
Mini-Review: Potential of Diatom-Derived Silica for Biomedical Applications





Abstract
1. Introduction
2. Silicon Capture from the Environment, Transport and Storage, Frustule Formation
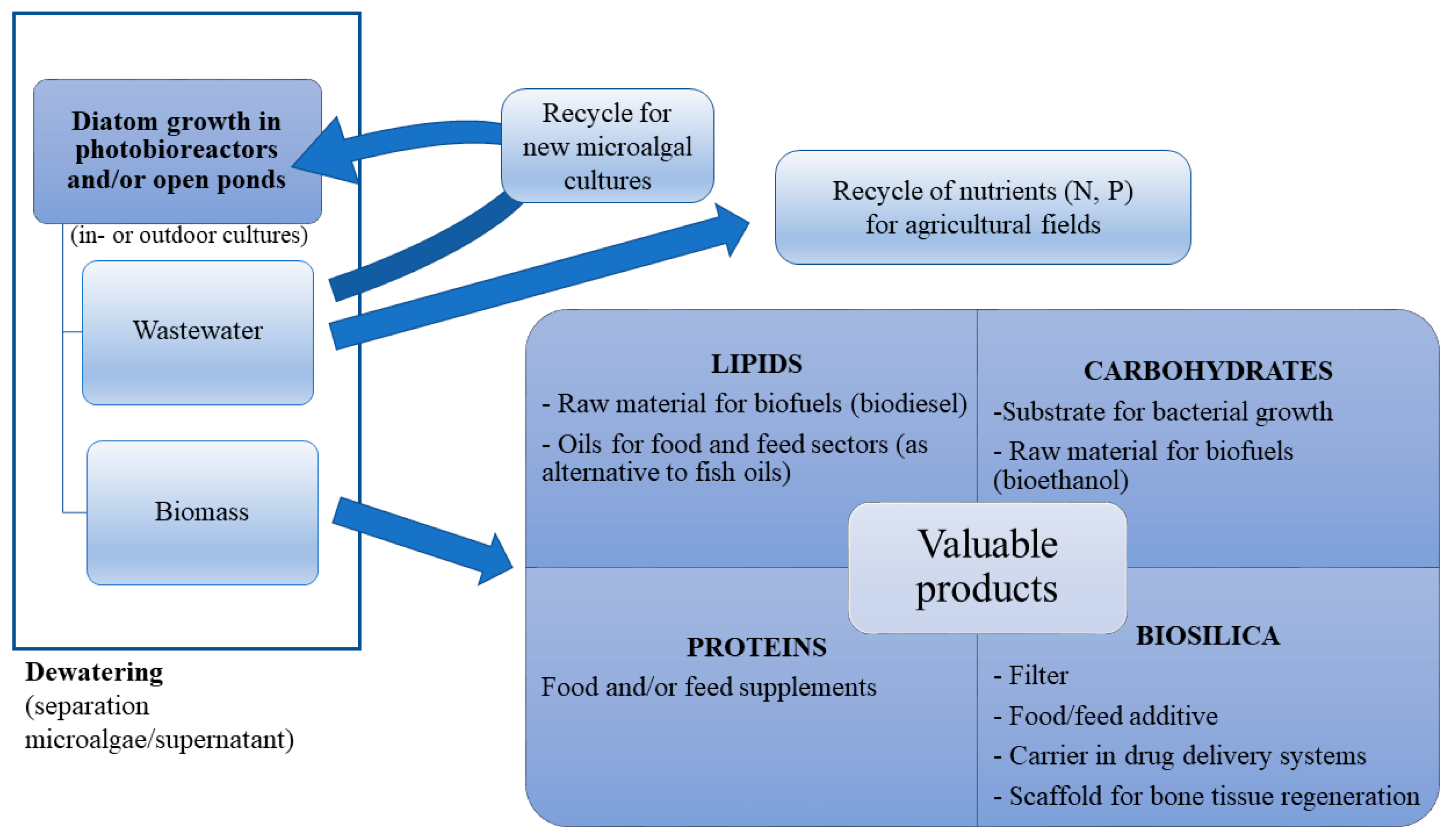
3. Diatom Biosilica Sources
4. Frustule Cleaning/Purification: Main Techniques and Technical Issues
5. Silica for Biomedical Applications: Advantages
5.1. Surface Functionalization for Drug Loading and for Biosensing Chips for Biomedical Applications
5.2. Biocompatibility
5.3. Employment of Genetically Engineered Diatom Frustules for Protein Immobilization
5.4. Availability of Biosilica Feedstocks
6. Silica for Biomedical Applications: Bottlenecks
7. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mann, D.G.; Vanormelingen, P. An inordinate fondness? The number, distributions, and origins of diatom species. J. Eukaryot. Microbiol. 2013, 60, 414–420. [Google Scholar] [CrossRef]
- Armbrust, E.V.; Berges, J.A.; Bowler, C.; Green, B.R.; Martinez, D.; Putnam, N.H.; Zhou, S.G.; Allen, A.E.; Apt, K.E.; Bechner, M.; et al. The genome of the diatom Thalassiosira pseudonana: Ecology, evolution, and metabolism. Science 2004, 306, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Bowler, C.; Allen, A.E.; Badger, J.H.; Grimwood, J.; Jabbari, K.; Kuo, A.; Maheswari, U.; Martens, C.; Maumus, F.; Otillar, R.P.; et al. The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 2008, 456, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Lommer, M.; Specht, M.; Roy, A.S.; Kraemer, L.; Andreson, R.; Gutowska, M.A.; Wolf, J.; Bergner, S.V.; Schilhabel, M.B.; Klostermeier, U.C.; et al. Genome and low-iron response of an oceanic diatom adapted to chronic iron limitation. Genome Biol. 2012, 13. [Google Scholar] [CrossRef] [PubMed]
- Galachyants, Y.P.; Zakharova, Y.R.; Petrova, D.P.; Morozov, A.A.; Sidorov, I.A.; Marchenkov, A.M.; Logacheva, M.D.; Markelov, M.L.; Khabudaev, K.V.; Likhoshway, Y.V.; et al. Sequencing of the complete genome of an araphid pennate diatom Synedra acus subsp. radians from Lake Baikal. Dokl. Biochem. Biophys. 2015, 461, 84–88. [Google Scholar] [CrossRef]
- Tanaka, T.; Maeda, Y.; Veluchamy, A.; Tanaka, M.; Abida, H.; Marechal, E.; Bowler, C.; Muto, M.; Sunaga, Y.; Tanaka, M.; et al. Oil accumulation by the oleaginous diatom Fistulifera solaris as revealed by the genome and transcriptome. Plant Cell 2015, 27, 162–176. [Google Scholar] [CrossRef]
- Mock, T.; Otillar, R.P.; Strauss, J.; McMullan, M.; Paajanen, P.; Schmutz, J.; Salamov, A.; Sanges, R.; Toseland, A.; Ward, B.J.; et al. Evolutionary genomics of the cold-adapted diatom Fragilariopsis cylindrus. Nature 2017, 541, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Osuna-Cruz, C.M.; Bilcke, G.; Vancaester, E.; De Decker, S.; Bones, A.M.; Winge, P.; Poulsen, N.; Bulankova, P.; Verhelst, B.; Audoor, S.; et al. The Seminavis robusta genome provides insights into the evolutionary adaptations of benthic diatoms. Nat. Commun. 2020, 11, 3320. [Google Scholar] [CrossRef]
- Pendergrass, A.; Roberts, W.; Ruck, E.C.; Lewis, J.A.; Alverson, A.J. The genome of a nonphotosynthetic diatom provides insights into the metabolic shift to heterotrophy and constraints on the loss of photosynthesis. BioRxiv 2020. [Google Scholar] [CrossRef]
- Mourelle, M.; Gómez, C.; Legido, J. The Potential Use of Marine Microalgae and Cyanobacteria in Cosmetics and Thalassotherapy. Cosmetics 2017, 4, 46. [Google Scholar] [CrossRef]
- Cui, Y.; Thomas-Hall, S.R.; Schenk, P.M. Phaeodactylum tricornutum microalgae as a rich source of omega-3 oil: Progress in lipid induction techniques towards industry adoption. Food Chem. 2019, 297, 124937. [Google Scholar] [CrossRef]
- Pudney, A.; Gandini, C.; Economou, C.K.; Smith, R.; Goddard, P.; Napier, J.A.; Spicer, A.; Sayanova, O. Multifunctionalizing the marine diatom Phaeodactylum tricornutum for sustainable co-production of omega-3 long chain polyunsaturated fatty acids and recombinant phytase. Sci. Rep. 2019, 9, 11444. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, A.; Melchor-Martinez, E.M.; Saxena, A.; Kapoor, N.; Singh, K.J.; Saldarriaga-Hernandez, S.; Parra-Saldivar, R.; Iqbal, H.M.N. Therapeutic attributes and applied aspects of biological macromolecules (polypeptides, fucoxanthin, sterols, fatty acids, polysaccharides, and polyphenols) from diatoms—A review. Int. J. Biol. Macromol. 2021, 171, 398–413. [Google Scholar] [CrossRef]
