
The Structural Characteristics of Collagen in Swim Bladders with 25-Year Sequence Aging: The Impact of Age


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Moisture and Collagen Content
2.3. Composition of Amino Acids
2.4. Thermogravimetric and Differential Scanning Calorimetry (TG-DSC) Analysis
2.5. Attenuated Total Reflectance–Fourier Transform Infrared Spectroscopy (ATR–FTIR)
2.6. X-Ray Diffraction (XRD) Analysis
2.7. Atomic Force Microscope (AFM) Analysis
2.8. Optical Microscopy Analysis
2.9. Scanning Electron Microscope (SEM) Analysis
2.10. Statistical Analysis
3. Results
3.1. Moisture, Collagen, and Amino Acid Compositions of Swim Bladders
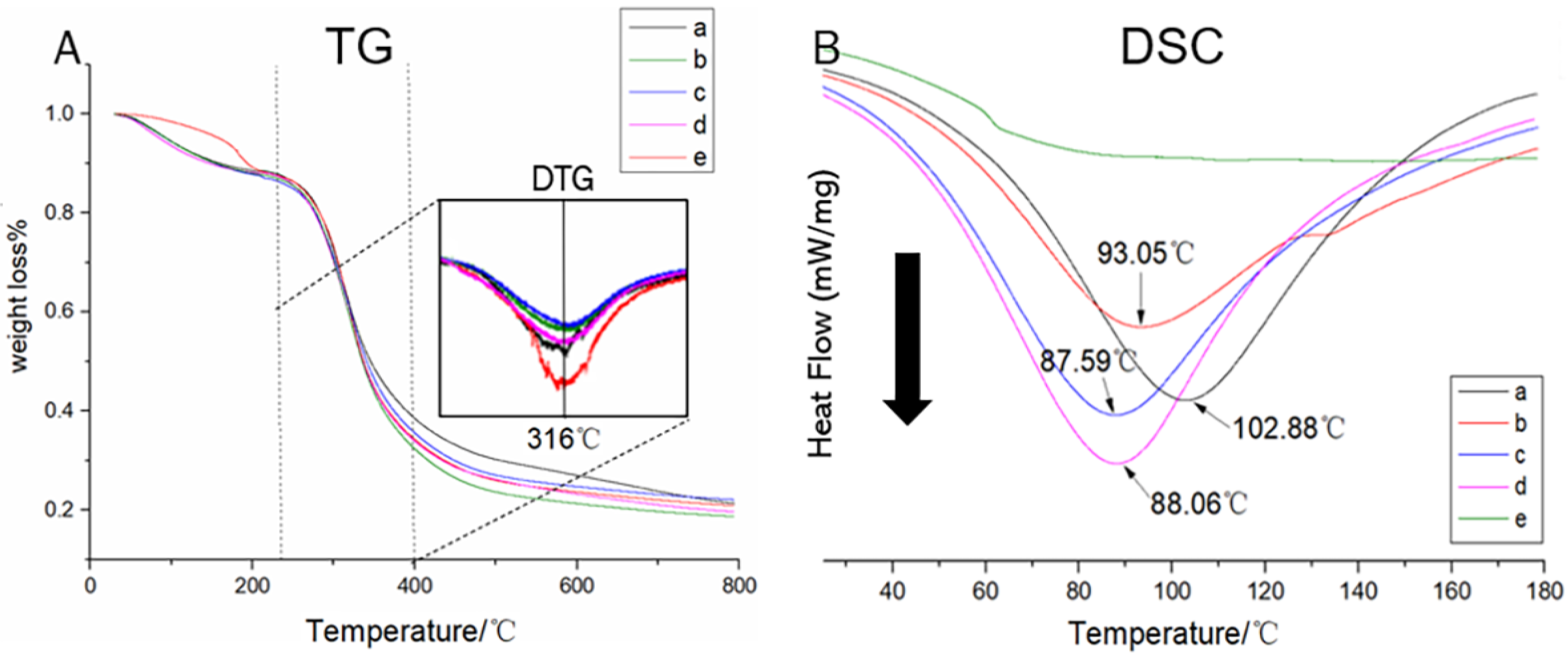
3.2. Thermal Decomposition Profiles of the Swim Bladder
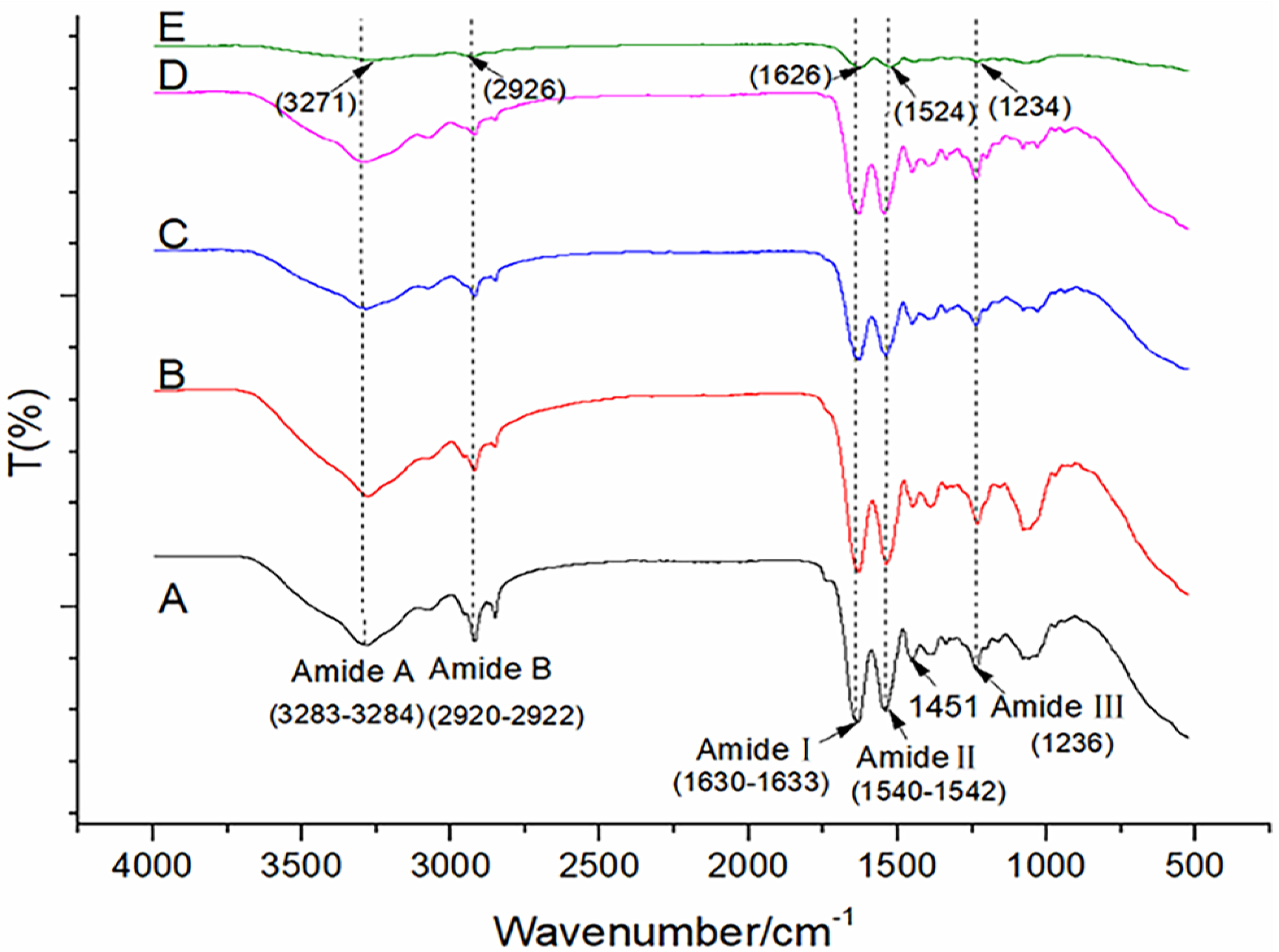
3.3. ATR–FTIR Spectrum of the Triple Helix in the Swim Bladder
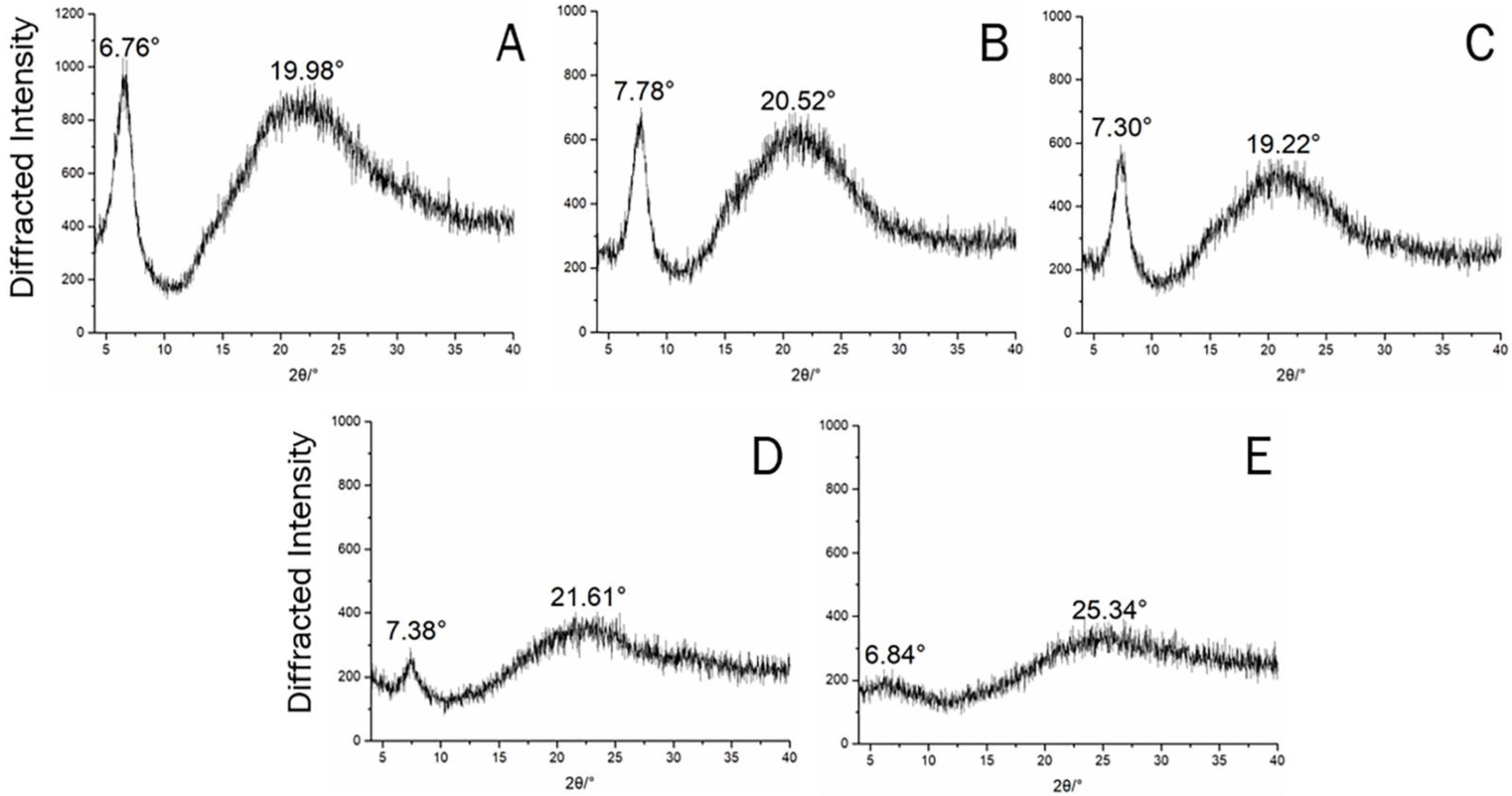
3.4. XRD Spectrum of Triple Helix Structures in the Swim Bladder
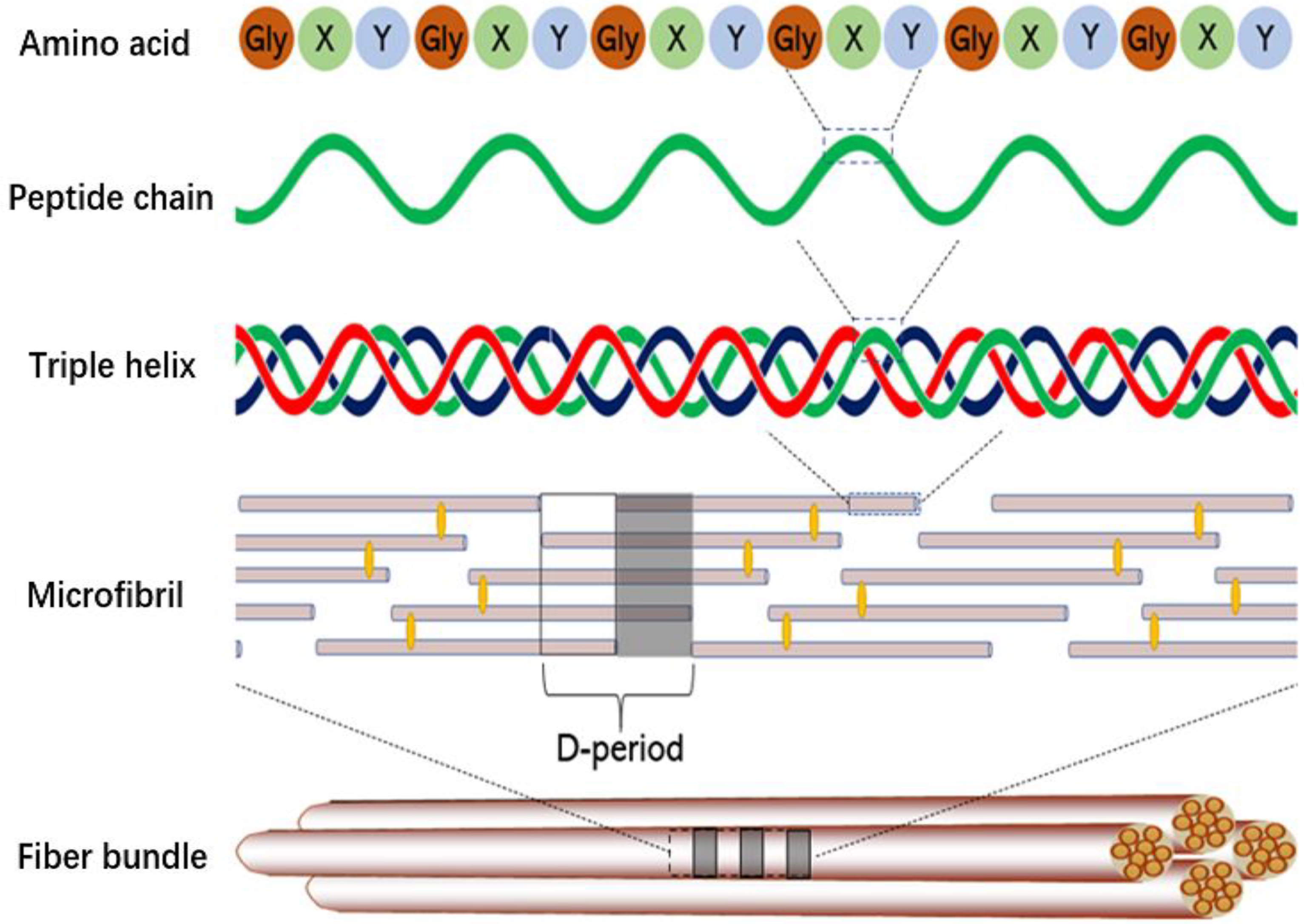
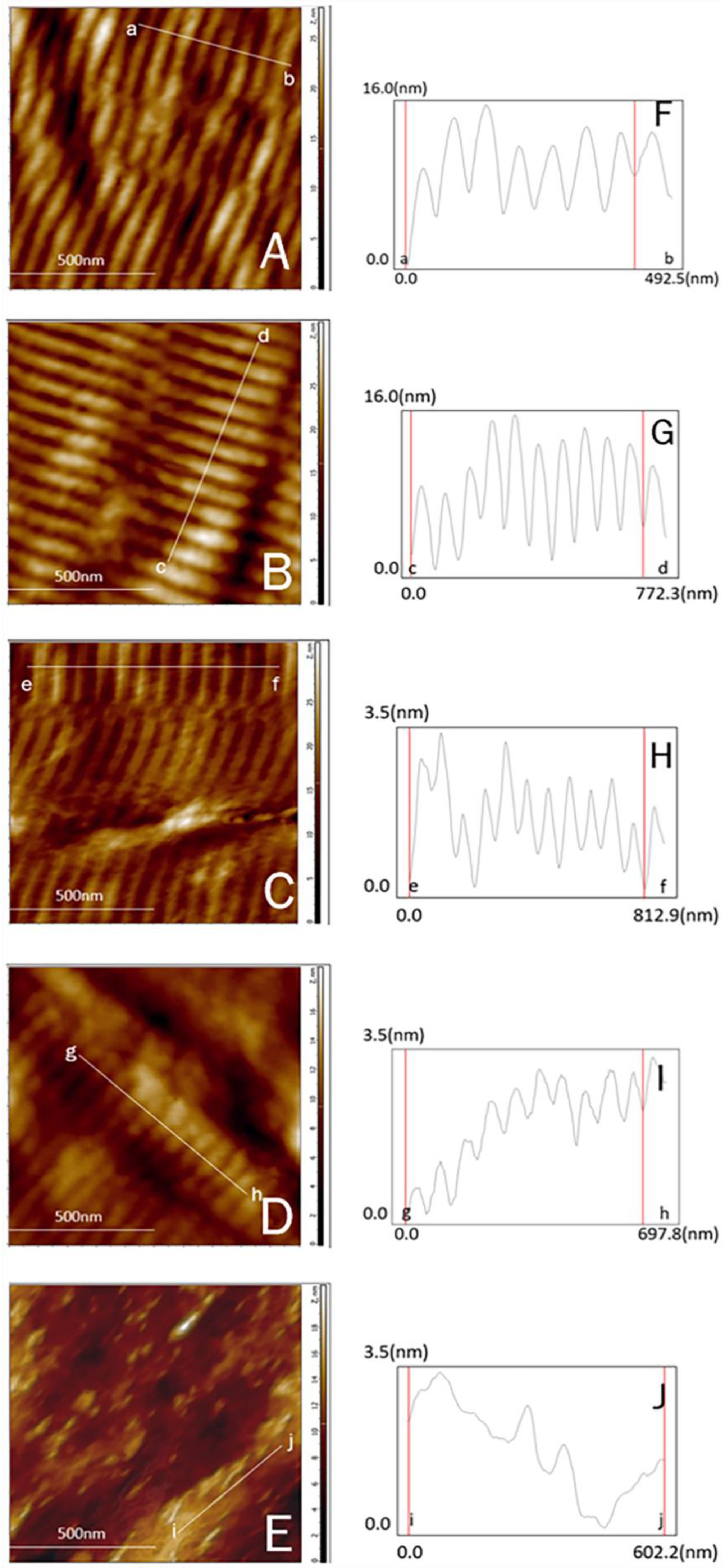
3.5. Characteristics of Microfibrils in Swim Bladders
3.6. Optical Microscopy and SEM Observation of Collagen Fiber Bundles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Fänge, R. Gas exchange in fish swim bladder. Rev. Physiol. Biochem. Pharmacol. 1983, 97, 111–158. [Google Scholar] [CrossRef] [PubMed]
- De Mitcheson, Y.S.; To, A.W.-L.; Wong, N.W.; Kwan, H.Y.; Bud, W.S. Emerging from the murk: Threats, challenges and opportunities for the global swim bladder trade. Rev. Fish Biol. Fisher. 2019, 29, 809–835. [Google Scholar] [CrossRef]
