
Comparison of the Potency of Pterostilbene with NF-κB Inhibitors in Platelet Activation: Mutual Activation by Akt-NF-κB Signaling in Human Platelets

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Platelet Preparation and Aggregation Study
2.3. Immunoblotting
2.4. Confocal Laser Fluorescence Microscopy
2.5. Acute Pulmonary Thromboembolism in Mice
2.6. Data Analysis
3. Results
3.1. BAY11-7082, Ro106-9920, and PTE Inhibit Collagen-Induced Platelet Aggregation
3.2. PTE Suppressed NF-κB Signals in Collagen-Activated Platelets
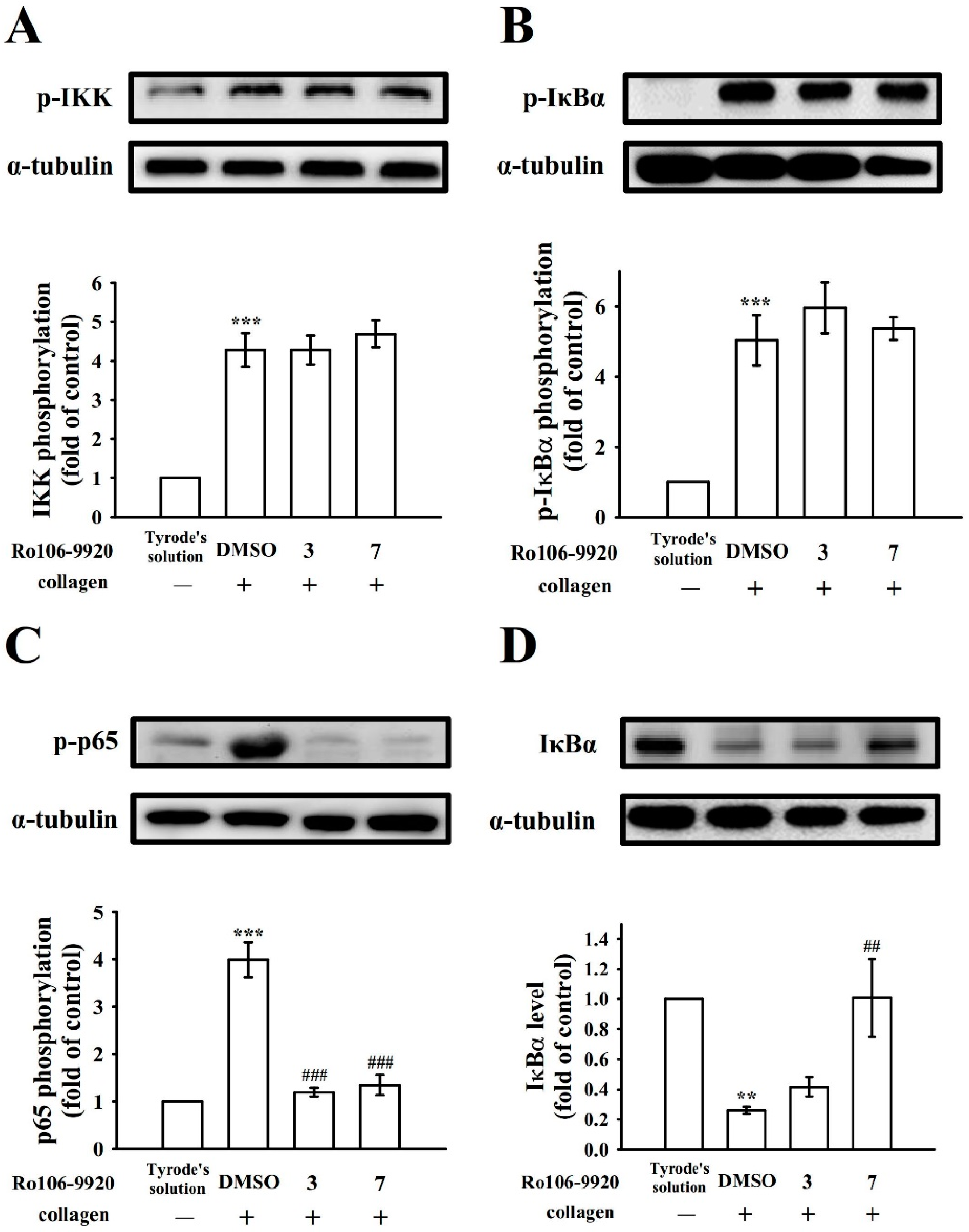
3.3. Relative Activity of BAY11-7082 and Ro106-9920 in NF-κB Activation
3.4. Effect of PTE, BAY11-7082, and Ro106-9920 in Immunostaining of p65 Phosphorylation by Confocal Microscopy
3.5. Signal Crosstalk Between Akt and NF-κB
3.6. Comparison of the Potency of PTE and BAY11-7082 in Reducing Acute Pulmonary Thrombosis Mortality
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kojok, K.; El-Kadiry, A.E.; Merhi, Y. Role of NF-κB in platelet function. Int. J. Mol. Sci. 2019, 20, 4185. [Google Scholar] [CrossRef]
- Cheng, H.S.; Sivachandran, N.; Lau, A.; Boudreau, E.; Zhao, J.L.; Baltimore, D.; Delgado-Olguin, P.; Cybulsky, M.I.; Fish, J.E. MicroRNA-146 represses endothelial activation by inhibiting pro-inflammatory pathways. EMBO Mol. Med. 2013, 5, 949–966. [Google Scholar] [CrossRef]
- Gawaz, M.; Langer, H.; May, A.E. May platelets in inflammation and atherogenesis. J. Clin. Investig. 2005, 115, 3378–3384. [Google Scholar] [CrossRef]
- Penz, S.; Reininger, A.J.; Brandl, R.; Goyal, P.; Rabie, T.; Bernlochner, I.; Rother, E.; Goetz, C.; Engelmann, B.; Smethurst, P.A.; et al. Human atheromatous plaques stimulate thrombus formation by activating platelet glycoprotein VI. FASEB J. 2005, 19, 898–909. [Google Scholar] [CrossRef]
- Tedgui, A.; Mallat, Z. Cytokines in atherosclerosis: Pathogenic and regulatory pathways. Physiol. Rev. 2006, 86, 515–581. [Google Scholar] [CrossRef]
- Monaco, C.; Andreakos, E.; Kiriakidis, S.; Mauri, C.; Bicknell, C.; Foxwell, B.; Cheshire, N.; Paleolog, E.; Feldmann, M. Canonical pathway of nuclear factor kappa B activation selectively regulates proinflammatory and prothrombotic responses in human atherosclerosis. Proc. Natl. Acad. Sci. USA 2004, 101, 5634–5639. [Google Scholar] [CrossRef]
- Fuentes, E.; Rojas, A.; Palomo, I. NF-κB signaling pathway as target for antiplatelet activity. Blood Rev. 2016, 30, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Karim, Z.A.; Zhang, J.; Banerjee, M.; Chicka, M.C.; Al Hawas, R.; Hamilton, T.R.; Roche, P.A.; Whiteheart, S.W. IkappaB kinase phosphorylation of SNAP-23 controls platelet secretion. Blood 2013, 121, 4567–4574. [Google Scholar] [CrossRef] [PubMed]