- Grzesik, M.; Romanowska-Duda, Z.; Piotrowski, K.; Janas, R. Diatoms (Bacillariophyceae) as an effective base of a new generation of organic fertilizers. Przemys Chem. 2015, 94, 391–396. [Google Scholar] [CrossRef]
- Hedayatkhah, A.; Cretoiu, M.S.; Emtiazi, G.; Stal, L.J.; Bolhuis, H. Bioremediation of chromium contaminated water by diatoms with concomitant lipid accumulation for biofuel production. J. Environ. Manag. 2018, 227, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Mojiri, A.; Baharlooeian, M.; Zahed, M.A. The Potential of Chaetoceros muelleri in Bioremediation of Antibiotics: Performance and Optimization. Int. J. Environ. Res. Public Health 2021, 18, 977. [Google Scholar] [CrossRef]
- Lomora, M.; Shumate, D.; Rahman, A.A.; Pandit, A. Therapeutic applications of phytoplankton, with an emphasis on diatoms and coccolithophores. Adv. Ther. 2019, 2, 1800099. [Google Scholar] [CrossRef]
- Uthappa, U.T.; Brahmkhatri, V.; Sriram, G.; Jung, H.Y.; Yu, J.; Kurkuri, N.; Aminabhavi, T.M.; Altalhi, T.; Neelgund, G.M.; Kurkuri, M.D. Nature engineered diatom biosilica as drug delivery systems. J. Control. Release 2018, 281, 70–83. [Google Scholar] [CrossRef]
- Uthappa, U.T.; Sriram, G.; Arvind, O.R.; Kumar, S.; Ho Young, J.; Neelgund, G.M.; Losic, D.; Kurkuri, M.D. Engineering MIL-100(Fe) on 3D porous natural diatoms as a versatile high performing platform for controlled isoniazid drug release, Fenton’s catalysis for malachite green dye degradation and environmental adsorbents for Pb2+ removal and dyes. Appl. Surf. Sci. 2020, 528, 146974. [Google Scholar] [CrossRef]
- Ruggiero, I.; Terracciano, M.; Martucci, N.M.; De Stefano, L.; Migliaccio, N.; Tate, R.; Rendina, I.; Arcari, P.; Lamberti, A.; Rea, I. Diatomite silica nanoparticles for drug delivery. Nanoscale Res. Lett. 2014, 9, 7. [Google Scholar] [CrossRef]
- Dalgic, A.D.; Atila, D.; Karatas, A.; Tezcaner, A.; Keskin, D. Diatom shell incorporated PHBV/PCL-pullulan co-electrospun scaffold for bone tissue engineering. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 100, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Squire, K.; Li, E.; LeDuff, P.; Rorrer, G.L.; Tang, S.; Chen, B.; McKay, C.P.; Navarro-Gonzalez, R.; Wang, A.X. Chemical and Biological Sensing Using Diatom Photonic Crystal Biosilica With In-Situ Growth Plasmonic Nanoparticles. IEEE Trans. Nanobiosci. 2016, 15, 828–834. [Google Scholar] [CrossRef] [PubMed]
- Albert, K.; Huang, X.C.; Hsu, H.Y. Bio-templated silica composites for next-generation biomedical applications. Adv. Colloid. Interface Sci. 2017, 249, 272–289. [Google Scholar] [CrossRef] [PubMed]
- Terracciano, M.; De Stefano, L.; Rea, I. Diatoms green nanotechnology for biosilica-based drug delivery systems. Pharmaceutics 2018, 10, 242. [Google Scholar] [CrossRef] [PubMed]
- Delasoie, J.; Zobi, F. Natural diatom biosilica as microshuttles in drug delivery systems. Pharmaceutics 2019, 11, 537. [Google Scholar] [CrossRef]
- Tramontano, C.; Chianese, G.; Terracciano, M.; de Stefano, L.; Rea, I. Nanostructured biosilica of diatoms: From water world to biomedical applications. Appl. Sci. 2020, 10, 6811. [Google Scholar] [CrossRef]
- Lewin, J.C. Silicon metabolism in diatoms. III. Respiration and silicon uptake in Navicula pelliculosa. J. Gen. Physiol. 1955, 39, 1–10. [Google Scholar] [CrossRef]
- Annenkov, V.V.; Danilovtseva, E.N.; Pal’shin, V.A.; Ol’ga, N.V.; Zelinskiy, S.N.; Krishnan, U.M. Silicic acid condensation under the influence of water-soluble polymers: From biology to new materials. RSC Adv. 2017, 7, 20995–21027. [Google Scholar] [CrossRef]
- Thamatrakoln, K.; Hildebrand, M. Silicon uptake in diatoms revisited: A model for saturable and nonsaturable uptake kinetics and the role of silicon transporters. Plant Physiol. 2008, 146, 1397–1407. [Google Scholar] [CrossRef]
- Hildebrand, M.; Dahlin, K.; Volcani, B.E. Characterization of a silicon transporter gene family in Cylindrotheca fusiformis: Sequences, expression analysis, and identification of homologs in other diatoms. Mol. Gen. Genet. 1998, 260, 480–486. [Google Scholar] [CrossRef]
- Hildebrand, M. Biological processing of nanostructured silica in diatoms. Prog. Org. Coat. 2003, 47, 256–266. [Google Scholar] [CrossRef]
