
Hypoxic Exercise Training Elevates Erythrocyte Aggregation

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
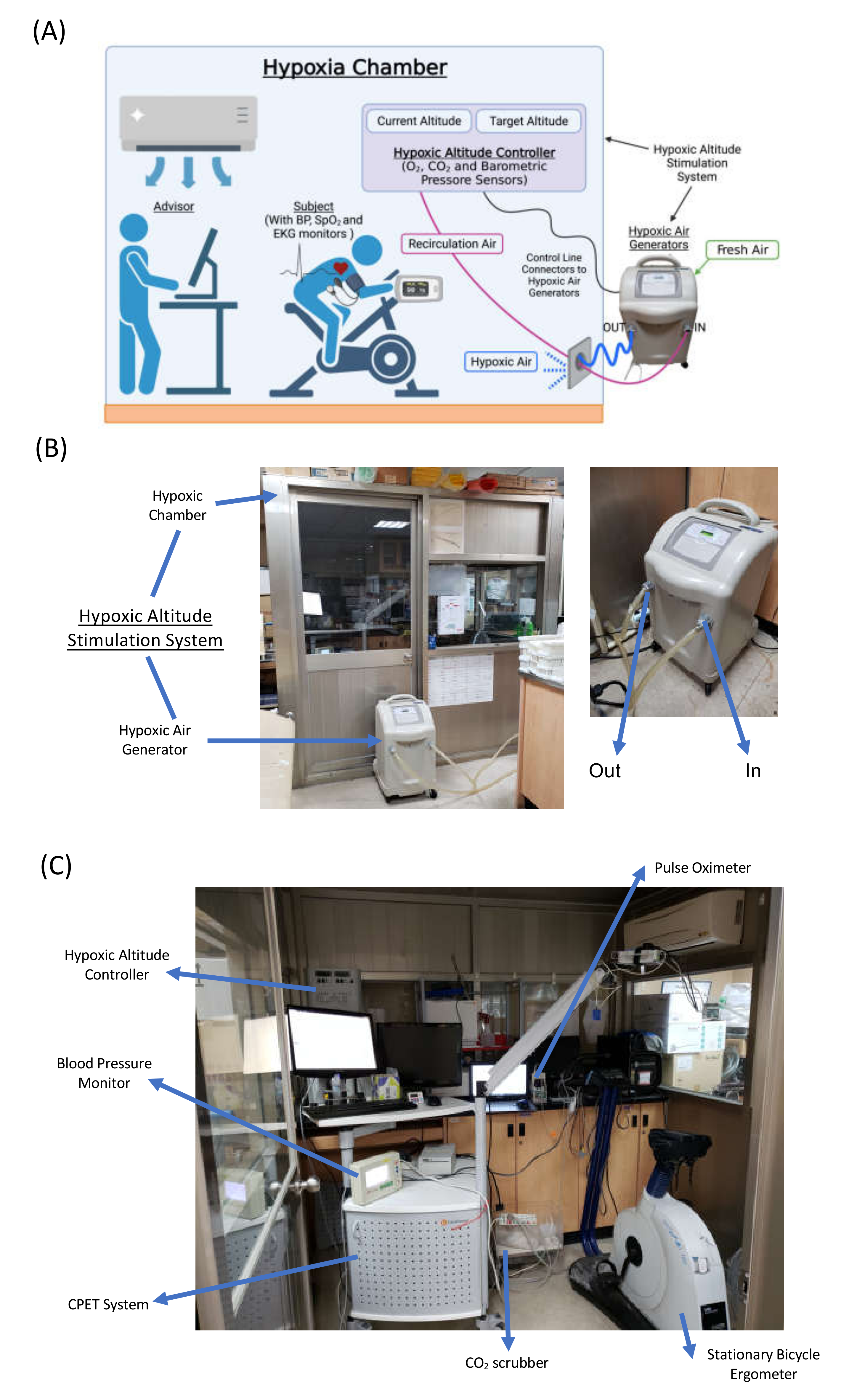
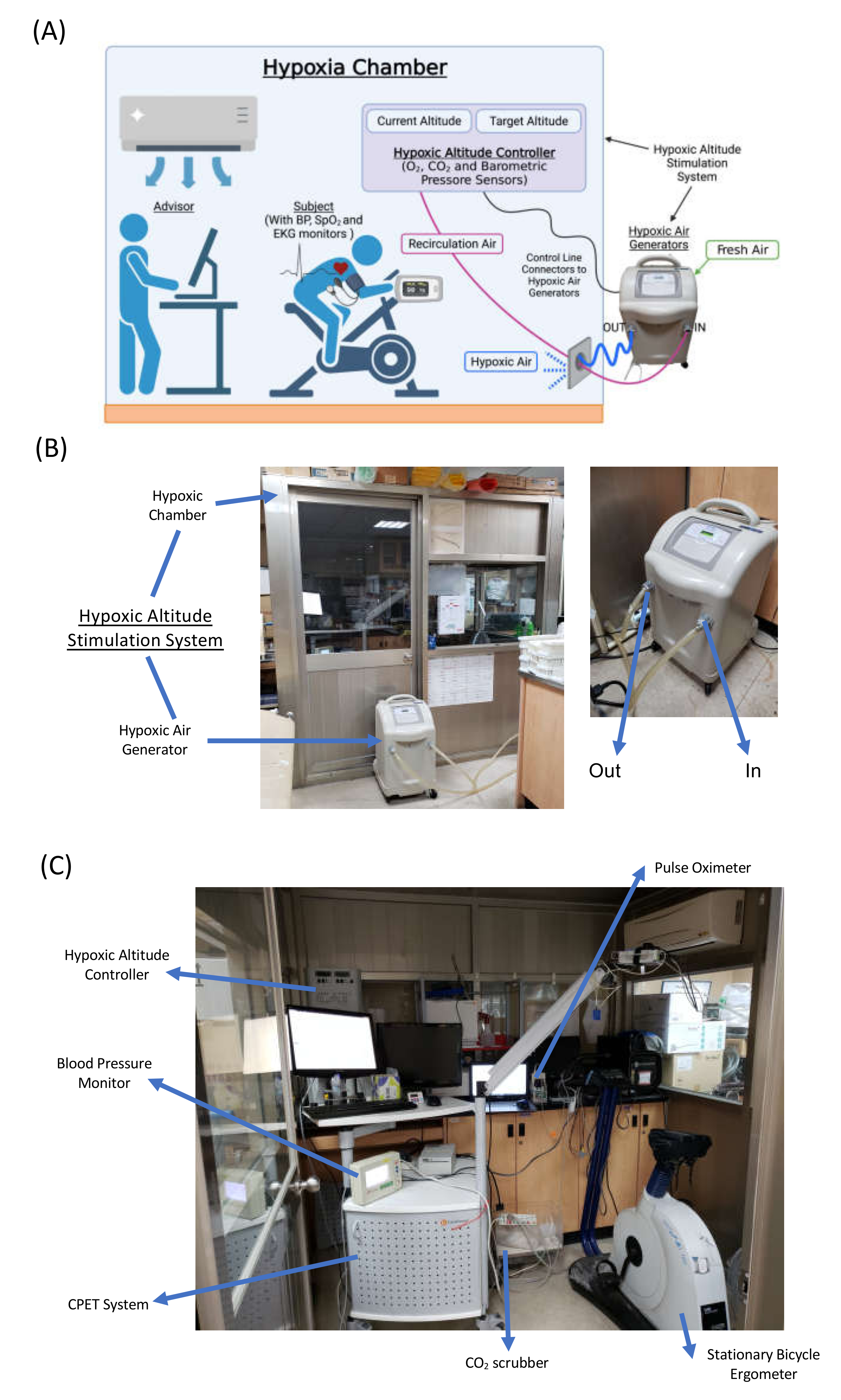
2.2. Protocol and Interventions
2.3. Cardiopulmonary Exercise Test (CPET)
2.4. Hypoxia Exercise Test (HET)
2.5. Erythrocyte Isolation and Blood Collection
2.6. Erythrocyte Aggregation
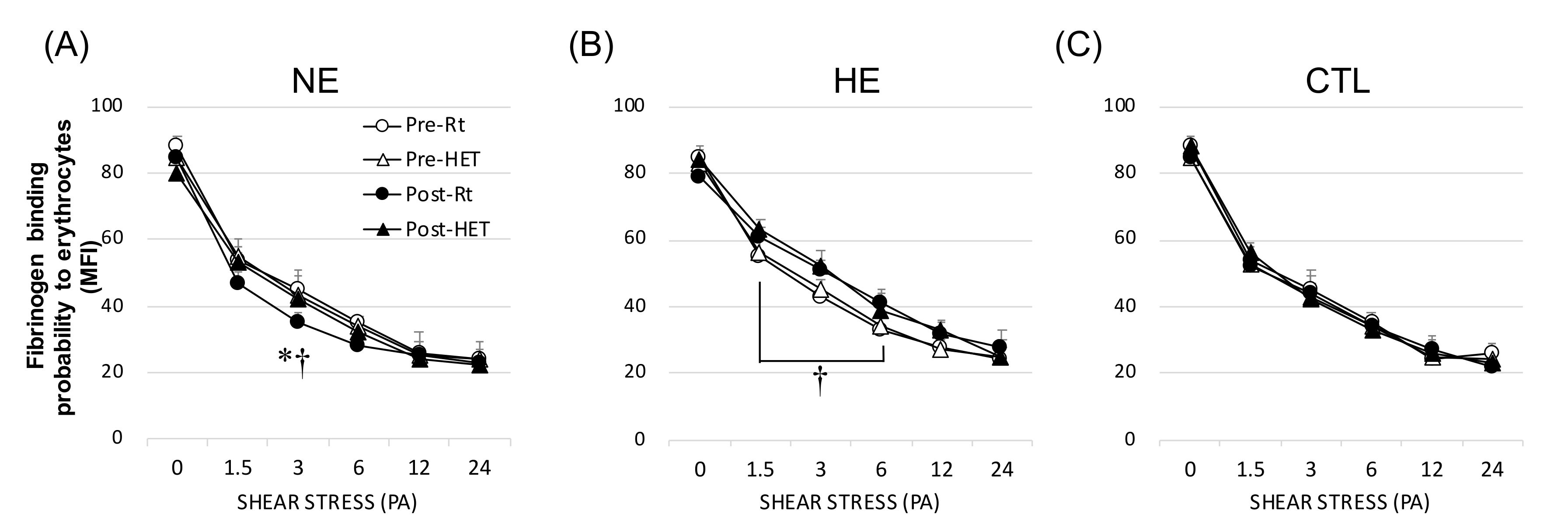
2.7. The Affinity between Erythrocytes and Fibrinogen
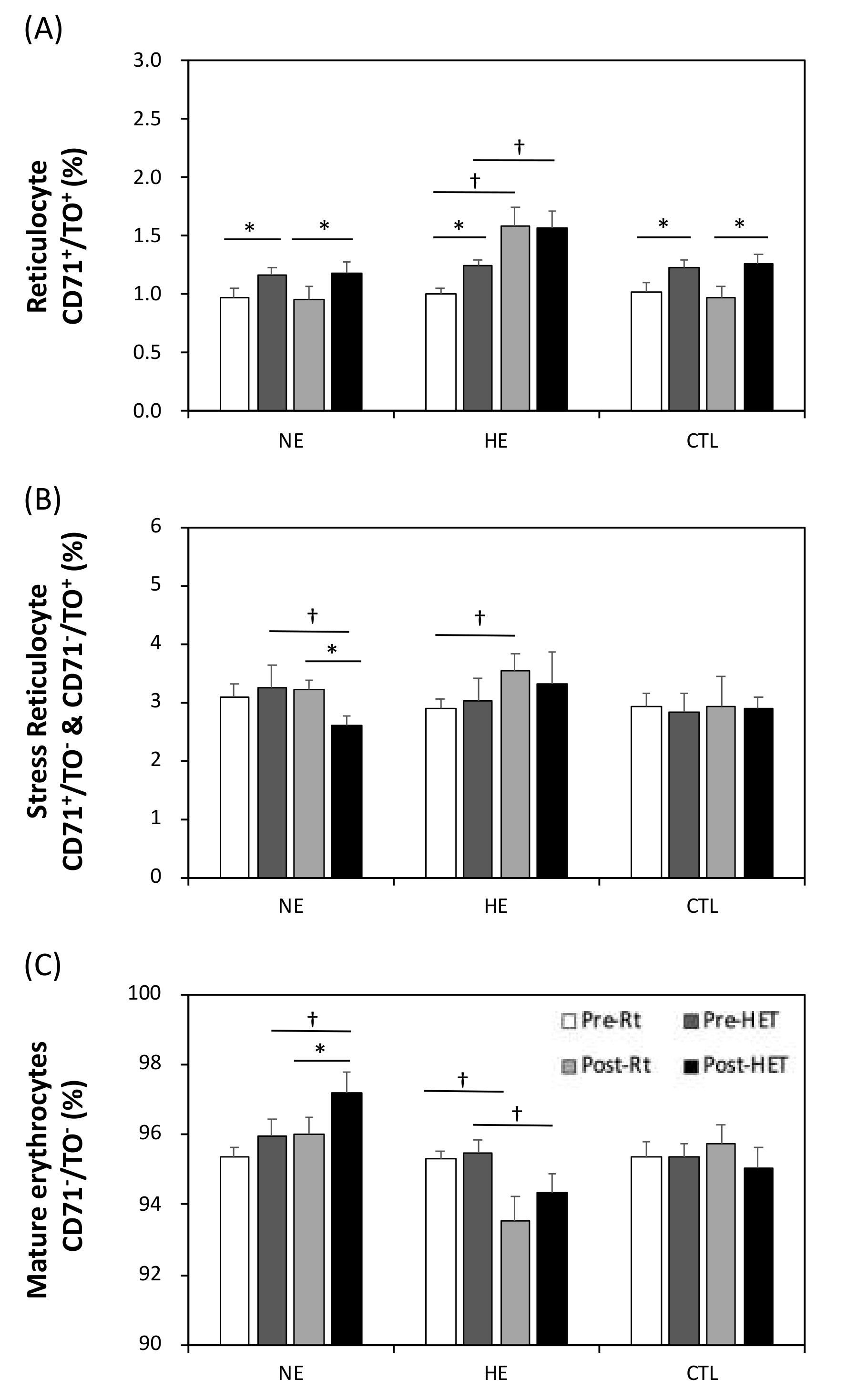
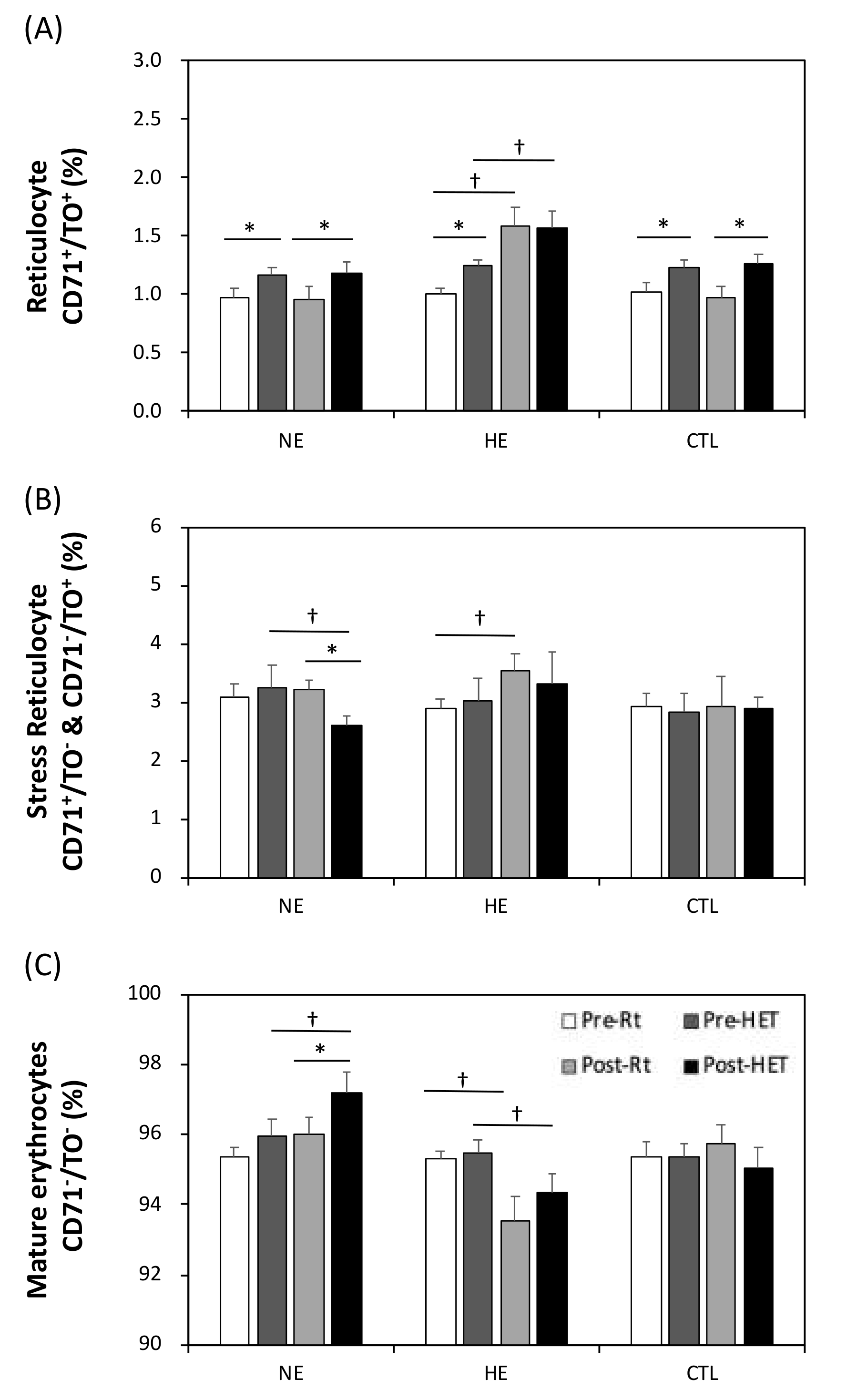
2.8. Reticulocyte Counts
2.9. Adhesion- and Senescence-Related Molecules on Erythrocytes
2.10. Statistical Analysis
3. Results
3.1. Cardiopulmonary Capacity
3.2. Hypoxia Exercise Test Performance
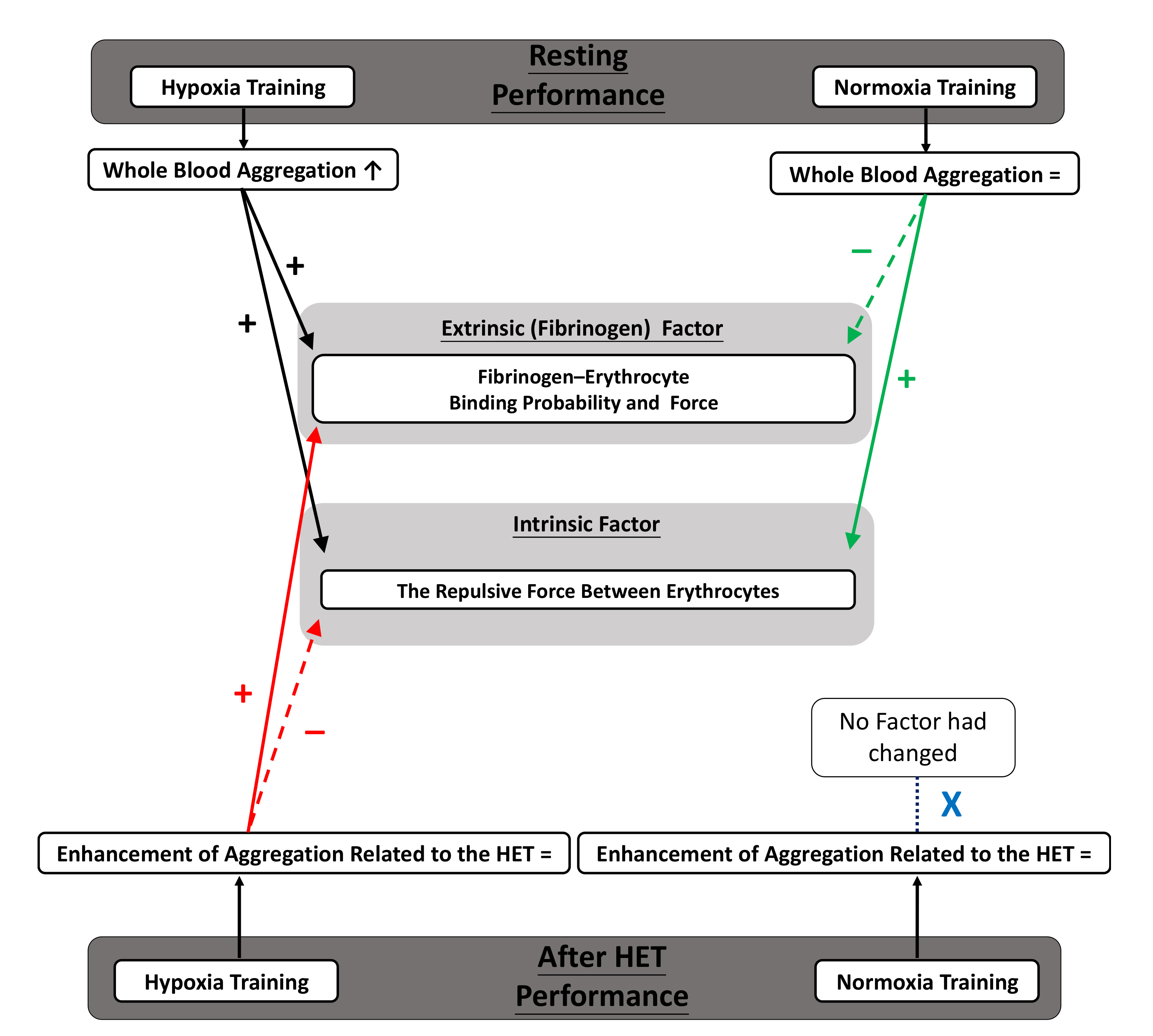
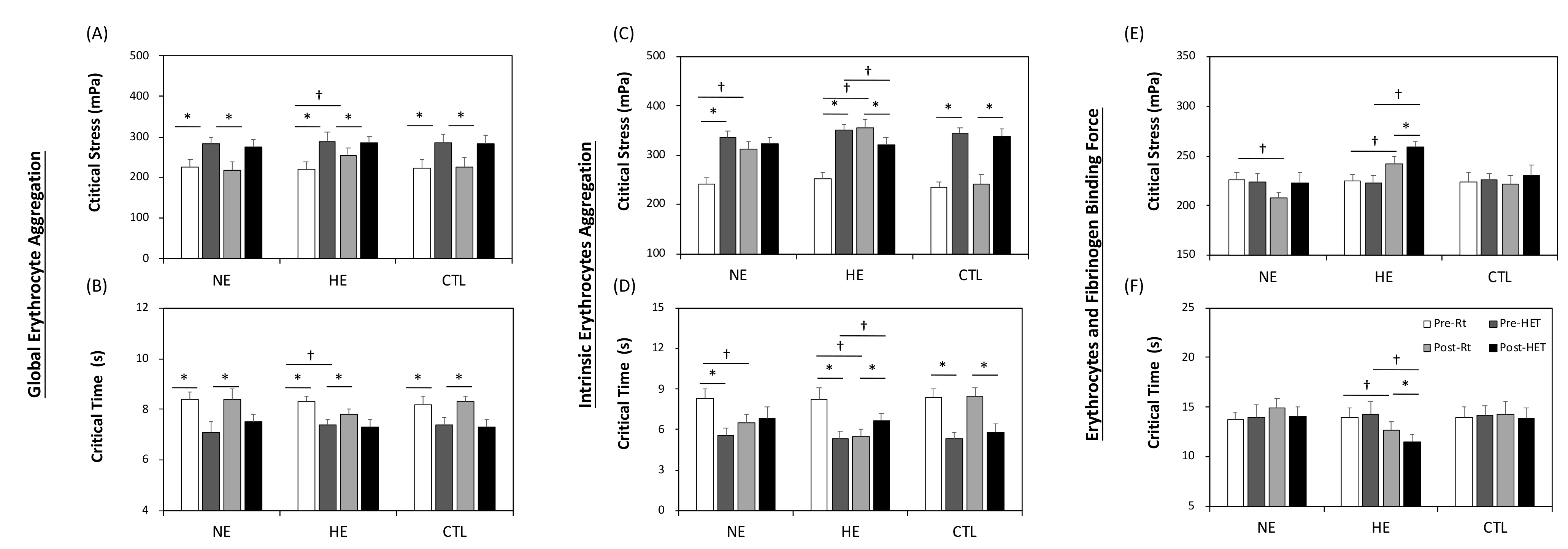
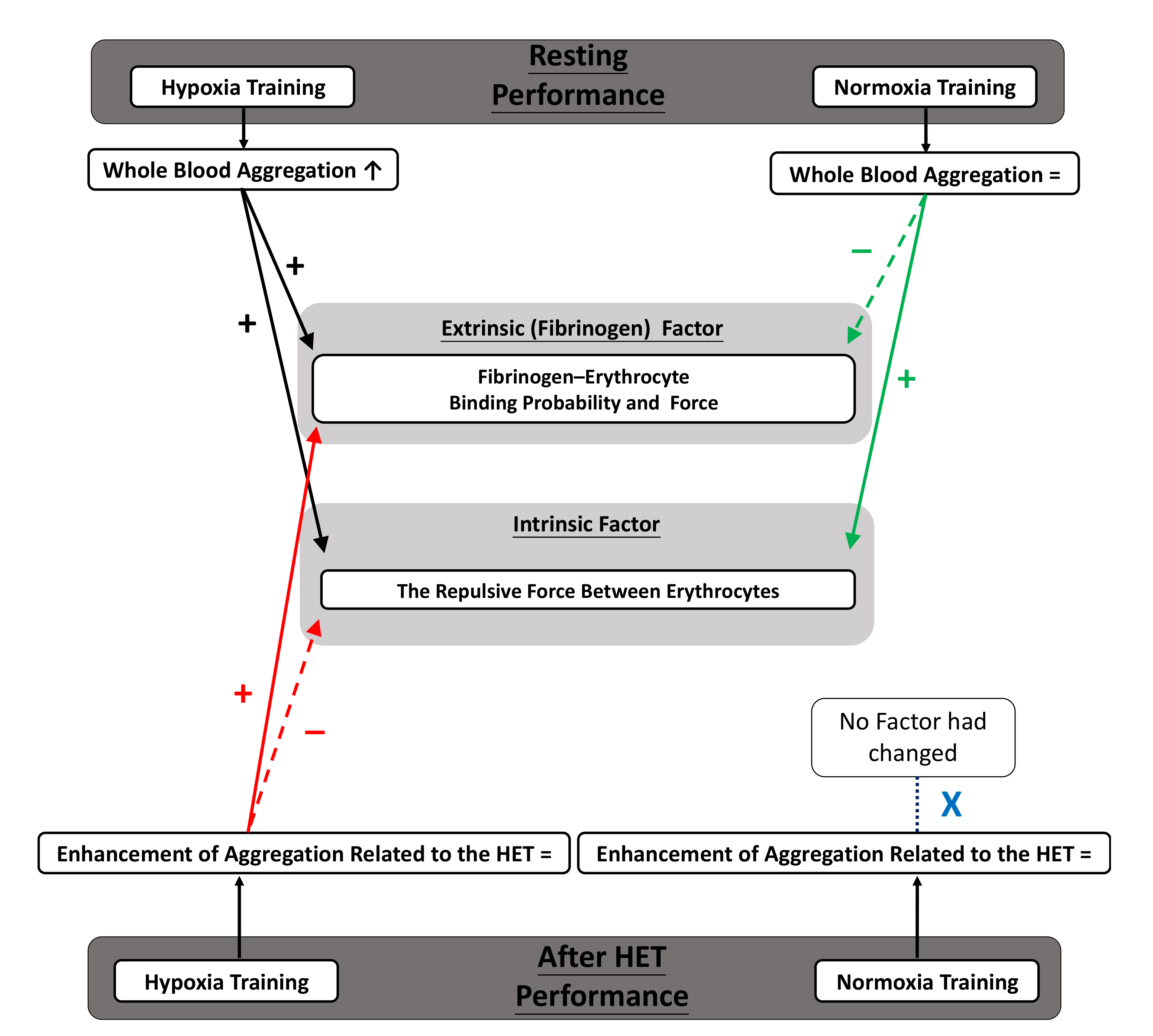
3.3. Whole Blood Aggregation
3.4. Intrinsic Factor of Aggregation
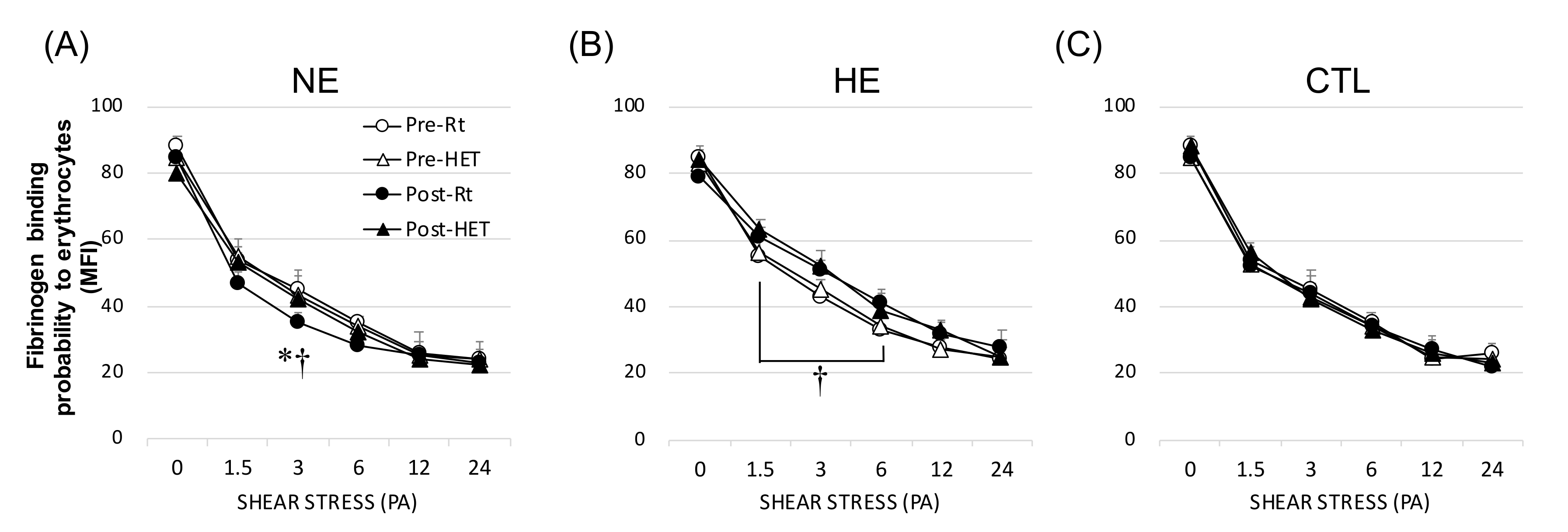
3.5. Fibrinogen (Extrinsic) Factor of Aggregation
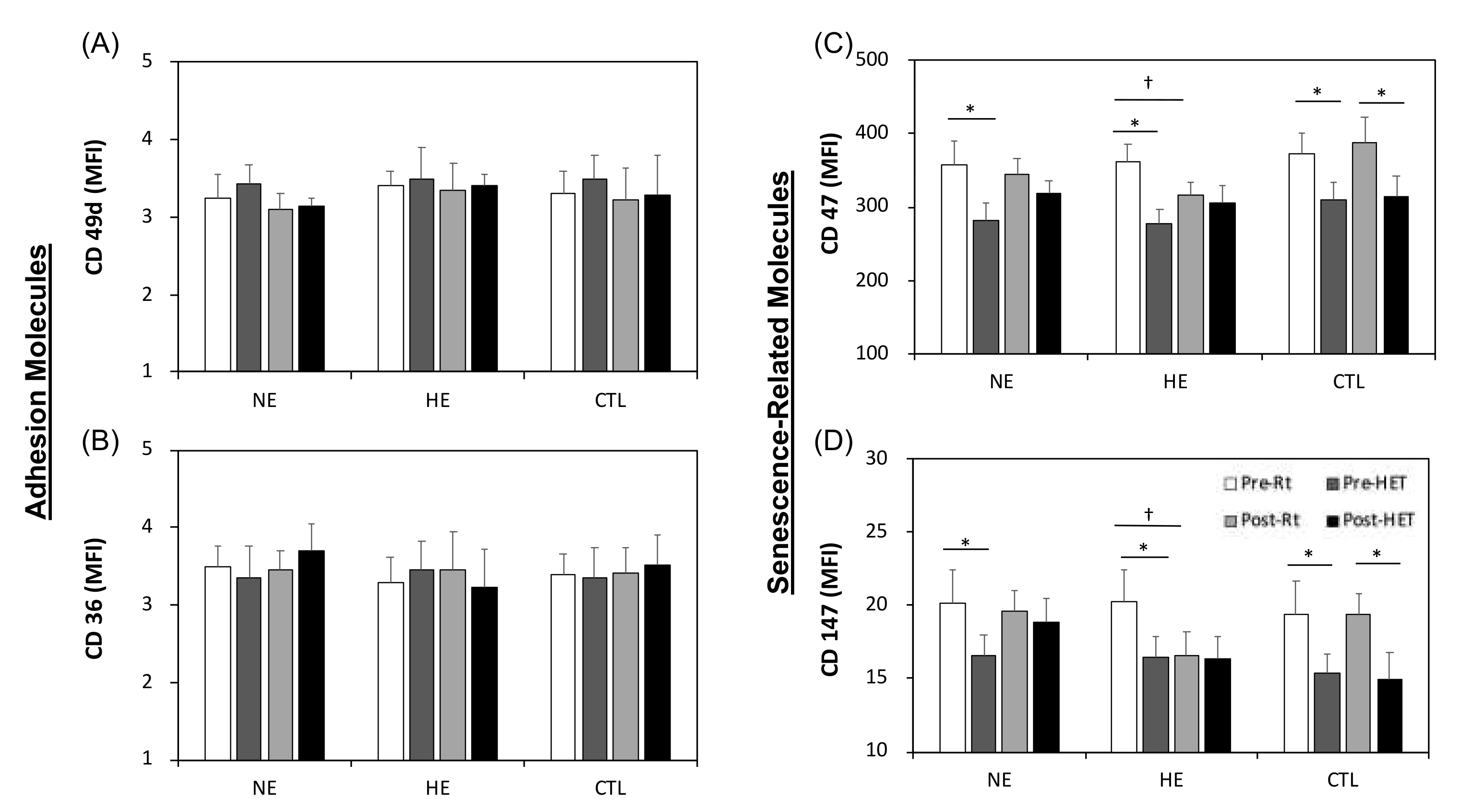
3.6. Adhesion- and Senescence-Related Molecules on Erythrocytes
3.7. The Relative Erythrocyte Subfractions
4. Discussion
4.1. Main Discussion
4.2. Clinical Prospects
4.3. Study Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saldanha, C.; P Lopes de Almeida, J. Erythrocyte as a link between basic and clinical research. Clin. Hemorheol. Microcirc. 2011, 49, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Tikhomirova, I.A.; Oslyakova, A.O.; Mikhailova, S.G. Microcirculation and blood rheology in patients with cerebrovascular disorders. Clin. Hemorheol. Microcirc. 2011, 49, 295–305. [Google Scholar] [CrossRef]
- Toker, S.; Rogowski, O.; Melamed, S.; Shirom, A.; Shapira, I.; Berliner, S.; Zeltser, D. Association of components of the metabolic syndrome with the appearance of aggregated red blood cells in the peripheral blood. An unfavorable hemorheological finding. Diabetes/Metab. Res. Rev. 2005, 21, 197–202. [Google Scholar] [CrossRef]
