Proc. Natl. Acad. Sci. USA 2019, 116, 25714–25720. [Google Scholar] [CrossRef]
- Thebault, E. Uncertain predictions of species responses to perturbations lead to underestimate changes at ecosystem level in diverse systems. Peer Community Ecol. 2020, 1, 100063. [Google Scholar] [CrossRef]
- Doak, D.F.; Bigger, D.; Harding, E.K.; Marvier, M.A.; O’Malley, R.E.; Thomson, D. The statistical inevitability of stability-diversity relationship in community ecology. Am. Nat. 1998, 151, 264–276. [Google Scholar] [CrossRef]
- Lehman, C.L.; Tilman, D. Biodiversity, stability, and productivity in competitive communities. Am. Nat. 2000, 156, 534–552. [Google Scholar] [CrossRef] [PubMed]
- Loreau, M.; de Mazancourt, C. Species Synchrony and Its Drivers: Neutral and Nonneutral Community Dynamics in Fluctuating Environments. Am. Nat. 2008, 172, E48–E66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilman, D.; Reich, P.B.; Knops, J.M.H. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 2006, 441, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Hector, A.; Hautier, Y.; Saner, P.; Wacker, L.; Bagchi, R.; Joshi, J.; Scherer-Lorenzen, M.; Spehn, E.M.; Bazeley-White, E.; Weilenmann, M.; et al. General stabilizing effects of plant diversity on grassland productivity through population asynchrony and overyielding. Ecology 2010, 91, 2213–2220. [Google Scholar] [CrossRef]
- Lhomme, J.-P.; Winkel, T. Diversity–stability relationships in community ecology: Re-examination of the Portfolio effect. Theor. Popul. Biol. 2002, 279, 271–279. [Google Scholar] [CrossRef]
- Thibaut, L.M.; Connolly, S.R. Understanding diversity-stability relationships: Towards a unified model of portfolio effects. Ecol. Lett. 2013, 16, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Mrowicki, R.J.; O’Connor, N.E.; Donohue, I. Temporal variability of a single population can determine the vulnerability of communities to perturbations. J. Ecol. 2016, 104, 887–897. [Google Scholar] [CrossRef] [Green Version]
- White, L.; O’Connor, N.E.; Yang, Q.; Emmerson, M.C.; Donohue, I. Individual species provide multifaceted contributions to the stability of ecosystems. Nat. Ecol. Evol. 2020, 4, 1594–1601. [Google Scholar] [CrossRef]
- Craven, D.; Eisenhauer, N.; Pearse, W.D.; Hautier, Y.; Isbell, F.; Roscher, C.; Bahn, M.; Beierkuhnlein, C.; Bönisch, G.; Buchmann, N.; et al. Multiple facets of biodiversity drive the diversity–stability relationship. Nat. Ecol. Evol. 2018, 2, 1579–1587. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Loreau, M. Ecosystem stability in space: Alpha, beta and gamma variability. Ecol. Lett. 2014, 17, 891–901. [Google Scholar] [CrossRef]
- Gouhier, T.C.; Guichard, F. Synchrony: Quantifying variability in space and time. Methods Ecol. Evol. 2014, 5, 524–533. [Google Scholar] [CrossRef]
- Morin, X.; Fahse, L.; de Mazancourt, C.; Scherer-Lorenzen, M.; Bugmann, H. Temporal stability in forest productivity increases with tree diversity due to asynchrony in species dynamics. Ecol. Lett. 2014, 17, 1526–1535. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Lu, X.; Hirota, M.; Bai, Y. Species asynchrony and response diversity determine multifunctional stability of natural grasslands. J. Ecol. 2019, 107, 1862–1875. [Google Scholar] [CrossRef]