- Liu, D.; Liang, L.; Regenstein, J.M.; Zhou, P. Extraction and characterisation of pepsin-solubilised collagen from fins, scales, skins, bones and swim bladders of bighead carp (Hypophthalmichthys nobilis). Food Chem. 2012, 133, 1441–1448. [Google Scholar] [CrossRef]
- Zhu, S.; Yuan, Q.; Yin, T.; You, J.; Gu, Z.; Xiong, S.; Hu, Y. Self-assembly of collagen-based biomaterials: Preparation, characterizations and biomedical applications. J. Mater. Chem. B 2018, 6, 2650–2676. [Google Scholar] [CrossRef]
- Palmieri, B. Heterologous collagen in wound healing: A clinical study. Int. J. Tissue React. 1992, 14, 21–25. [Google Scholar] [CrossRef]
- Brown, E.M. Development and utilization of a bovine type I collagen microfibril model. Int. J. Biol. Macromol. 2013, 53, 20–25. [Google Scholar] [CrossRef]
- Sousa, R.O.; Alves, A.L.; Carvalho, D.N.; Martins, E.; Oliveira, C.; Silva, T.H.; Reis, R.L. Acid and enzymatic extraction of collagen from Atlantic cod (Gadus Morhua) swim bladders envisaging health-related applications. J. Biomater. Sci. Polym. Ed. 2020, 31, 20–37. [Google Scholar] [CrossRef]
- Maxwell, C.A.; Wess, T.J.; Kennedy, C.J. X-ray Diffraction Study into the Effects of Liming on the Structure of Collagen. Biomacromolecules 2006, 7, 2321–2326. [Google Scholar] [CrossRef]
- Meng, Z.; Zheng, X.; Tang, K.; Liu, J.; Ma, Z.; Zhao, Q. Dissolution and regeneration of collagen fibers using ionic liquid. Int. J. Biol. Macromol. 2012, 51, 440–448. [Google Scholar] [CrossRef]
- Zhao, X.; Qian, Y.; Li, G.-J.; Tan, J. Preventive effects of the polysaccharide of Larimichthys crocea swim bladder on carbon tetrachloride (CCl4)-induced hepatic damage. Chin. J. Nat. Med. 2015, 13, 521–528. [Google Scholar] [CrossRef]