- Marchenkov, A.M.; Petrova, D.P.; Morozov, A.A.; Zakharova, Y.R.; Grachev, M.A.; Bondar, A.A. A family of silicon transporter structural genes in a pennate diatom Synedra ulna subsp. danica (Kutz.) Skabitsch. PLoS ONE 2018, 13, e0203161. [Google Scholar] [CrossRef]
- Hildebrand, M.; Volcani, B.E.; Gassmann, W.; Schroeder, J.I. A gene family of silicon transporters. Nature 1997, 385, 688–689. [Google Scholar] [CrossRef]
- Durkin, C.A.; Koester, J.A.; Bender, S.J.; Armbrust, E.V. The evolution of silicon transporters in diatoms. J. Phycol. 2016, 52, 716–731. [Google Scholar] [CrossRef]
- Brzezinski, M.A.; Conley, D.J. Silicon deposition during the cell-cycle of Thalassiosira weissflogii (Bacillariophyceae) determined using dual Rhodamine-123 and propidium iodide staining. J. Phycol. 1994, 30, 45–55. [Google Scholar] [CrossRef]
- Taylor, N.J. Silica incorporation in the diatom Cosinodiscus granii as affected by light intensity. Br. Phycol. J. 1985, 20, 365–374. [Google Scholar] [CrossRef]
- Martin-Jezequel, V.; Hildebrand, M.; Brzezinski, M.A. Silicon metabolism in diatoms: Implications for growth. J. Phycol. 2000, 36, 821–840. [Google Scholar] [CrossRef]
- Hildebrand, M. Diatoms, Biomineralization Processes, and Genomics. Chem. Rev. 2008, 108, 4855–4874. [Google Scholar] [CrossRef]
- Crawford, R.; Schmid, A.M. Ultrastructure of silica deposition. In Biomineralization in Lower Plants and Animals; Leadbeater, B.S., Riding, R., Eds.; Oxford University Press: Oxford, UK, 1986; Volume 30, pp. 291–314. [Google Scholar]
- Schmidt, A.-M. Aspects of morphogenesis and function of diatom cell walls with implications for taxonomy. Protoplasma 1994, 181, 43–60. [Google Scholar] [CrossRef]
- Li, C.W.; Volcani, B.E. Aspects of silicification in wall morphogenesis of diatoms. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1984, 304, 519–528. [Google Scholar] [CrossRef]
- Tesson, B.; Hildebrand, M. Extensive and intimate association of the cytoskeleton with forming silica in diatoms: Control over patterning on the meso- and micro-scale. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Sumper, M.; Kroger, N. Silica formation in diatoms: The function of long-chain polyamines and silaffins. J. Mater. Chem. 2004, 14, 2059–2065. [Google Scholar] [CrossRef]
- Pahl, S.L.; Lewis, D.M.; Chen, F.; King, K.D. Heterotrophic growth and nutritional aspects of the diatom Cyclotella cryptica (Bacillariophyceae): Effect of some environmental factors. J. Biosci. Bioeng. 2010, 109, 235–239. [Google Scholar] [CrossRef]
- Pahl, S.L.; Lewis, D.M.; Chen, F.; King, K.D. Growth dynamics and the proximate biochemical composition and fatty acid profile of the heterotrophically grown diatom Cyclotella Cryptica. J. Appl. Phycol. 2010, 22, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Karmakar, R.; Das, B.; Diba, F.; Razu, M.H. Recent advances in microalgal biotechnology. In Heterotrophic Growth of Micro Algae; Jin, L., Zheng, S., Henri, G., Eds.; OMICS Group eBooks: Foster City, CA, USA, 2016; pp. 1–18. [Google Scholar]
- Mao, X.M.; Chen, S.H.Y.; Lu, X.; Yu, J.F.; Liu, B. High silicate concentration facilitates fucoxanthin and eicosapentaenoic acid (EPA) production under heterotrophic condition in the marine diatom Nitzschia laevis. Algal Res. Biomass Biofuels Bioprod. 2020, 52. [Google Scholar] [CrossRef]
- Chen, G.Q.; Chen, F. Growing phototrophic cells without light. Biotechnol. Lett. 2006, 28, 607–616. [Google Scholar] [CrossRef]
- Morales-Sanchez, D.; Martinez-Rodriguez, O.A.; Kyndt, J.; Martinez, A. Heterotrophic growth of microalgae: Metabolic aspects. World J. Microbiol. Biotechnol. 2015, 31, 1–9. [Google Scholar] [CrossRef]
- D’Ippolito, G.; Sardo, A.; Paris, D.; Vella, F.M.; Adelfi, M.G.; Botte, P.; Gallo, C.; Fontana, A. Potential of lipid metabolism in marine diatoms for biofuel production. Biotechnol. Biofuels 2015, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Botte, P.; D’Ippolito, G.; Gallo, C.; Sardo, A.; Fontana, A. Combined exploitation of CO2 and nutrient replenishment for increasing biomass and lipid productivity of the marine diatoms Thalassiosira weissflogii and Cyclotella cryptica. J. Appl. Phycol. 2017, 30, 243–251. [Google Scholar] [CrossRef]
- Orefice, I.; Musella, M.; Smerilli, A.; Sansone, C.; Chandrasekaran, R.; Corato, F.; Brunet, C. Role of nutrient concentrations and water movement on diatom’s productivity in culture. Sci. Rep. 2019, 9, 1479. [Google Scholar] [CrossRef]