- Lipowsky, H.H. Microvascular rheology and hemodynamics. Microcirculation 2005, 12, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.A.; de Oliveira, S.; Freitas, T.; Gonçalves, S.; Santos, N.C. Variations on fibrinogen-erythrocyte interactions during cell aging. PLoS ONE 2011, 6, e18167. [Google Scholar] [CrossRef] [Green Version]
- Hierso, R.; Waltz, X.; Mora, P.; Romana, M.; Lemonne, N.; Connes, P.; Hardy-Dessources, M.D. Effects of oxidative stress on red blood cell rheology in sickle cell patients. Br. J. Haematol. 2014, 166, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Sewchand, L.S.; Canham, P.B. Modes of rouleaux formation of human red blood cells in polyvinylpyrrolidone and dextran solutions. Can. J. Physiol. Pharmacol. 1979, 57, 1213–1222. [Google Scholar] [CrossRef]
- Deng, L.H.; Barbenel, J.C.; Lowe, G.D. Influence of hematocrit on erythrocyte aggregation kinetics for suspensions of red blood cells in autologous plasma. Biorheology 1994, 31, 193–205. [Google Scholar] [CrossRef]
- Huang, Y.C.; Hsu, C.C.; Wang, J.S. High-Intensity Interval Training Improves Erythrocyte Osmotic Deformability. Med. Sci. Sports Exerc. 2019, 51, 1404–1412. [Google Scholar] [CrossRef] [PubMed]
- Raynaud, E.; Perez-Martin, A.; Brun, J.; Aïssa-Benhaddad, A.; Fédou, C.; Mercier, J. Relationships between fibrinogen and insulin resistance. Atherosclerosis 2000, 150, 365–370. [Google Scholar] [CrossRef]
- Wang, J.S.; Wu, M.H.; Mao, T.Y.; Fu, T.C.; Hsu, C.C. Effects of normoxic and hypoxic exercise regimens on cardiac, muscular, and cerebral hemodynamics suppressed by severe hypoxia in humans. J. Appl. Physiol. 2010, 109, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grygorczyk, R.; Orlov, S.N. Effects of Hypoxia on Erythrocyte Membrane Properties-Implications for Intravascular Hemolysis and Purinergic Control of Blood Flow. Front. Physiol. 2017, 8, 1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, T.Y.; Fu, L.L.; Wang, J.S. Hypoxic exercise training causes erythrocyte senescence and rheological dysfunction by depressed Gardos channel activity. J. Appl. Physiol. 2011, 111, 382–391. [Google Scholar] [CrossRef] [Green Version]
- Vij, A.G. Effect of prolonged stay at high altitude on platelet aggregation and fibrinogen levels. Platelets 2009, 20, 421–427. [Google Scholar] [CrossRef]