- Rivadeneyra, L.; Carestia, A.; Etulain, J.; Pozner, R.G.; Fondevila, C.; Negrotto, S.; Schattner, M. Regulation of platelet responses triggered by Toll-like receptor 2 and 4 ligands is another non-genomic role of nuclear factor-kappaB. Thromb. Res. 2014, 133, 235–243. [Google Scholar] [CrossRef]
- Jackson, S.P.; Yap, C.L.; Anderson, K.E. Phosphoinositide 3-kinases and the regulation of platelet function. Biochem. Soc. Trans. 2004, 32, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Mangin, P.; Dangelmaier, C.; Lillian, R.; Jackson, S.P.; Daniel, J.L.; Kunapuli, S.P. Role of phosphoinositide 3-kinase beta in glycoprotein VI-mediated Akt activation in platelets. J. Biol. Chem. 2009, 284, 33763–33772. [Google Scholar] [CrossRef]
- Xiang, B.; Zhang, G.; Liu, J.; Morris, A.J.; Smyth, S.S.; Gartner, T.K.; Li, Z. A G(i)-independent mechanism mediating Akt phosphorylation in platelets. J. Thromb. Haemost. 2010, 8, 2032–2041. [Google Scholar] [CrossRef]
- Dvorakova, M.; Landa, P. Anti-inflammatory activity of natural stilbenoids: A review. Pharmacol. Res. 2017, 124, 126–145. [Google Scholar] [CrossRef]
- Zhu, Q.; Tang, T.; Liu, H.; Sun, Y.; Wang, X.; Liu, Q.; Yang, L.; Lei, Z.; Huang, Z.; Chen, Z.; et al. Pterostilbene attenuates cocultured BV-2 microglial inflammation-mediated SH-SY5Y neuronal oxidative injury via SIRT-1 signaling. Oxid. Med. Cell. Longev. 2020, 2020, 3986348. [Google Scholar] [CrossRef]
- Park, E.S.; Lim, Y.; Hong, J.T.; Yoo, H.S.; Lee, C.K.; Pyo, M.Y.; Yun, Y.P. Pterostilbene, a natural dimethylated analog of resveratrol, inhibits rat aortic vascular smooth muscle cell proliferation by blocking Akt-dependent pathway. Vascular Pharmacol. 2010, 53, 61–67. [Google Scholar] [CrossRef]
- Huang, W.C.; Liu, J.C.; Hsia, C.W.; Fong, T.H.; Hsia, C.H.; Tran, O.T.; Velusamy, M.; Yang, C.H.; Sheu, J.R. Pterostilbene, a dimethylether analogue of resveratrol, possesses high potency in the prevention of platelet activation in humans and the reduction of vascular thrombosis in mice. J. Agric. Food Chem. 2021, 69, 4697–4707. [Google Scholar] [CrossRef]
- Huang, W.C.; Lin, K.C.; Hsia, C.W.; Hsia, C.H.; Chen, T.Y.; Bhavan, P.S.; Sheu, J.R.; Hou, S.M. The antithrombotic agent pterostilbene interferes with integrin αIIbβ3-mediated inside-out and outside-in signals in human platelets. Int. J. Mol. Sci. 2021, 22, 3643. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.F.; Lee, J.J.; Chang, C.C.; Lin, K.H.; Wang, S.H.; Sheu, J.R. Platelet protease-activated receptor (PAR)4, but not PAR1, associated with neutral sphingomyelinase responsible for thrombin-stimulated ceramide-NF-κB signaling in human platelets. Haematologica 2013, 98, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Crosby, D.; Poole, A.W. Physical and functional interaction between protein kinase Cγ and Fyn tyrosine kinase in human platelets. J. Biol. Chem. 2003, 278, 24533–24541. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.R.; Hung, W.C.; Wu, C.H.; Lee, Y.M.; Yen, M.H. Antithrombotic effect of rutaecarpine, an alkaloid isolated from Evodia rutaecarpa, on platelet plug formation in in vivo experiments. Br. J. Haematol. 2000, 110, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Hsia, C.W.; Wu, M.P.; Shen, M.Y.; Hsia, C.H.; Chung, C.L.; Sheu, J.R. Regulation of human platelet activation and prevention of arterial thrombosis in mice by auraptene through inhibition of NF-κB pathway. Int. J. Mol. Sci. 2020, 21, 4810. [Google Scholar] [CrossRef]
- Lin, Y.; Bai, L.; Chen, W.; Xu, S. The NF-kappaB activation pathways, emerging molecular targets for cancer prevention and therapy. Expert Opin. Ther. Targets 2010, 14, 45–55. [Google Scholar] [CrossRef]
- Swinney, D.C.; Xu, Y.Z.; Scarafia, L.E.; Lee, I.; Mak, A.Y.; Gan, Q.F.; Ramesha, C.S.; Mulkins, M.A.; Dunn, J.; So, O.Y.; et al. A small molecule ubiquitination inhibitor blocks NF-kappa B-dependent cytokine expression in cells and rats. J. Biol. Chem. 2002, 277, 23573–23581. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Hayden, M.S. New regulators of NF-kappaB in inflammation. Nat. Rev. Immunol. 2008, 8, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Manning, B.D.; Cantley, L.C. AKT/PKB signaling: Navigating downstream. Cell 2007, 129, 1261–1274. [Google Scholar] [CrossRef]
- Fulda, S.; Debatin, K.M. Resveratrol modulation of signal transduction in apoptosis and cell survival: A mini-review. Cancer Detect. Prev. 2006, 30, 217–223. [Google Scholar] [CrossRef]
- Lin, H.S.; Yue, B.D.; Ho, P.C. Determination of pterostilbene in rat plasma by a simple HPLC-UV method and its application in pre-clinical pharmacokinetic study. Biomed. Chromatogr. 2009, 23, 1308–1315. [Google Scholar] [CrossRef]
- Kapetanovic, I.M.; Muzzio, M.; Huang, Z.; Thompson, T.N.; McCormick, D.L. Pharmacokinetics, oral bioavailability, and metabolic profile of resveratrol and its dimethylether analog, pterostilbene, in rats. Cancer Chemother. Pharmacol. 2011, 68, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.Y.; Davidson, S.J.; Moraes, L.A.; Traves, S.L.; Paul-Clark, M.; Bishop-Bailey, D.; Warner, T.D.; Mitchell, J.A. Role of nuclear receptor signaling in platelets: Antithrombotic effects of PPARbeta. FASEB J. 2006, 20, 326–328. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, L.M.; Freedman, J.E. NFkappaB regulation of platelet function: No nucleus, no genes, no problem? J. Thromb. Haemost. 2009, 7, 1329–1332. [Google Scholar] [CrossRef][Green Version]