- Lutyński, M.; Sakiewicz, P.; Lutyńska, S. Characterization of diatomaceous earth and halloysite resources of poland. Minerals 2019, 9, 670. [Google Scholar] [CrossRef]
- Qin, T.; Gutu, T.; Jiao, J.; Chang, C.H.; Rorrer, G.L. Photoluminescence of silica nanostructures from bioreactor culture of marine diatom Nitzschia frustulum. J. Nanosci. Nanotechnol. 2008, 8, 2392–2398. [Google Scholar] [CrossRef]
- Wang, Y.; Cai, J.; Jiang, Y.G.; Jiang, X.G.; Zhang, D.Y. Preparation of biosilica structures from frustules of diatoms and their applications: Current state and perspectives. Appl. Microbiol. Biotechnol. 2013, 97, 453–460. [Google Scholar] [CrossRef]
- Qi, Y.R.; Wang, X.; Cheng, J.J. Preparation and characteristics of biosilica derived from marine diatom biomass of Nitzschia closterium and Thalassiosira. Chin. J. Oceanol. Limnol. 2017, 35, 668–680. [Google Scholar] [CrossRef]
- Horn, M.G.; Robinson, R.S.; Rynearson, T.A.; Sigman, D.M. Nitrogen isotopic relationship between diatom-bound and bulk organic matter of cultured polar diatoms. Paleoceanography 2011, 26. [Google Scholar] [CrossRef]
- Mejia, L.M.; Isensee, K.; Mendez-Vicente, A.; Pisonero, J.; Shimizu, N.; Gonzalez, C.; Monteleone, B.; Stoll, H. B content and Si/C ratios from cultured diatoms (Thalassiosira pseudonana and Thalassiosira weissflogii): Relationship to seawater pH and diatom carbon acquisition. Geochim. Cosmochim. Acta 2013, 123, 322–337. [Google Scholar] [CrossRef]
- Jiang, W.K.; Luo, S.P.; Liu, P.W.; Deng, X.Y.; Jing, Y.; Bai, C.Y.; Li, J.B. Purification of biosilica from living diatoms by a two-step acid cleaning and baking method. J. Appl. Phycol. 2014, 26, 1511–1518. [Google Scholar] [CrossRef]
- Umemura, K.; Noguchi, Y.; Ichinose, T.; Hirose, Y.; Kuroda, R.; Mayama, S. Diatom Cells Grown and Baked on a Functionalized Mica Surface. J. Biol. Phys. 2008, 34, 189–196. [Google Scholar] [CrossRef]
- Watanabe, T.; Kodama, Y.; Mayama, S. Application of a novel cleaning method using low-temperature plasma on tidal flat diatoms with heterovalvy or delicate frustule structure. Proc. Acad. Nat. Sci. Phila. 2010, 160, 83–87. [Google Scholar] [CrossRef]
- Saad, E.M.; Pickering, R.A.; Shoji, K.; Hossain, M.I.; Glover, T.G.; Krause, J.W.; Tang, Y.Z. Effect of cleaning methods on the dissolution of diatom frustules. Mar. Chem. 2020, 224. [Google Scholar] [CrossRef]
- Jeffryes, C.; Solanki, R.; Rangineni, Y.; Wang, W.; Chang, C.H.; Rorrer, G.L. Electroluminescence and photoluminescence from nanostructured diatom frustules containing metabolically inserted germanium. Adv. Mater. 2008, 20, 2633–2637. [Google Scholar] [CrossRef]
- Townley, H.E.; Parker, A.R.; White-Cooper, H. Exploitation of diatom frustules for nanotechnology: Tethering active biomolecules. Adv. Funct. Mater. 2008, 18, 369–374. [Google Scholar] [CrossRef]
- Abramson, L.; Wirick, S.; Lee, C.; Jacobsen, C.; Brandes, J.A. The use of soft X-ray spectromicroscopy to investigate the distribution and composition of organic matter in a diatom frustule and a biomimetic analog. Deep Sea Res. Part II Top. Stud. Oceanogr. 2009, 56, 1369–1380. [Google Scholar] [CrossRef]
- Lin, K.C.; Kunduru, V.; Bothara, M.; Rege, K.; Prasad, S.; Ramakrishna, B.L. Biogenic nanoporous silica-based sensor for enhanced electrochemical detection of cardiovascular biomarkers proteins. Biosens. Bioelectron. 2010, 25, 2336–2342. [Google Scholar] [CrossRef] [PubMed]
- Bariana, M.; Aw, M.S.; Kurkuri, M.; Losic, D. Tuning drug loading and release properties of diatom silica microparticles by surface modifications. Int. J. Pharm. 2013, 443, 230–241. [Google Scholar] [CrossRef]
- Van Eynde, E.; Lenaerts, B.; Tytgat, T.; Verbruggen, S.W.; Hauchecorne, B.; Blust, R.; Lenaerts, S. Effect of pretreatment and temperature on the properties of Pinnularia biosilica frustules. RSC Adv. 2014, 4, 56200–56206. [Google Scholar] [CrossRef]
- Lettieri, S.; Setaro, A.; De Stefano, L.; De Stefano, M.; Maddalena, P. The gas-detection properties of light-emitting diatoms. Adv. Funct. Mater. 2008, 18, 1257–1264. [Google Scholar] [CrossRef]
- De Stefano, L.; Rendina, I.; De Stefano, M.; Bismuto, A.; Maddalena, P. Marine diatoms as optical chemical sensors. Appl. Phys. Lett. 2005, 87. [Google Scholar] [CrossRef]