- Sasaki, T.; Lauenroth, W.K. Dominant species, rather than diversity, regulates temporal stability of plant communities. Oecologia 2011, 166, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Blüthgen, N.; Simons, N.K.; Jung, K.; Prati, D.; Renner, S.C.; Boch, S.; Fischer, M.; Hölzel, N.; Klaus, V.H.; Kleinebecker, T.; et al. Land use imperils plant and animal community stability through changes in asynchrony rather than diversity. Nat. Commun. 2016, 7, 10697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maestre, F.T.; Castillo-Monroy, A.P.; Bowker, M.A.; Ochoa-Hueso, R. Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern. J. Ecol. 2012, 100, 317–330. [Google Scholar] [CrossRef]
- Tredennick, A.T.; de Mazancourt, C.; Loreau, M.; Adler, P.B. Environmental responses, not species interactions, determine synchrony of dominant species in semiarid grasslands. Ecology 2017, 98, 971–981. [Google Scholar] [CrossRef] [Green Version]
- Worm, B.; Duffy, J.E. Biodiversity, productivity and stability in real food webs. Trends Ecol. Evol. 2003, 18, 628–632. [Google Scholar] [CrossRef]
- Hillebrand, H.; Langenheder, S.; Lebret, K.; Lindström, E.; Östman, Ö.; Striebel, M. Decomposing multiple dimensions of stability in global change experiments. Ecol. Lett. 2018, 21, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Ives, A.R.; Carpenter, S.R. Stability and diversity of ecosystems. Science 2007, 317, 58–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brose, U.; Hillebrand, H. Biodiversity and ecosystem functioning in dynamic landscapes. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hautier, Y.; Seabloom, E.W.; Borer, E.T.; Adler, P.B.; Harpole, W.S.; Hillebrand, H.; Lind, E.M.; MacDougall, A.S.; Stevens, C.J.; Bakker, J.D.; et al. Eutrophication weakens stabilizing effects of diversity in natural grasslands. Nature 2014, 508, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Ren, H.; Li, M.-H.; van Ruijven, J.; Han, X.; Wan, S.; Li, H.; Yu, Q.; Jiang, Y.; Jiang, L. Environmental changes drive the temporal stability of semi-arid natural grasslands through altering species asynchrony. J. Ecol. 2015, 103, 1308–1316. [Google Scholar] [CrossRef]
- Hallett, L.M.; Hsu, J.S.; Cleland, E.E.; Collins, S.L.; Dickson, T.L.; Farrer, E.C.; Gherardi, L.A.; Gross, K.L.; Hobbs, R.J.; Turnbull, L.; et al. Biotic mechanisms of community stability shift along a precipitation gradient. Ecology 2014, 95, 1693–1700. [Google Scholar] [CrossRef] [Green Version]
- Seabloom, E.W. Compensation and the stability of restored grassland communities. Ecol. Appl. 2007, 17, 1876–1885. [Google Scholar] [CrossRef]
- Duan, M.; Liu, Y.; Yu, Z.; Baudry, J.; Li, L.; Wang, C.; Axmacher, J.C. Disentangling effects of abiotic factors and biotic interactions on cross-taxon congruence in species turnover patterns of plants, moths and beetles. Sci. Rep. 2016, 6, 23511. [Google Scholar] [CrossRef] [Green Version]
- Gossner, M.M.; Getzin, S.; Lange, M.; Pašalić, E.; Türke, M.; Wiegand, K.; Weisser, W.W. The importance of heterogeneity revisited from a multiscale and multitaxa approach. Biol. Conserv. 2013, 166, 212–220. [Google Scholar] [CrossRef]
- Rooney, R.C.; Azeria, E.T. The strength of cross-taxon congruence in species composition varies with the size of regional species pools and the intensity of human disturbance. J. Biogeogr. 2015, 42, 439–451. [Google Scholar] [CrossRef]
- Yong, D.L.; Barton, P.S.; Okada, S.; Crane, M.; Lindenmayer, D.B. Birds as surrogates for mammals and reptiles: Are patterns of cross-taxonomic associations stable over time in a human-modified landscape? Ecol. Indic. 2016, 69, 152–164. [Google Scholar] [CrossRef]