- Semenza, G.L.; Shimoda, L.A.; Prabhakar, N.R. Regulation of gene expression by HIF-1. Novartis Found. Symp. 2006, 272, 2–36. [Google Scholar]
- Rhodes, M.M.; Koury, S.T.; Kopsombut, P.; Alford, C.E.; Price, J.O.; Koury, M.J. Stress reticulocytes lose transferrin receptors by an extrinsic process involving spleen and macrophages. Am. J. Hematol. 2016, 91, 875–882. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.S.; Lin, H.Y.; Cheng, M.L.; Wong, M.K. Chronic intermittent hypoxia modulates eosinophil- and neutrophil-platelet aggregation and inflammatory cytokine secretion caused by strenuous exercise in men. J. Appl. Physiol. 2007, 103, 305–314. [Google Scholar] [CrossRef] [Green Version]
- Pialoux, V.; Hanly, P.J.; Foster, G.E.; Brugniaux, J.V.; Beaudin, A.E.; Hartmann, S.E.; Pun, M.; Duggan, C.T.; Poulin, M.J. Effects of exposure to intermittent hypoxia on oxidative stress and acute hypoxic ventilatory response in humans. Am. J. Respir. Crit. Care Med. 2009, 180, 1002–1009. [Google Scholar] [CrossRef]
- Fu, Q.; Townsend, N.E.; Shiller, S.M.; Martini, E.R.; Okazaki, K.; Shibata, S.; Truijens, M.J.; Rodríguez, F.A.; Gore, C.J.; Stray-Gundersen, J.; et al. Intermittent hypobaric hypoxia exposure does not cause sustained alterations in autonomic control of blood pressure in young athletes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1977–R1984. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.H.; Macfarlane, D.J.; Lam, T.H.; Stewart, S.M. Validity of the International Physical Activity Questionnaire Short Form (IPAQ-SF): A systematic review. Int. J. Behav. Nutr. Phys. Act. 2011, 8, 115. [Google Scholar] [CrossRef] [Green Version]
- Ortega, R.M.; Perez-Rodrigo, C.; Lopez-Sobaler, A.M. Dietary assessment methods: Dietary records. Nutr. Hosp. 2015, 31 (Suppl. 3), 38–45. [Google Scholar] [CrossRef] [PubMed]
- Thompson, W.R.; Hubertus, G.M.; Pescatello, L.S. Clinical Exercise Testing. In ACSM’s Guidelines for Exercise Testing and Prescription; American College of Sports Medicine: Indianapolis, IN, USA, 2017. [Google Scholar]