- De Stefano, L.; Rotiroti, L.; De Stefano, M.; Lamberti, A.; Lettieri, S.; Setaro, A.; Maddalena, P. Marine diatoms as optical biosensors. Biosens. Bioelectron. 2009, 24, 1580–1584. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, D.Y.; Cai, J.; Pan, J.F.; Chen, M.L.; Li, A.B.; Jiang, Y.G. Biosilica structures obtained from Nitzschia, Ditylum, Skeletonema, and Coscinodiscus diatom by a filtration-aided acid cleaning method. Appl. Microbiol. Biotechnol. 2012, 95, 1165–1178. [Google Scholar] [CrossRef]
- Delalat, B.; Sheppard, V.C.; Rasi Ghaemi, S.; Rao, S.; Prestidge, C.A.; McPhee, G.; Rogers, M.L.; Donoghue, J.F.; Pillay, V.; Johns, T.G.; et al. Targeted drug delivery using genetically engineered diatom biosilica. Nat. Commun. 2015, 6, 8791. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, M.A.A.; Pack, S.P. Biomimetic and bioinspired silicifications: Recent advances for biomaterial design and applications. Acta Biomater. 2021, 120, 38–56. [Google Scholar] [CrossRef] [PubMed]
- Vasani, R.B.; Losic, D.; Cavallaro, A.; Voelcker, N.H. Fabrication of stimulus-responsive diatom biosilica microcapsules for antibiotic drug delivery. J. Mater. Chem. B 2015, 3, 4325–4329. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.; Hanini, A.; Shah, A.; Patel, D.; Patel, S.; Bhatt, P.; Pathak, V.Y. Surface modification of nanoparticles for targeted drug delivery. In Surface Modification of Nanoparticles for Targeted Drug Delivery; Pathak, V.Y., Ed.; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Bao, Z.; Weatherspoon, M.R.; Shian, S.; Cai, Y.; Graham, P.D.; Allan, S.M.; Ahmad, G.; Dickerson, M.B.; Church, B.C.; Kang, Z.; et al. Chemical reduction of three-dimensional silica micro-assemblies into microporous silicon replicas. Nature 2007, 446, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Losic, D.; Yu, Y.; Aw, M.S.; Simovic, S.; Thierry, B.; Addai-Mensah, J. Surface functionalisation of diatoms with dopamine modified iron-oxide nanoparticles: Toward magnetically guided drug microcarriers with biologically derived morphologies. Chem. Commun. 2010, 46, 6323–6325. [Google Scholar] [CrossRef]
- Jantschke, A.; Fischer, C.; Hensel, R.; Braun, H.G.; Brunner, E. Directed assembly of nanoparticles to isolated diatom valves using the non-wetting characteristics after pyrolysis. Nanoscale 2014, 6, 11637–11645. [Google Scholar] [CrossRef]
- Diab, M.; Mokari, T. Bioinspired hierarchical porous structures for engineering advanced functional inorganic materials. Adv. Mater. 2018, 30, e1706349. [Google Scholar] [CrossRef]
- Aw, M.S.; Simovic, S.; Yu, Y.; Addai-Mensah, J.; Losic, D. Porous silica microshells from diatoms as biocarrier for drug delivery applications. Powder Technol. 2012, 223, 52–58. [Google Scholar] [CrossRef]
- Kumeria, T.; Bariana, M.; Altalhi, T.; Kurkuri, M.; Gibson, C.T.; Yang, W.; Losic, D. Graphene oxide decorated diatom silica particles as new nano-hybrids: Towards smart natural drug microcarriers. J. Mater. Chem. B 2013, 1, 6302–6311. [Google Scholar] [CrossRef]
- Delasoie, J.; Rossier, J.; Haeni, L.; Rothen-Rutishauser, B.; Zobi, F. Slow-targeted release of a ruthenium anticancer agent from vitamin B12 functionalized marine diatom microalgae. Dalton Trans. 2018, 47, 17221–17232. [Google Scholar] [CrossRef]
- Maher, S.; Santos, A.; Kumeria, T.; Kaur, G.; Lambert, M.; Forward, P.; Evdokiou, A.; Losic, D. Multifunctional microspherical magnetic and pH responsive carriers for combination anticancer therapy engineered by droplet-based microfluidics. J. Mater. Chem. B 2017, 5, 4097–4109. [Google Scholar] [CrossRef] [PubMed]
- Milovic, M.; Simovic, S.; Losic, D.; Dashevskiy, A.; Ibric, S. Solid self-emulsifying phospholipid suspension (SSEPS) with diatom as a drug carrier. Eur. J. Pharm. Sci. 2014, 63, 226–232. [Google Scholar] [CrossRef]
- Singh, R.; Khan, M.J.; Rane, J.; Gajbhiye, A.; Vinayak, V.; Joshi, K.B. Biofabrication of Diatom Surface by Tyrosine-Metal Complexes: Smart Microcontainers to Inhibit Bacterial Growth. Chemistryselect 2020, 5, 3091–3097. [Google Scholar] [CrossRef]
- Esfandyari, J.; Shojaedin-Givi, B.; Hashemzadeh, H.; Mozafari-Nia, M.; Vaezi, Z.; Naderi-Manesh, H. Capture and detection of rare cancer cells in blood by intrinsic fluorescence of a novel functionalized diatom. Photodiagn. Photodyn. Ther. 2020, 30, 101753. [Google Scholar] [CrossRef] [PubMed]