- Okada, T.; Hayashi, T.; Ikada, Y. Degradation of collagen suture in vitro and in vivo. Biomaterials 1992, 13, 448–454. [Google Scholar] [CrossRef]
- Bretaud, S.; Nauroy, P.; Malbouyres, M.; Ruggiero, F. Fishing for collagen function: About development, regeneration and disease. Sem. Cell Dev. Biol. 2019, 89, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Hickman, D.; Sims, T.J.; Miles, C.A.; Bailey, A.J.; de Mari, M.; Koopmans, M. Isinglass/collagen: Denaturation and functionality. J. Biotechnol. 2000, 79, 245–257. [Google Scholar] [CrossRef]
- Teng, B.; Petrie, P.R.; Smith, P.A.; Bindon, K.A. Comparison of water addition and early-harvest strategies to decrease alcohol concentration in Vitis vinifera cv. Shiraz wine: Impact on wine phenolics, tannin composition and colour properties. Aust. J. Grape Wine Res. 2020, 26, 158–171. [Google Scholar] [CrossRef]
- Sun, J.; Shi, X.; Zhang, H.; Xia, L.; Guo, Y.; Sun, X. Detection of moisture content in peanut kernels using hyperspectral imaging technology coupled with chemometrics. J. Food Process Eng. 2019, 42, e13263. [Google Scholar] [CrossRef]
- Kesava Reddy, G.; Enwemeka, C.S. A simplified method for the analysis of hydroxyproline in biological tissues. Clin. Biochem. 1996, 29, 225–229. [Google Scholar] [CrossRef]
- Heinrikson, R.L.; Meredith, S.C. Amino acid analysis by reverse-phase high-performance liquid chromatography: Precolumn derivatization with phenylisothiocyanate. Anal. Biochem. 1984, 136, 65–74. [Google Scholar] [CrossRef]