- Chen, Y.C.; Ho, C.W.; Tsai, H.H.; Wang, J.S. Interval and continuous exercise regimens suppress neutrophil-derived microparticle formation and neutrophil-promoted thrombin generation under hypoxic stress. Clin. Sci. 2015, 128, 425–436. [Google Scholar] [CrossRef]
- Samsel, L.; McCoy, J.P., Jr. Imaging flow cytometry for the study of erythroid cell biology and pathology. J. Immunol. Methods 2015, 423, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.G.; Chen, C.H.; Chiu, L.A. Thiazole orange: A new dye for reticulocyte analysis. Cytometry 1986, 7, 508–517. [Google Scholar] [CrossRef]
- Baskurt, O.; Meiselman, H. Blood Rheology and Hemodynamics. Semin. Thromb. Hemost. 2003, 29, 435–450. [Google Scholar] [CrossRef] [Green Version]
- Brun, J.F.; Connes, P.; Varlet-Marie, E. Alterations of blood rheology during and after exercise are both consequences and modifiers of body’s adaptation to muscular activity. Sci. Sports 2007, 22, 251–266. [Google Scholar] [CrossRef]
- Mairbäurl, H. Red blood cells in sports: Effects of exercise and training on oxygen supply by red blood cells. Front. Physiol. 2013, 4, 332. [Google Scholar] [CrossRef] [Green Version]
- Morceau, F.; Dicato, M.; Diederich, M. Pro-Inflammatory Cytokine-Mediated Anemia: Regarding Molecular Mechanisms of Erythropoiesis. Mediat. Inflamm. 2009, 405016. [Google Scholar] [CrossRef] [Green Version]
- Prabhakar, N.R.; Kumar, G.K.; Nanduri, J.; Semenza, G.L. ROS Signaling in Systemic and Cellular Responses to Chronic Intermittent Hypoxia. Antioxid. Redox Signal. 2007, 9, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- Dyugovskaya, L.; Polyakov, A.; Ginsberg, D.; Lavie, P.; Lavie, L. Molecular pathways of spontaneous and TNF-α-mediated neutrophil apoptosis under intermittent hypoxia. Am. J. Respir. Cell Mol. Biol. 2011, 45, 154–162. [Google Scholar] [CrossRef]
- Wang, J.S.; Fu, T.C.; Wang, C.H.; Chou, S.L.; Liu, M.H.; Cherng, W.J. Exertional periodic breathing potentiates erythrocyte rheological dysfunction by elevating pro-inflammatory status in patients with anemic heart failure. Int. J. Cardiol. 2013, 167, 1289–1297. [Google Scholar] [CrossRef]