- De Stefano, L.; Lamberti, A.; Rotiroti, L.; De Stefano, M. Interfacing the nanostructured biosilica microshells of the marine diatom Coscinodiscus wailesii with biological matter. Acta Biomater. 2008, 4, 126–130. [Google Scholar] [CrossRef]
- Gale, D.K.; Gutu, T.; Jiao, J.; Chang, C.-H.; Rorrer, G.L. Photoluminescence Detection of Biomolecules by Antibody-Functionalized Diatom Biosilica. Adv. Funct. Mater. 2009, 19, 926–933. [Google Scholar] [CrossRef]
- Cicco, S.R.; Vona, D.; Gristina, R.; Sardella, E.; Ragni, R.; Lo Presti, M.; Farinola, G.M. Biosilica from Living Diatoms: Investigations on Biocompatibility of Bare and Chemically Modified Thalassiosira weissflogii Silica Shells. Bioengineering 2016, 3, 35. [Google Scholar] [CrossRef]
- Aw, M.S.; Bariana, M.; Yu, Y.; Addai-Mensah, J.; Losic, D. Surface-functionalized diatom microcapsules for drug delivery of water-insoluble drugs. J. Biomater. Appl. 2013, 28, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Martucci, N.M.; Migliaccio, N.; Ruggiero, I.; Albano, F.; Cali, G.; Romano, S.; Terracciano, M.; Rea, I.; Arcari, P.; Lamberti, A. Nanoparticle-based strategy for personalized B-cell lymphoma therapy. Int. J. Nanomed. 2016, 11, 6089–6101. [Google Scholar] [CrossRef]
- Rea, I.; Martucci, N.M.; De Stefano, L.; Ruggiero, I.; Terracciano, M.; Dardano, P.; Migliaccio, N.; Arcari, P.; Tate, R.; Rendina, I.; et al. Diatomite biosilica nanocarriers for siRNA transport inside cancer cells. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 3393–3403. [Google Scholar] [CrossRef]
- Zhang, H.; Shahbazi, M.A.; Makila, E.M.; da Silva, T.H.; Reis, R.L.; Salonen, J.J.; Hirvonen, J.T.; Santos, H.A. Diatom silica microparticles for sustained release and permeation enhancement following oral delivery of prednisone and mesalamine. Biomaterials 2013, 34, 9210–9219. [Google Scholar] [CrossRef] [PubMed]
- Terracciano, M.; Shahbazi, M.A.; Correia, A.; Rea, I.; Lamberti, A.; De Stefano, L.; Santos, H.A. Surface bioengineering of diatomite based nanovectors for efficient intracellular uptake and drug delivery. Nanoscale 2015, 7, 20063–20074. [Google Scholar] [CrossRef]
- Amoda, A.; Borkiewicz, L.; Rivero-Müller, A.; Alam, P. Sintered nanoporous biosilica diatom frustules as high efficiency cell-growth and bone-mineralisation platforms. Mater. Today Commun. 2020, 24, 100923. [Google Scholar] [CrossRef]
- Terracciano, M.; De Stefano, L.; Tortiglione, C.; Tino, A.; Rea, I. In Vivo Toxicity Assessment of Hybrid Diatomite Nanovectors Using Hydra vulgaris as a Model System. Adv. Biosyst. 2019, 3, e1800247. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, N.; Berne, C.; Spain, J.; Kroger, N. Silica immobilization of an enzyme through genetic engineering of the diatom Thalassiosira pseudonana. Angew. Chem. Int. Ed. 2007, 46, 1843–1846. [Google Scholar] [CrossRef]
- Kumari, E.; Görlich, S.; Poulsen, N.; Kröger, N. Genetically Programmed Regioselective Immobilization of Enzymes in Biosilica Microparticles. Adv. Funct. Mater. 2020, 30, 2000442. [Google Scholar] [CrossRef]
- Ford, N.R.; Hecht, K.A.; Hu, D.H.; Orr, G.; Xiong, Y.J.; Squier, T.C.; Rorrer, G.L.; Roesijadi, G. Antigen Binding and Site-Directed Labeling of Biosilica-Immobilized Fusion Proteins Expressed in Diatoms. ACS Synth. Biol. 2016, 5, 193–199. [Google Scholar] [CrossRef]
- Vona, D.; Urbano, L.; Bonifacio, M.A.; De Giglio, E.; Cometa, S.; Mattioli-Belmonte, M.; Palumbo, F.; Ragni, R.; Cicco, S.R.; Farinola, G.M. Data from two different culture conditions of Thalassiosira weissflogii diatom and from cleaning procedures for obtaining monodisperse nanostructured biosilica. Data Brief 2016, 8, 312–319. [Google Scholar] [CrossRef]
- Saxena, A.; Tiwari, A.; Kaushik, R.; Iqbal, H.M.N.; Parra-Saldivar, R. Diatoms recovery from wastewater: Overview from an ecological and economic perspective. J. Water Process Eng. 2021, 39. [Google Scholar] [CrossRef]
- Diab, R.; Canilho, N.; Pavel, I.A.; Haffner, F.B.; Girardon, M.; Pasc, A. Silica-based systems for oral delivery of drugs, macromolecules and cells. Adv. Colloid Interface Sci. 2017, 249, 346–362. [Google Scholar] [CrossRef]
- Cauda, V.; Schlossbauer, A.; Bein, T. Bio-degradation study of colloidal mesoporous silica nanoparticles: Effect of surface functionalization with organo-silanes and poly(ethylene glycol). Microporous Mesoporous Mater. 2010, 132, 60–71. [Google Scholar] [CrossRef]