- Maes, D.; Bonte, D. Using distribution patterns of five threatened invertebrates in a highly fragmented dune landscape to develop a multispecies conservation approach. Biol. Conserv. 2006, 133, 490–499. [Google Scholar] [CrossRef]
- Flynn, D.F.B.; Gogol-Prokurat, M.; Nogeire, T.; Molinari, N.; Richers, B.T.; Lin, B.B.; Simpson, N.; Mayfield, M.M.; DeClerck, F. Loss of functional diversity under land use intensification across multiple taxa. Ecol. Lett. 2009, 12, 22–33. [Google Scholar] [CrossRef]
- Vihervaara, P.; D’Amato, D.; Forsius, M.; Angelstam, P.; Baessler, C.; Balvanera, P.; Boldgiv, B.; Bourgeron, P.; Dick, J.; Kanka, R.; et al. Using long-term ecosystem service and biodiversity data to study the impacts and adaptation options in response to climate change: Insights from the global ILTER sites network. Curr. Opin. Environ. Sustain. 2013, 5, 53–66. [Google Scholar] [CrossRef]
- Turner, M.G.; Collins, S.L.; Lugo, A.L.; Magnuson, J.J.; Rupp, T.S.; Swanson, F.J. Disturbance dynamics and ecological response: The contribution of Long-Term Ecological Research. Bioscience 2003, 53, 46. [Google Scholar] [CrossRef] [Green Version]
- Carboni, M.; Zelený, D.; Acosta, A.T.R. Measuring ecological specialization along a natural stress gradient using a set of complementary niche breadth indices. J. Veg. Sci. 2016, 27, 892–903. [Google Scholar] [CrossRef]
- Bonte, D.; Baert, L.; Maelfait, J.-P. Spider assemblage structure and stability in a heterogeneous coastal dune system (Belgium). J. Arachnol. 2002, 30, 331–343. [Google Scholar] [CrossRef]
- Ruocco, M.; Bertoni, D.; Sarti, G.; Ciccarelli, D. Mediterranean coastal dune systems: Which abiotic factors have the most influence on plant communities? Estuar. Coast. Shelf Sci. 2014, 149, 213–222. [Google Scholar] [CrossRef]
- Ciccarelli, D.; Bacaro, G.; Chiarucci, A. Coastline dune vegetation dynamics: Evidence of no stability. Folia Geobot. 2012, 47, 263–275. [Google Scholar] [CrossRef]
- Van Der Wurff, A.W.G.; Kools, S.A.E.; Boivin, M.E.Y.; Van Den Brink, P.J.; Van Megen, H.H.M.; Riksen, J.A.G.; Doroszuk, A.; Kammenga, J.E. Type of disturbance and ecological history determine structural stability. Ecol. Appl. 2007, 17, 190–202. [Google Scholar] [CrossRef]
- Isermann, M. Expansion of Rosa rugosa and Hippophae rhamnoides in coastal grey dunes: Effects at different spatial scales. Flora 2008, 203, 273–280. [Google Scholar] [CrossRef]
- Pye, K.; Blott, S.J.; Howe, M.A. Coastal dune stabilization in Wales and requirements for rejuvenation. J. Coast. Conserv. 2014, 18, 27–54. [Google Scholar] [CrossRef]
- Bird, T.L.F.; Dorman, M.; Ramot, A.; Bouskila, A.; Bar Kutiel, P.; Groner, E. Shrub encroachment effects on habitat heterogeneity and beetle diversity in a Mediterranean coastal dune system. Land Degrad. Dev. 2017, 28, 2553–2562. [Google Scholar] [CrossRef]
- Zuo, X.; Zhao, X.; Wang, S.; Li, Y.; Lian, J.; Zhou, X. Influence of dune stabilization on relationship between plant diversity and productivity in Horqin Sand Land, Northern China. Environ. Earth Sci. 2012, 67, 1547–1556. [Google Scholar] [CrossRef]
- Rosenzweig, M.L.; Abramsky, Z. How are diversity and productivity related? In Species Diversity in Ecological Communities: Historical and Geographical Perspectives; Ricklefs, R., Schluter, D., Eds.; University of Chicago Press: Chicago, IL, USA, 1993; pp. 52–65. [Google Scholar]