- Wang, J.-S.; Chen, W.-L.; Weng, T.-P. Hypoxic exercise training reduces senescent T-lymphocyte subsets in blood. BrainBehav. Immun. 2011, 25, 270–278. [Google Scholar] [CrossRef]
- Wang, J.S.; Lin, C.T. Systemic hypoxia promotes lymphocyte apoptosis induced by oxidative stress during moderate exercise. Eur. J. Appl. Physiol. 2010, 108, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Varlet-Marie, E.; Brun, J.F. Reciprocal relationships between blood lactate and hemorheology in athletes: Another hemorheologic paradox? Clin. Hemorheol. Microcirc. 2004, 30, 331–337. [Google Scholar]
- Guzy, R.D.; Hoyos, B.; Robin, E.; Chen, H.; Liu, L.; Mansfield, K.D.; Simon, M.C.; Hammerling, U.; Schumacker, P.T. Mitochondrial complex III is required for hypoxia-induced ROS production and cellular oxygen sensing. Cell Metab. 2005, 1, 401–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Yoon, D.; Christensen, R.D.; Horvathova, M.; Thiagarajan, P.; Prchal, J.T. HIF-mediated increased ROS from reduced mitophagy and decreased catalase causes neocytolysis. J. Mol. Med. 2015, 93, 857–866. [Google Scholar] [CrossRef]
- Ventura, N.; Hoppeler, H.; Seiler, R.; Binggeli, A.; Mullis, P.-E.; Vogt, M. The Response of Trained Athletes to Six Weeks of Endurance Training in Hypoxia or Normoxia. Int. J. Sports Med. 2003, 24, 166–172. [Google Scholar] [CrossRef]
- Huang, Y.X.; Wu, Z.J.; Mehrishi, J.; Huang, B.T.; Chen, X.Y.; Zheng, X.J.; Liu, W.J.; Luo, M. Human red blood cell aging: Correlative changes in surface charge and cell properties. J. Cell. Mol. Med. 2011, 15, 2634–2642. [Google Scholar] [CrossRef] [Green Version]
- Baskurt, O.N.; Neu., B.; Meiselman, H.J. Red Blood Cell Aggregation, 1st ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 31–59. [Google Scholar]
- Neu, B.; Sowemimo-Coker, S.O.; Meiselman, H.J. Cell-cell affinity of senescent human erythrocytes. Biophys. J. 2003, 85, 75–84. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, S.; Vitorino de Almeida, V.; Calado, A.; Rosário, H.S.; Saldanha, C. Integrin-associated protein (CD47) is a putative mediator for soluble fibrinogen interaction with human red blood cells membrane. Biochim. Et Biophys. Acta 2012, 1818, 481–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiodorenko-Dumas, Ż.; Dumas, I.; Mastej, K.; Jakobsche-Policht, U.; Bittner, J.; Adamiec, R. Receptor GP IIb/IIIa as an Indicator of Risk in Vascular Events. Clin. Appl. Thromb. Hemost. 2019, 25, 1076029619845056. [Google Scholar] [CrossRef] [Green Version]