- Anderson, S.H.C.; Elliott, H.; Wallis, D.J.; Canham, L.T.; Powell, J.J. Dissolution of different forms of partially porous silicon wafers under simulated physiological conditions. Phys. Status Solidi A Appl. Mater. Sci. 2003, 197, 331–335. [Google Scholar] [CrossRef]
- Popplewell, J.F.; King, S.J.; Day, J.P.; Ackrill, P.; Fifield, L.K.; Cresswell, R.G.; Di Tada, M.L.; Liu, K. Kinetics of uptake and elimination of silicic acid by a human subject: A novel application of Si-32 and accelerator mass spectrometry. J. Inorg. Biochem. 1998, 69, 177–180. [Google Scholar] [CrossRef]
- Zhang, Y.; Hsu, B.Y.W.; Ren, C.L.; Li, X.; Wang, J. Silica-based nanocapsules: Synthesis, structure control and biomedical applications. Chem. Soc. Rev. 2015, 44, 315–335. [Google Scholar] [CrossRef]
- Wallace, A.K.; Chanut, N.; Voigt, C.A. Silica Nanostructures Produced Using Diatom Peptides with Designed Post-Translational Modifications. Adv. Funct. Mater. 2020, 30. [Google Scholar] [CrossRef]
- Rogato, A.; De Tommasi, E. Physical, Chemical, and Genetic Techniques for Diatom Frustule Modification: Applications in Nanotechnology. Appl. Sci. 2020, 10, 8738. [Google Scholar] [CrossRef]
- Sun, Z.; Zhou, Z.; Li, J.; Feng, C.; Chen, X.; Zhang, Y. Industrialized System for Producing Bio-Silicon, Has Centrifugal Shaft That is Connected to Barrel Structure of Inner Barrel, and Flocculation Tank Whose Outlet is Connected to Algae-Liquid Separation Tank. China Patent CN110616139-A, 27 December 2019. CN110616139-B, 28 August 2020. [Google Scholar]
- Kaya, M.; Bilican, I.; Mujtaba, M.; Sargin, I.; Haskoylu, M.E.; Oner, E.T.; Zheng, K.; Boccaccini, A.R.; Cansaran-Duman, D.; Onses, M.S.; et al. Sponge-derived natural bioactive glass microspheres with self-assembled surface channel arrays opening into a hollow core for bone tissue and controlled drug release applications. Chem. Eng. J. 2021, 407. [Google Scholar] [CrossRef]
- Granito, R.N.; Custodio, M.R.; Renno, A.C.M. Natural marine sponges for bone tissue engineering: The state of art and future perspectives. J. Biomed. Mater. Res. Part B Appl. Biomater. 2017, 105, 1717–1727. [Google Scholar] [CrossRef]
- Miricioiu, M.G.; Niculescu, V.C.; Filote, C.; Raboaca, M.S.; Nechifor, G. Coal Fly Ash Derived Silica Nanomaterial for MMMs-Application in CO2/CH4 Separation. Membranes 2021, 11, 78. [Google Scholar] [CrossRef]
- Miricioiu, M.G.; Niculescu, V.C. Fly ash, from recycling to potential raw material for mesoporous silica synthesis. Nanomaterials 2020, 10, 474. [Google Scholar] [CrossRef]
- Jeffryes, C.; Agathos, S.N.; Rorrer, G. Biogenic nanomaterials from photosynthetic microorganisms. Curr. Opin. Biotechnol. 2015, 33, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Castillo, R.R.; Vallet-Regi, M. Functional mesoporous silica nanocomposites: Biomedical applications and biosafety. Int. J. Mol. Sci. 2019, 20, 929. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Principle for Organic Matter Removal | Diatom Species | Diatom Silica Source | Advantages | Drawbacks | Reference(s) | |
|---|---|---|---|---|---|---|---|
| Pre-treatments | HCl | oxidizing washing | Nitzschia closterium, Thalassiosira sp. | freeze-dried samples | high purity of frustules | possible frustule erosion depending on acid strength | [56] |
| KMnO4 + C2H2O4 | oxidizing washing | Fragilariopsis cylindrus, Fragilariopsis kerguelensis, Pseudonitzschia seriata, Thalassiosira nordenskioeldii, Thalassiosira aestivalis, Thalassiosira pseudonana, Thalassiosira weissflogii | wet pellets washed with sodium lauryl sulfate | no frustule erosion | removal of the only external organic matter | [57,58] | |
| Treatments | baking | high temperature | Navicula sp. | APS-fuctionalised diatoms on a mika surface | reduction in hazardous chemicals | possible alterations of pore size, possible post-treatments with acid solutions | [60] |
| low-temperature plasma ashing | ionised gas | Navicula, Amphora, Cocconeis, Planothidium spp. | desalted drops of cultures, freeze-dried samples | no frustule dissolution | unsuitable for saltwater species, expensive, post-treatments with hazardous chemicals | [61,62] | |
| H2O2 | oxidation | DE, Ni tzschia frustulum, Pinnularia and Coscinodiscus spp., Thalassiosira pseudonana, Cylindrotheca closterium | desalted and freeze-dried cultures, diatom composites | less dangerous than strong acids | long incubation, high-temperature post-treatments needful to increase efficiency | [54,62,63,64,65,66,67] | |