- Yu, X.; Guo, L.; Liu, M.; Cao, X.; Shang, S.; Liu, Z.; Huang, D.; Cao, Y.; Cui, F.; Tian, L. Callicarpa nudiflora loaded on chitosan-collagen/organomontmorillonite composite membrane for antibacterial activity of wound dressing. Int. J. Biol. Macromol. 2018, 120, 2279–2284. [Google Scholar] [CrossRef]
- Safandowska, M.; Pietrucha, K. Effect of fish collagen modification on its thermal and rheological properties. Int. J. Biol. Macromol. 2013, 53, 32–37. [Google Scholar] [CrossRef]
- Belbachir, K.; Noreen, R.; Gouspillou, G.; Petibois, C. Collagen types analysis and differentiation by FTIR spectroscopy. Anal. Bioanal. Chem. 2009, 395, 829–837. [Google Scholar] [CrossRef]
- Zhu, L.; Li, J.; Wang, Y.; Sun, X.; Li, B.; Poungchawanwong, S.; Hou, H. Structural feature and self-assembly properties of type II collagens from the cartilages of skate and sturgeon. Food Chem. 2020, 331, 127340. [Google Scholar] [CrossRef]
- Susanto, A.; Satari, M.H.; Abbas, B.; Koesoemowidodo, R.S.A.; Cahyanto, A. Fabrication and characterization of chitosan-collagen membrane from barramundi (Lates calcarifer) scales for guided tissue regeneration. Eur. J. Dent. 2019, 13, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Badii, F.; MacNaughtan, W.; Mitchell, J.R.; Farhat, I.A. The Effect of drying temperature on physical properties of thin gelatin films. Dry. Technol. 2014, 32, 30–38. [Google Scholar] [CrossRef]
- Gaurav Kumar, P.; Nidheesh, T.; Govindaraju, K.; Jyoti; Suresh, P.V. Enzymatic extraction and characterisation of a thermostable collagen from swim bladder of rohu (Labeo rohita). J. Sci. Food Agric. 2017, 97, 1451–1458. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Tian, Z.; Li, C.; Li, G. Thermal denaturation of fish collagen in solution: A calorimetric and kinetic analysis. Thermochim. Acta. 2014, 581, 32–40. [Google Scholar] [CrossRef]