- Viscor, G.; Torrella, J.R.; Corral, L.; Ricart, A.; Javierre, C.; Pages, T.; Ventura, J.L. Physiological and Biological Responses to Short-Term Intermittent Hypobaric Hypoxia Exposure: From Sports and Mountain Medicine to New Biomedical Applications. Front. Physiol. 2018, 9, 814. [Google Scholar] [CrossRef]
- Latshang, T.D.; Tardent, R.P.M.; Furian, M.; Flueck, D.; Segitz, S.D.; Mueller-Mottet, S.; Kohler, M.; Ulrich, S.; Bloch, K.E. Sleep and breathing disturbances in patients with chronic obstructive pulmonary disease traveling to altitude: A randomized trial. Sleep 2019, 42. [Google Scholar] [CrossRef]
- Mestry, N.; Thirumaran, M.; Tuggey, J.M.; Macdonald, W.; Elliott, M.W. Hypoxic challenge flight assessments in patients with severe chest wall deformity or neuromuscular disease at risk for nocturnal hypoventilation. Thorax 2009, 64, 532–534. [Google Scholar] [CrossRef] [Green Version]
- Soguel Schenkel, N.; Burdet, L.; de Muralt, B.; Fitting, J.W. Oxygen saturation during daily activities in chronic obstructive pulmonary disease. Eur. Respir. J. 1996, 9, 2584–2589. [Google Scholar] [CrossRef]
- Seccombe, L.M.; Kelly, P.T.; Wong, C.K.; Rogers, P.G.; Lim, S.; Peters, M.J. Effect of simulated commercial flight on oxygenation in patients with interstitial lung disease and chronic obstructive pulmonary disease. Thorax 2004, 59, 966–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verges, S.; Chacaroun, S.; Godin-Ribuot, D.; Baillieul, S. Hypoxic Conditioning as a New Therapeutic Modality. Front. Pediatrics 2015, 3, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ugurlu, E.; Kilic-Toprak, E.; Can, I.; Kilic-Erkek, O.; Altinisik, G.; Bor-Kucukatay, M. Impaired Hemorheology in Exacerbations of COPD. Can. Respir. J. 2017, 2017, 1286263. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NE | HE | CTL | ||||
|---|---|---|---|---|---|---|