- Báez, S.; Collins, S.L. Shrub Invasion Decreases Diversity and Alters Community Stability in Northern Chihuahuan Desert Plant Communities. PLoS ONE 2008, 3, e2332. [Google Scholar] [CrossRef]
- Bird, T.L.F.; Bouskila, A.; Groner, E.; Bar Kutiel, P.; Kutiel, P.B. Can Vegetation Removal Successfully Restore Coastal Dune Biodiversity? Appl. Sci. 2020, 10, 2310. [Google Scholar] [CrossRef] [Green Version]
- Huston, M.A. Disturbance, productivity, and species diversity: Empiricism vs. logic in ecological theory. Ecology 2014, 95, 2382–2396. [Google Scholar] [CrossRef]
- Ferreira, S.M.; van Aarde, R.J. Maintaining diversity through intermediate disturbances: Evidence from rodents colonizing rehabilitating coastal dunes. Afr. J. Ecol. 2000, 38, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Kuiters, A.T.; Kramer, K.; Van der Hagen, H.G.J.M.; Schaminée, J.H.J. Plant diversity, species turnover and shifts in functional traits in coastal dune vegetation: Results from permanent plots over a 52-year period. J. Veg. Sci. 2009, 20, 1053–1063. [Google Scholar] [CrossRef]
- Isermann, M. Patterns in species diversity during succession of coastal dunes. J. Coast. Res. 2011, 274, 661–671. [Google Scholar]
- Kuiters, A.T. Diversity-stability relationships in plant communities of contrasting habitats. J. Veg. Sci. 2013, 24, 453–462. [Google Scholar] [CrossRef]
- Bar, P. Restoration of coastal sand dunes for conservation of biodiversity: The Israeli experience. In Restoration of Coastal Dunes; Springer Series on Environmental Management; Springer: Berlin/Heidelberg, Germany, 2013; pp. 173–185. [Google Scholar]
- Shacham, B. Dune Management and Reptiles—Implications for Habitat Reconstruction and Conservation Strategies. Ph.D. Thesis, Ben-Gurion University of the Negev, Be’er Sheva, Israel, 2010. [Google Scholar]
- Shacham, B.; Bouskila, A. Vegetation removal as a management tool in Nizzanim dunes (Israel): Preliminary assessment of effects on reptile and mammal populations. In Proceedings of the International Conference on Management and Restoration of Coastal Dunes (ICCD), Santander, Spain, 3–5 October 2007; pp. 144–145. [Google Scholar]
- Perry, M. Perennial Plants Impact on Annual Plant Diversity in Sand Dunes at Different Spatial Scales. Master’s Thesis, Ben Gurion University of the Negev, Be’er Sheva, Israel, 2008. [Google Scholar]
- Ramot, A. Effect of Plant Cover on Arthropod Community in Nizzanim Coastal Dunes. Master’s Thesis, Ben Gurion University of the Negev, Be’er Sheva, Israel, 2007. [Google Scholar]
- Levin, N.; Kidron, G.J.; Ben-Dor, E. A field quantification of coastal dune perennial plants as indicators of surface stability, erosion or deposition. Sedimentology 2008, 55, 751–772. [Google Scholar] [CrossRef]
- Kutiel, P.; Cohen, O.; Shoshany, M.; Shub, M. Vegetation establishment on the southern Israeli coastal sand dunes between the years 1965 and 1999. Landsc. Urban Plan. 2004, 67, 141–156. [Google Scholar] [CrossRef]
- Tsoar, H.; Blumberg, D.G. Formation of parabolic dunes from barchan and transverse dunes along Israel’s Mediterranean coast. Earth Surf. Process. Landf. 2002, 27, 1147–1161. [Google Scholar] [CrossRef]
- Kutiel, P.; Danin, A.; Orshan, G. Vegetation of the sandy soils near Caesarea, Israel. I. Plant communities, environment and succession. Isr. J. Bot. 1979, 28, 20–35. [Google Scholar]