- Chadefaux, C.; Le Hô, A.-S.; Bellot-Gurlet, L.; Reiche, I. Curve-fitting micro-ATR-FTIR studies of the amide I and II bands of type I collagen in archaeological bone materials. E-Preser. Sci. 2009, 6, 129–137. [Google Scholar] [CrossRef]
- Petibois, C.; Gouspillou, G.; Wehbe, K.; Delage, J.P.; Déléris, G. Analysis of type I and IV collagens by FT-IR spectroscopy and imaging for a molecular investigation of skeletal muscle connective tissue. Anal. Bioanal. Chem. 2006, 386, 1961–1966. [Google Scholar] [CrossRef]
- Şulea, D.; Micutz, M.; Albu, M.; Staicu, T.; Leca, M. Collagen-thuja tincture biomaterials for wound treatment. 2. Hydrogels and porous matrices. Rev. Roum. Chim. 2011, 56, 129–136. [Google Scholar] [CrossRef]
- Strasser, S.; Zink, A.; Janko, M.; Heckl, W.M.; Thalhammer, S. Structural investigations on native collagen type I fibrils using AFM. Biochem. Bioph. Res. Co. 2007, 354, 27–32. [Google Scholar] [CrossRef]
- Hashim, P.; Mohd Ridzwan, M.S.; Bakar, J.; Mat Hashim, D. Collagen in food and beverage industries. Int. Food Res. J. 2015, 22, 1–8. [Google Scholar] [CrossRef]
- Subhan, F.; Hussain, Z.; Tauseef, I.; Shehzad, A.; Wahid, F. A review on recent advances and applications of fish collagen. Crit. Rev. Food Sci. Nutr. 2021, 61, 1027–1037. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Zhang, H.; Yu, J.; Yu, C.; Chen, C.; Sun, Z.; Wang, S.; Wen, X. Effects of dietary selenium on growth performance, antioxidative status and tissue selenium deposition of juvenile Chu’s croaker (Nibea coibor). Aquaculture 2021, 536, 736439. [Google Scholar] [CrossRef]