| Pretraining | Post-Training | Pretraining | Post-Training | Pretraining | Post-Training | |
| Anthropometric variables | ||||||
| Age (years) | 22.4 ± 0.9 | --- | 21.9 ± 0.7 | --- | 23.1 ± 0.9 | --- |
| Height (cm) | 173 ± 2 | --- | 173 ± 3 | --- | 174 ± 2 | --- |
| Weight (kg) | 68.1 ± 2.2 | 66.9 ± 3.1 | 66.9 ± 2.5 | 66.7 ± 2.3 | 65.7 ± 2.3 | 66.2 ± 1.8 |
| Ventilation threshold performance | ||||||
| Work Rate (watt) | 121 ± 7 | 155 ± 7 * | 123 ± 4 | 156 ± 4 * | 122 ± 5 | 124 ± 4 |
| E (L min−1) | 55.5 ± 3.6 | 72.0 ± 4.6 * | 55.8 ± 4.5 | 70.4 ± 4.3 * | 55.6 ± 4.1 | 56.3 ± 4.2 |
| O2 (mL kg−1 min−1) | 23.7 ± 1.3 | 30.0 ± 1.8 * | 23.9 ± 1.7 | 28.2 ± 1.3 * | 23.4 ± 1.6 | 23.2 ± 1.5 |
| HR (bpm) | 149 ± 3 | 157 ± 3 * | 147 ± 4 | 153 ± 3 | 147 ± 4 | 149 ± 3 |
| CO (L min−1) | 14.3 ± 0.6 | 15.1 ± 0.6 | 14.4 ± 0.5 | 15.6 ± 0.8 | 14.1 ± 0.4 | 14.4 ± 0.6 |
| MAP (mmHg) | 119 ± 2 | 123 ± 3 | 117 ± 2 | 118 ± 2 | 116 ± 3 | 115 ± 2 |
| Peak performance | ||||||
| Work Rate (watt) | 187 ± 5 | 216 ± 5 * | 189 ± 5 | 228 ± *,† | 188 ± 5 | 189 ± 5 |
| E (L min−1) | 117.9 ± 3.4 | 124.4 ± 3.9 * | 117.7 ± 4.8 | 137.9 ± 3.6 *,† | 116.86 ± 3.9 | 117.9 ± 3.1 |
| O2 (ml kg−1 min−1) | 33.6 ± 1.1 | 36.3 ± 1.1 * | 33.4 ± 1.4 | 38.8 ± 1.3 *,† | 33.8 ± 1.3 | 32.2 ± 1.1 |
| HR (bpm) | 195 ± 2 | 193 ± 2 | 193 ± 2 | 193 ± 2 | 193 ± 2 | 194 ± 3 |
| CO (L min−1) | 16.0 ± 0.5 | 16.3 ± 0.4 | 16.3 ± 0.6 | 18 ± 0.5 * | 16.2 ± 0.4 | 15.9 ± 0.5 |
| MAP (mmHg) | 128 ± 2 | 133 ± 3 | 129 ± 2 | 132 ± 2 | 129 ± 2 | 128 ± 3 |
| NE | HE | CTL | |||||
|---|---|---|---|---|---|---|---|
| Pretraining | Post-Training | Pretraining | Post-Training | Pretraining | Post-Training | ||
| HR (bpm) | |||||||
| Rt | 77 ± 3 | 81 ± 5 | 80 ± 2 | 78 ± 3 | 77 ± 2 | 78 ± 4 | |
| Ex | 154 ± 2 * | 142 ± 5 *,† | 156 ± 3 * | 141 ± 2 *,† | 153 ± 3 * | 151 ± 3 * | |
| SpO2 (%) | |||||||
| Rt | 94 ± 1 | 94 ± 1 | 93 ± 1.1 | 94 ± 1 | 93 ± 2 | 94 ± 1 | |
| Ex | 79 ± 1 * | 80 ± 1 * | 80 ± 1 * | 79 ± 1 * | 80 ± 1 * | 80 ± 1 * | |
| Lac (mM) | |||||||
| Rt | 1.4 ± 0.2 | 1.1 ± 0.2 | 1 ± 0.2 | 1 ± 0.1 | 1 ± 0.2 | 1.4 ± 0.2 | |
| Ex | 10.3 ± 0.5 * | 8.4 ± 0.5 *,† | 10.9 ± 1 * | 7.0 ± 0.5 *,†,‡ | 11.2 ± 0.8 * | 10.7 ± 0.5 * | |