- Kutiel, P. Annual vegetation of the coastal sand dunes of the northern Sharon, Israel. Isr. J. Plant Sci. 1998, 46, 287–298. [Google Scholar] [CrossRef]
- Kutiel, P. Conservation and management of the Mediterranean coastal sand dunes in Israel. J. Coast. Conserv. 2001, 7, 183–192. [Google Scholar] [CrossRef]
- Tsoar, H. Sand dunes mobility and stability in relation to climate. Phys. A Stat. Mech. Its Appl. 2005, 357, 50–56. [Google Scholar] [CrossRef]
- Rubinstein, Y.; Groner, E.; Yizhaq, H.; Svoray, T.; Bar (Kutiel), P. An eco-spatial index for evaluating stabilization state of sand dunes. Aeolian Res. 2013, 9, 75–87. [Google Scholar] [CrossRef]
- Fenu, G.; Carboni, M.; Acosta, A.T.R.; Bacchetta, G. Environmental factors influencing coastal vegetation pattern: New insights from the Mediterranean Basin. Folia Geobot. 2013, 48, 493–508. [Google Scholar] [CrossRef]
- Brittain, S.; Böhning, D. Estimators in capture–recapture studies with two sources. AStA Adv. Stat. Anal. 2009, 93, 23–47. [Google Scholar] [CrossRef]
- Carpenter, S.; Walker, B.; Anderies, J.M.; Abel, N. From metaphor to measurement: Resilience of what to what? Ecosystems 2001, 4, 765–781. [Google Scholar] [CrossRef]
- Cottingham, K.L.; Brown, B.L.; Lennon, J.T. Biodiversity may regulate the temporal variability of ecological systems. Ecol. Lett. 2001, 4, 72–85. [Google Scholar] [CrossRef]
- Elmqvist, T.; Folke, C.; Nyström, M.; Peterson, G.; Bengtsson, J.; Walker, B.; Norberg, J. Response diversity, ecosystem change, and resilience. Front. Ecol. Environ. 2003, 1, 488–494. [Google Scholar] [CrossRef]
- Tilman, D. Biodiversity: Population versus ecosystem stability. Ecology 1995, 77, 350–363. [Google Scholar] [CrossRef]
- Lepš, J. Variability in population and community biomass in a grassland community affected by environmental productivity and diversity. Oikos 2004, 107, 64–71. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: https://www.r-project.org/ (accessed on 10 October 2018).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- de Mazancourt, C.; Isbell, F.; Larocque, A.; Berendse, F.; De Luca, E.; Grace, J.B.; Haegeman, B.; Wayne Polley, H.; Roscher, C.; Schmid, B.; et al. Predicting ecosystem stability from community composition and biodiversity. Ecol. Lett. 2013, 16, 617–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilcox, K.R.; Tredennick, A.T.; Koerner, S.E.; Grman, E.; Hallett, L.M.; Avolio, M.L.; La Pierre, K.J.; Houseman, G.R.; Isbell, F.; Johnson, D.S.; et al. Asynchrony among local communities stabilises ecosystem function of metacommunities. Ecol. Lett. 2017, 20, 1534–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Jiang, L.; Li, L.; Li, A.; Wu, M.; Wan, S. Diversity-dependent stability under mowing and nutrient addition: Evidence from a 7-year grassland experiment. Ecol. Lett. 2012, 15, 619–626. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R. Fluctuations of animal populations and a measure of community stability. Ecology 1955, 36, 533–536. [Google Scholar] [CrossRef]
- Elton, C.S. The Ecology of Invasions by Plants and Animals; Methuen: London, UK, 1958. [Google Scholar]
- Hautier, Y.; Tilman, D.; Isbell, F.; Seabloom, E.W.; Borer, E.T.; Reich, P.B. Anthropogenic environmental changes affect ecosystem stability via biodiversity. Science 2015, 348, 336–340. [Google Scholar] [CrossRef] [Green Version]