| H2SO4 | strong oxidation | Thalassiosira rotula, Coscinodiscus wailesii | living cultures | high efficiency in organic matter removal | hazardous chemicals, dissolution of thin frustules, time-consuming post treatments | [69,70] | |
| H2SO4 + PTFE filters | strong oxidation under vacuum | Nitzschia, Ditylum, Skeletonema, Coscinodiscus | living cultures on a filter cloth | reduced acid amounts | unsuitable for thin frustules | [72] | |
| HNO3 | strong oxidation | Pinnularia sp., Coscinodiscus concinnus | harvested cells | high efficiency in organic matter removal | high-temperature treatments needful to increase efficiency | [68,71] | |
| Piranha solution (H2SO4 + H2O2) | strong oxidation | Thalassiosira pseudonana | PBS-washed cells | high efficiency in organic matter removal | time-consuming post-treatments | [73] |
| Diatom Source | Type of Functionalization | Main Application | Aim | Reference(s) |
|---|---|---|---|---|
| Coscinodiscus wailesii | Silanization and antibody conjugation | Biosensor | Specific recognition antigen–antibody (murine monoclonal antibody) | [88] |
| Coscinodiscus wailesii | Silanization and antibody conjugation | Biosensor | Tethering and detecting antibodies (mix of normal rabbit serum and purified Ig-Y) | [64] |
| Cyclotella sp. | Silanization and antibody conjugation | Biosensor | Selective and label-free photoluminescence-based detection of antigen–antibody (IgG-rabbit) complex formation | [89] |
| Chaetoceros sp. | Iron oxide nanoparticles and antibody conjugation | Biosensor (with magnetic properties) | Selective targeting of SKBR3 cancer cells through the employment of antibody (Trastuzumab) bioconjugation | [87] |
| Thalassiosira weissflogii | Nitroxide 2,6,6-tetramethylpiperidine-N-oxyl (TEMPO) conjugation | Drug carrier | Ciprofloxacin delivery in fibroblasts and osteoblasts | [90] |
| Aulacoseira sp. | Silanization, and oligo (ethylene glycol) methacrylate copolymers addition | Drug carrier | Improvement of levofloxacin delivery | [75] |
| Nitzschia palea | Amino acid (Tyr-ZnII) conjugation | Drug carrier | Inhibition of bacterial growth | [86] |
| Diatomaceous earth | Silanization and phosphonic acids conjugation—self-assembling monolayer | Drug carrier | Improvement of indomethacin and gentamicin delivery | [67] |
| Diatomaceous earth | Silanization and phosphonic acids modifications | Drug carrier | Improvement of indomethacin delivery | [91] |
| DE mineral rocks | Graphene oxide, silanization | Drug carrier | Improvement of indomethacin delivery | [82] |
| Diatomaceous earth | Dopamine modified iron-oxide nanoparticles (DOPA/Fe3O4) | Drug carrier (with magnetic properties) | Improvement of indomethacin delivery | [78] |
| Diatomaceous earth | vitamin B12 and ruthenium (II) complex | Drug carrier | Improvement of the anticancer tris-tetraethyl [2,2′-bipyridine]-4,4′-diamine–ruthenium (II) complex delivery (tested on HT-29 and MCF-7 cancer cells) | [83] |
| Calcined diatomite | Silanization and siRNA conjugation | Drug carrier | Vehiculating siRNA into tumour cells to downregulate the expression of cancer-associated genes (tested on murine A20 lymphoma cells) | [92] |
| Calcined diatomite | Silanization and siRNA conjugation | Drug carrier | Vehiculating siRNA into tumour cells to downregulate the expression of cancer-associated genes (tested on H1355 cancer cells) | [93] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sardo, A.; Orefice, I.; Balzano, S.; Barra, L.; Romano, G. Mini-Review: Potential of Diatom-Derived Silica for Biomedical Applications. Appl. Sci. 2021, 11, 4533. https://doi.org/10.3390/app11104533
Sardo A, Orefice I, Balzano S, Barra L, Romano G. Mini-Review: Potential of Diatom-Derived Silica for Biomedical Applications. Applied Sciences. 2021; 11(10):4533. https://doi.org/10.3390/app11104533
Chicago/Turabian StyleSardo, Angela, Ida Orefice, Sergio Balzano, Lucia Barra, and Giovanna Romano. 2021. "Mini-Review: Potential of Diatom-Derived Silica for Biomedical Applications" Applied Sciences 11, no. 10: 4533. https://doi.org/10.3390/app11104533
APA StyleSardo, A., Orefice, I., Balzano, S., Barra, L., & Romano, G. (2021). Mini-Review: Potential of Diatom-Derived Silica for Biomedical Applications. Applied Sciences, 11(10), 4533. https://doi.org/10.3390/app11104533