| pH (unit) | |||||||
| Rt | 7.37 ± 0.01 | 7.37 ± 0.01 | 7.36 ± 0.01 | 7.36 ± 0.01 | 7.36 ± 0.02 | 7.37 ± 0.01 | |
| Ex | 7.31 ± 0.01 * | 7.35 ± 0.01 † | 7.29 ± 0.01 * | 7.33 ± 0.01 *,† | 7.30 ± 0.01 * | 7.31 ± 0.01 * | |
| PCO2 (mmHg) | |||||||
| Rt | 52.2 ± 2.3 | 47.4 ± 2.1 | 52.4 ± 1.5 | 47.5 ± 1.7 | 52.4 ± 1.5 | 47.5 ± 1.7 | |
| Ex | 41.1 ± 2.8 * | 40.3 ± 1.5 * | 39.2 ± 2 * | 39.2 ± 1 * | 39.2 ± 2 * | 39.2 ± 1 * | |
| Erythrocyte (106/µL) | |||||||
| Rt | 4.65 ± 0.06 | 4.64 ± 0.07 | 4.65 ± 0.08 | 4.57 ± 0.08 | 4.72 ± 0.11 | 4.57 ± 0.08 | |
| Ex | 4.92 ± 0.07 * | 4.84 ± 0.07 * | 5.03 ± 0.12 * | 4.83 ± 0.08 * | 4.99 ± 0.09 * | 4.93 ± 0.08 * | |
| Hemoglobin (g/dL) | |||||||
| Rt | 14.1 ± 0.2 | 14.0 ± 0.2 | 13.9 ± 0.2 | 14.0 ± 0.2 | 14.1 ± 0.1 | 14.0 ± 0.3 | |
| Ex | 15.0 ± 0.2 * | 14.7 ± 0.3 * | 14.8 ± 0.2 * | 14.3 ± 0.2 * | 14.3 ± 0.2 * | 14.4 ± 0.2 * | |
| Hematocrit (%) | |||||||
| Rt | 41.2 ± 0.6 | 41.2 ± 0.6 | 40.8 ± 0.4 | 39.7 ± 0.5 | 40.9 ± 0.4 | 40.2 ± 0.6 | |
| Ex | 44.0 ± 0.6 * | 43.3 ± 0.7 * | 43.7 ± 0.4 * | 42.2 ± 0.6 * | 43.3 ± 0.3 * | 42.1 ± 0.5 * | |
| Erythropoietin (mIU/mL) | |||||||
| Rt | 17.5 ± 1.9 | 20.9 ± 2.7 | 16.9 ± 2.6 | 22.2 ± 2.8 † | 17.1 ± 2.5 | 17.6 ± 2.5 | |
| Ex | 28.2 ± 2.6 * | 23.6 ± 2.5 | 32.8 ± 3.7 * | 25.8 ± 2.6 † | 32.2 ± 3.2 * | 31.8 ± 2.2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.-L.; Wang, J.-S.; Fu, T.-C.; Hsu, C.-C.; Huang, Y.-C. Hypoxic Exercise Training Elevates Erythrocyte Aggregation. Appl. Sci. 2021, 11, 6038. https://doi.org/10.3390/app11136038
Lin C-L, Wang J-S, Fu T-C, Hsu C-C, Huang Y-C. Hypoxic Exercise Training Elevates Erythrocyte Aggregation. Applied Sciences. 2021; 11(13):6038. https://doi.org/10.3390/app11136038
Chicago/Turabian StyleLin, Chin-Li, Jong-Shyan Wang, Tieh-Cheng Fu, Chih-Chin Hsu, and Yu-Chieh Huang. 2021. "Hypoxic Exercise Training Elevates Erythrocyte Aggregation" Applied Sciences 11, no. 13: 6038. https://doi.org/10.3390/app11136038
APA StyleLin, C.-L., Wang, J.-S., Fu, T.-C., Hsu, C.-C., & Huang, Y.-C. (2021). Hypoxic Exercise Training Elevates Erythrocyte Aggregation. Applied Sciences, 11(13), 6038. https://doi.org/10.3390/app11136038