- Gauza-Włodarczyk, M.; Kubisz, L.; Włodarczyk, D. Amino acid composition in determination of collagen origin and assessment of physical factors effects. Int. J. Biol. Macromol. 2017, 104, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Sinthusamran, S.; Benjakul, S.; Kishimura, H. Comparative study on molecular characteristics of acid soluble collagens from skin and swim bladder of seabass (Lates calcarifer). Food Chem. 2013, 138, 2435–2441. [Google Scholar] [CrossRef]
- Lin, F.; Rong, H.; Lin, J.; Yuan, Y.; Yu, J.; Yu, C.; You, C.; Wang, S.; Sun, Z.; Wen, X. Enhancement of collagen deposition in swim bladder of Chu’s croaker (Nibea coibor) by proline: View from in-vitro and in-vivo study. Aquaculture 2020, 523, 735175. [Google Scholar] [CrossRef]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef]
- Teng, B.; Jian, X.; Gao, Y.; Chen, W. Comparison of polyflavonoids in bayberry tanning effluent and commercial bayberry tannin: Prerequisite information for vegetable tanning effluent recycling. J. Clean. Prod. 2016, 112, 972–979. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FSB | 1-YSB | 5-YSB | 25-YSB | |
|---|---|---|---|---|
| Collagen (%) | 35.41 ± 0.70 b | 75.36 ± 0.47 a | 77.03 ± 13.38 a | 90.38 ± 3.02 a |
| Moisture (%) | 56.98 ± 1.04 a | 16.00 ± 0.46 b | 14.63 ± 0.57 b | 10.31 ± 0.20 c |
| Moisture and collagen in total (%) | 92.21 ± 0.43 a | 91.12 ± 0.16 a | 91.34 ± 13.19 a | 100.58 ± 2.95 a |
| Amino acid composition (mol %) | ||||
| Glycine | 33.50 ± 0.23 a | 35.34 ± 2.34 a | 33.48 ± 0.62 a | 33.54 ± 0.44 a |
| Hydroxyproline | 9.57 ± 1.54 a | 8.85 ± 2.46 a | 8.80 ± 0.21 a | 9.42 ± 0.25 a |