- Isbell, F.I.; Polley, H.W.; Wilsey, B.J. Biodiversity, productivity and the temporal stability of productivity: Patterns and processes. Ecol. Lett. 2009, 12, 443–451. [Google Scholar] [CrossRef] [Green Version]
- Collins, S.L. Disturbance frequency and community stability in native tallgrass prairie. Am. Nat. 2000, 155, 311–325. [Google Scholar] [CrossRef]
- Donohue, I.; Hillebrand, H.; Montoya, J.M.; Petchey, O.L.; Pimm, S.L.; Fowler, M.S.; Healy, K.; Jackson, A.L.; Lurgi, M.; McClean, D.; et al. Navigating the complexity of ecological stability. Ecol. Lett. 2016, 19, 1172–1185. [Google Scholar] [CrossRef] [PubMed]
- Hovick, T.J.; Elmore, R.D.; Fuhlendorf, S.D.; Engle, D.M.; Hamilton, R.G. Spatial heterogeneity increases diversity and stability in grassland bird communities. Ecol. Appl. 2015, 25, 662–672. [Google Scholar] [CrossRef]
- Oberdorff, T.; Hugueny, B.; Vigneron, T. Is assemblage variability related to environmental variability? An answer for riverine fish. Oikos 2001, 93, 419–428. [Google Scholar] [CrossRef]
- Ziv, Y.; Smallwood, J.A. Gerbils and Heteromyids—Interspecific Competition and the Spatio-Temporal Niche. In Activity Patterns in Small Mammals; Springer: Berlin/Heidelberg, Germany, 2000; pp. 159–176. [Google Scholar]
- Kotler, B.P.; Brown, J.S.; Subach, A. Mechanisms of Species Coexistence of Optimal Foragers: Temporal Partitioning by Two Species of Sand Dune Gerbils. Oikos 1993, 67, 548. [Google Scholar] [CrossRef]
- Campbell, V.; Murphy, G.; Romanuk, T.N. Experimental design and the outcome and interpretation of diversity-stability relations. Oikos 2011, 120, 399–408. [Google Scholar] [CrossRef]
- Anderson, K.J. Temporal patterns in rates of community change during succession. Am. Nat. 2007, 169, 780–793. [Google Scholar] [CrossRef]
- Connor, E.F.; Simberloff, D. The Assembly of Species Communities: Chance or Competition? Ecology 1979, 60, 1132. [Google Scholar] [CrossRef]
- Cornell, H.V.; Lawton, J.H. Species Interactions, Local and Regional Processes, and Limits to the Richness of Ecological Communities: A Theoretical Perspective. J. Anim. Ecol. 1992, 61, 1. [Google Scholar] [CrossRef] [Green Version]
- May, R.M. Will a large complex system be stable? Nature 1972, 238, 413–414. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D. The ecological consequences of changes in biodiversity: A search for general principles. Ecology 1999, 80, 1455–1474. [Google Scholar] [CrossRef]
- Harvey, E.; Gounand, I.; Ward, C.L.; Altermatt, F. Bridging ecology and conservation: From ecological networks to ecosystem function. J. Appl. Ecol. 2017, 54, 371–379. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bird, T.L.F.; Bar, P.; Groner, E.; Bouskila, A. Asynchrony Drives Plant and Animal Community Stability in Mediterranean Coastal Dunes. Appl. Sci. 2021, 11, 6214. https://doi.org/10.3390/app11136214
Bird TLF, Bar P, Groner E, Bouskila A. Asynchrony Drives Plant and Animal Community Stability in Mediterranean Coastal Dunes. Applied Sciences. 2021; 11(13):6214. https://doi.org/10.3390/app11136214
Chicago/Turabian StyleBird, Tania L.F., Pua Bar (Kutiel), Elli Groner, and Amos Bouskila. 2021. "Asynchrony Drives Plant and Animal Community Stability in Mediterranean Coastal Dunes" Applied Sciences 11, no. 13: 6214. https://doi.org/10.3390/app11136214
APA StyleBird, T. L. F., Bar, P., Groner, E., & Bouskila, A. (2021). Asynchrony Drives Plant and Animal Community Stability in Mediterranean Coastal Dunes. Applied Sciences, 11(13), 6214. https://doi.org/10.3390/app11136214