- Malaver, E.; Romaniuk, M.A.; D’Atri, L.P.; Pozner, R.G.; Negrotto, S.; Benzadon, R.; Schattner, M. NF-kappaB inhibitors impair platelet activation responses. J. Thromb. Haemost. 2009, 7, 1333–1343. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; De, S.; Damron, D.S.; Chen, W.S.; Hay, N.; Byzova, T.V. Impaired platelet responses to thrombin and collagen in AKT-1-deficient mice. Blood 2004, 104, 1703–1710. [Google Scholar] [CrossRef] [PubMed]
- Stojanovic, A.; Marjanovic, J.A.; Brovkovych, V.M.; Peng, X.; Hay, N.; Skidgel, R.A.; Du, X. A phosphoinositide 3-kinase-AKT-nitric oxide-cGMP signaling pathway in stimulating platelet secretion and aggregation. J. Biol. Chem. 2006, 281, 16333–16339. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.A.; Stojanovic-Terpo, A.; Hay, N.; Du, X. An important role for Akt3 in platelet activation and thrombosis. Blood 2011, 118, 4215–4223. [Google Scholar] [CrossRef]
- Moore, S.F.; van den Bosch, M.T.; Hunter, R.W.; Sakamoto, K.; Poole, A.W.; Hers, I. Dual regulation of glycogen synthase kinase 3 (GSK3)alpha/beta by protein kinase C (PKC)alpha and Akt promotes thrombin-mediated integrin alphaIIbbeta3 activation and granule secretion in platelet. J. Biol. Chem. 2013, 288, 3918–3928. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Number (n) | Number of Deaths (n) | Mortality (%) | |
|---|---|---|---|
| ADP (700 mg/kg) | |||
| + solvent control (0.1% DMSO) | 12 | 12 | 100 |
| + PTE (2 mg/kg) | 12 | 4 | 33 |
| + BAY11-7082 (2 mg/kg) | 12 | 8 | 67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsia, C.-W.; Huang, W.-C.; Yang, C.-H.; Hsia, C.-H.; Jayakumar, T.; Bhavan, P.S.; Sheu, J.-R.; Chiou, K.-R. Comparison of the Potency of Pterostilbene with NF-κB Inhibitors in Platelet Activation: Mutual Activation by Akt-NF-κB Signaling in Human Platelets. Appl. Sci. 2021, 11, 6149. https://doi.org/10.3390/app11136149
Hsia C-W, Huang W-C, Yang C-H, Hsia C-H, Jayakumar T, Bhavan PS, Sheu J-R, Chiou K-R. Comparison of the Potency of Pterostilbene with NF-κB Inhibitors in Platelet Activation: Mutual Activation by Akt-NF-κB Signaling in Human Platelets. Applied Sciences. 2021; 11(13):6149. https://doi.org/10.3390/app11136149
Chicago/Turabian StyleHsia, Chih-Wei, Wei-Chieh Huang, Chih-Hao Yang, Chih-Hsuan Hsia, Thanasekaran Jayakumar, Periyakali Saravana Bhavan, Joen-Rong Sheu, and Kuan-Rau Chiou. 2021. "Comparison of the Potency of Pterostilbene with NF-κB Inhibitors in Platelet Activation: Mutual Activation by Akt-NF-κB Signaling in Human Platelets" Applied Sciences 11, no. 13: 6149. https://doi.org/10.3390/app11136149
APA StyleHsia, C.-W., Huang, W.-C., Yang, C.-H., Hsia, C.-H., Jayakumar, T., Bhavan, P. S., Sheu, J.-R., & Chiou, K.-R. (2021). Comparison of the Potency of Pterostilbene with NF-κB Inhibitors in Platelet Activation: Mutual Activation by Akt-NF-κB Signaling in Human Platelets. Applied Sciences, 11(13), 6149. https://doi.org/10.3390/app11136149