| Proline | 18.81 ± 0.53 a | 12.31 ± 0.57 b | 13.15 ± 0.28 b | 12.43 ± 0.02 b |
| Arginine | 1.26 ± 0.02 c | 2.11 ± 0.13 a | 1.81 ± 0.03 a b | 1.72 ± 0.08 b |
| Valine | 2.13 ± 0.03 b | 2.65 ± 0.08 a | 1.25 ± 0.02 c | 1.38 ± 0.002 c |
| Methionine | 1.74 ± 0.04 b | 2.09 ± 0.07 a | 1.45 ± 0.02 c | 0.54 ± 0.003 d |
| Isoleucine | 0.67 ± 0.03 b | 3.10 ± 0.08 a | 0.46 ± 0.01 c | 0.44 ± 0.01 c |
| Leucine | 2.55 ± 0.02 a | 0.94 ± 0.04 c | 2.46 ± 0.05 a | 2.26 ± 0.02 b |
| Phenylalanine | 1.59 ± 0.05 b | 1.79 ± 0.05 a | 1.31 ± 0.02 c | 1.27 ± 0.001 c |
| Lysine | 3.36 ± 0.08 a | 3.35 ± 0.09 a | 2.77 ± 0.06 b | 2.96 ± 0.01 b |
| Alanine | 4.74 ± 0.22 d | 11.80 ± 0.12 a | 10.06 ± 0.16 c | 10.86 ± 0.12 b |
| Tyrosine | 0.79 ± 0.03 b | 1.01 ± 0.03 a | 0.71 ± 0.02 b | 0.42 ± 0.02 c |
| Aspartic acid | 7.87 ± 0.99 a | 5.10 ± 0.08 b | 5.74 ± 0.69 a b | 5.95 ± 0.04 a b |
| Glutamate | 6.04 ± 0.28 b | 6.42 ± 1.23 b | 11.54 ± 0.23 a | 11.37 ± 0.002 a |
| Serine | 5.38 ± 0.13 a | 3.13 ± 0.17 b | 4.09 ± 0.03 b | 5.44 ± 0.50 a |
| Total imino acids 2 | 28.38 ± 1.01 a | 21.16 ± 1.90 b | 21.94 ± 0.49 b | 21.85 ± 0.27 b |
| FSB | 1-YSB | 5-YSB | 25-YSB | TDSB | |
|---|---|---|---|---|---|
| From maximum absorbance/wavenumber calculation | |||||
| AIII/A1450 | 1.06 ± 0.01 ab | 1.14 ± 0.06 a | 1.01 ± 0.01 b | 1.10 ± 0.02 a | 0.95 ± 0.01 c |
| νI–νII | 92.41 ± 0.35 b | 92.30 ± 3.43 b | 88.66 ± 0.25 b | 88.97 ± 1.64 b | 102.50 ± 1.01 a |
| From fitting by Gaussian equation | |||||
| α-helix (%) | 24.35 ± 0.56 ab | 21.31 ± 1.94 b | 23.34 ± 0.71 ab | 25.50 ± 0.96 a | 20.57 ± 2.12 b |
| β-sheet (%) | 37.55 ± 1.30 a | 42.24 ± 2.90 a | 38.49 ± 0.95 a | 37.57 ± 2.03 a | 30.42 ± 1.71 b |
| β-turn (%) | 15.65 ± 0.56 ab | 12.82 ± 0.94 c | 14.01 ± 0.41 bc | 13.79 ± 1.12 bc | 17.69 ± 0.52 a |
| Random coil (%) | 15.22 ± 0.22 b | 17.30 ± 2.73 ab | 14.45 ± 0.77 b | 14.19 ± 0.24 b | 21.29 ± 1.44 a |
| FSB | 1-YSB | 5-YSB | 25-YSB | TDSB | |
|---|---|---|---|---|---|
| Peak area (6° to 8°) | 1182.8 | 753.4 | 603.5 | 180.7 | 165.9 |
| Peak area (19° to 22°) | 5481.4 | 3567.5 | 2806.9 | 1713.4 | 927.6 |
| Diameter of triple helix | 1.31 | 1.14 | 1.22 | 1.20 | -- |
| FSB | 1-YSB | 5-YSB | 25-YSB | |
|---|---|---|---|---|
| D-Period (nm) | 70.82 ± 0.66 b | 76.93 ± 0.24 a | 75.30 ± 2.40 a | 70.70 ± 0.73 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, X.; Xie, L.; Zhang, X.; Lin, F.; Wen, X.; Teng, B. The Structural Characteristics of Collagen in Swim Bladders with 25-Year Sequence Aging: The Impact of Age. Appl. Sci. 2021, 11, 4578. https://doi.org/10.3390/app11104578
He X, Xie L, Zhang X, Lin F, Wen X, Teng B. The Structural Characteristics of Collagen in Swim Bladders with 25-Year Sequence Aging: The Impact of Age. Applied Sciences. 2021; 11(10):4578. https://doi.org/10.3390/app11104578
Chicago/Turabian StyleHe, Xiaofeng, Liling Xie, Xiaoshan Zhang, Fan Lin, Xiaobo Wen, and Bo Teng. 2021. "The Structural Characteristics of Collagen in Swim Bladders with 25-Year Sequence Aging: The Impact of Age" Applied Sciences 11, no. 10: 4578. https://doi.org/10.3390/app11104578
APA StyleHe, X., Xie, L., Zhang, X., Lin, F., Wen, X., & Teng, B. (2021). The Structural Characteristics of Collagen in Swim Bladders with 25-Year Sequence Aging: The Impact of Age. Applied Sciences, 11(10), 4578. https://doi.org/10.3390/app11104578