Fascial or Muscle Stretching? A Narrative Review

 ,
,  ,
,  ,
, 
 and
and 
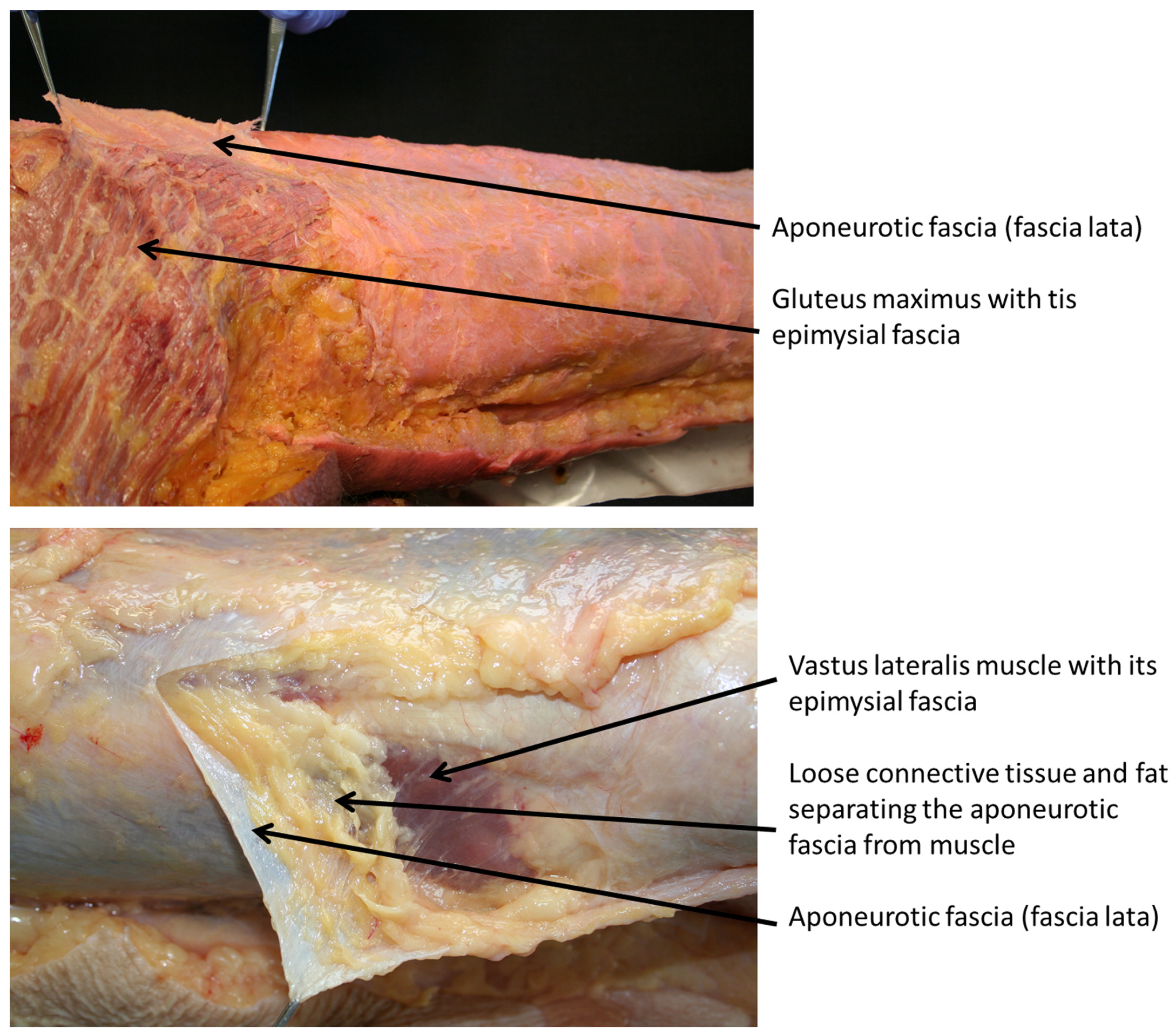{kind=link}
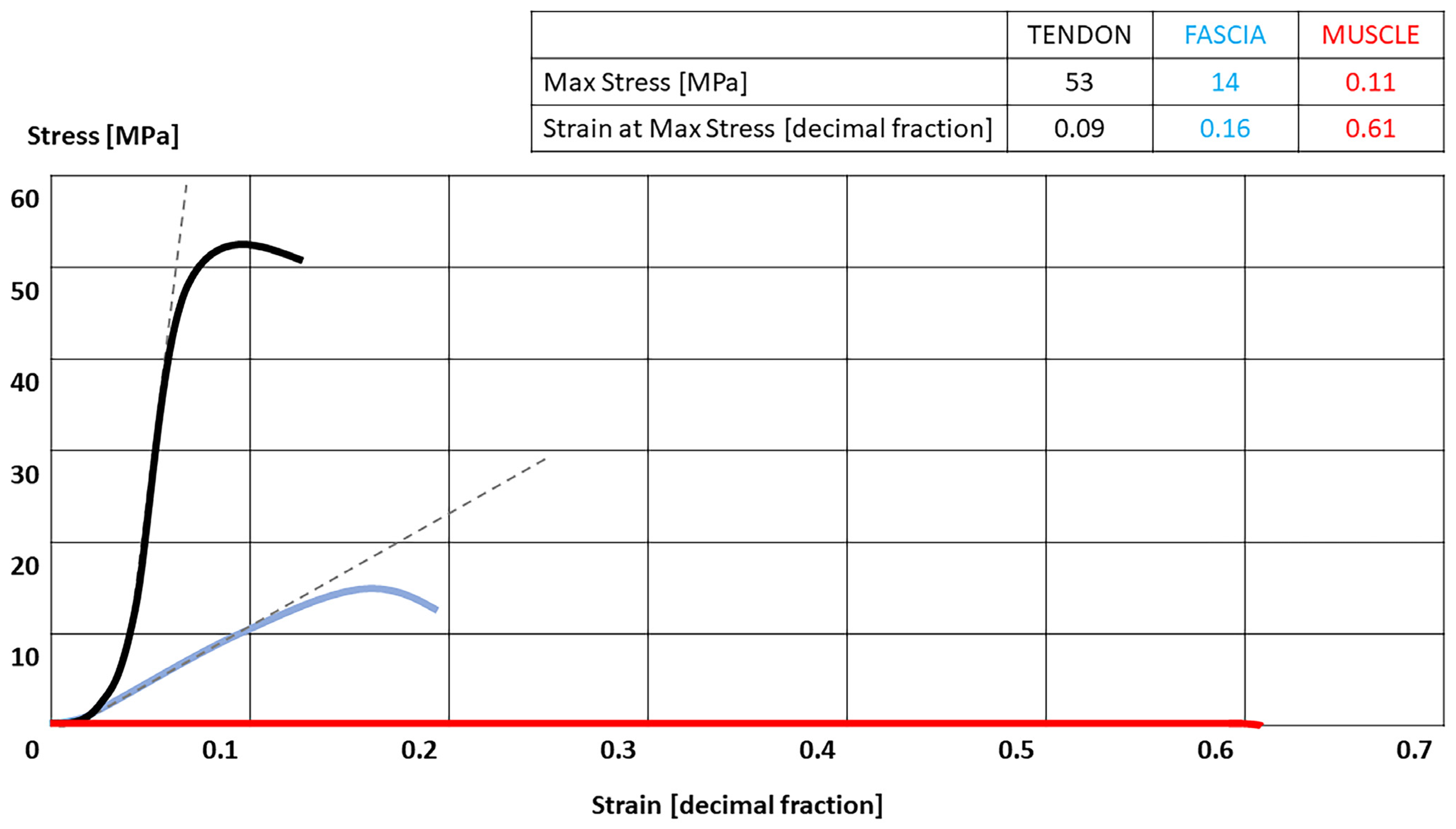{kind=link}
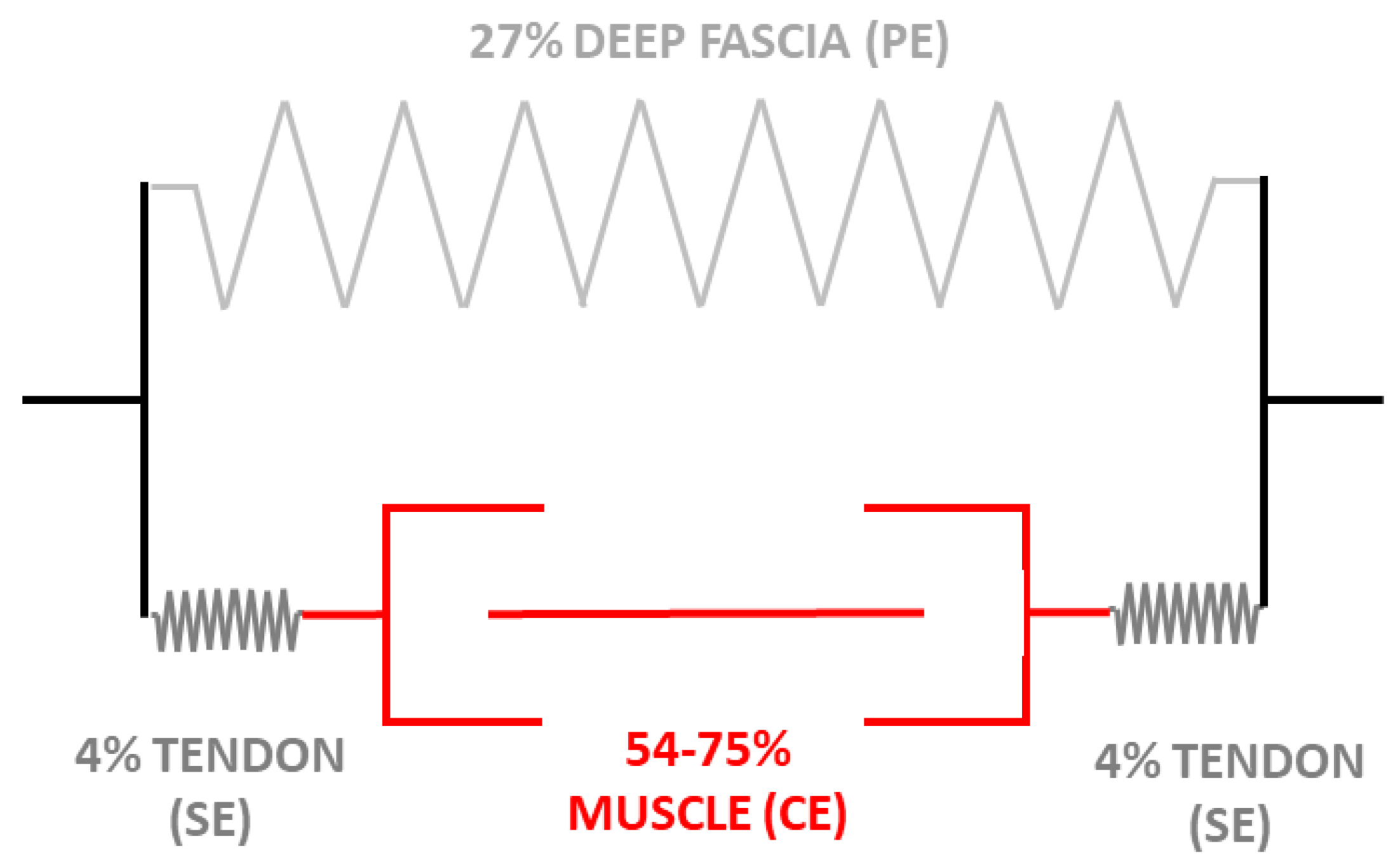{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Stress/Strain Curve
3.2. Stress/Relaxation Curve
3.3. Thixotropic Effect
4. Discussion
Limits
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cornelius, W.L.; Ebrahim, K.; Watson, J.; Hill, D.W. The Effects of Cold Application and Modified PNF Stretching Techniques on Hip Joint Flexibility in College Males. Res. Q. Exerc. Sport 1992, 63, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Konrad, A.; Tilp, M. Increased range of motion after static stretching is not due to changes in muscle and tendon structures. Clin. Biomech. 2014, 29, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Ikezoe, T.; Nishishita, S.; Umehara, J.; Kimura, M.; Ichihashi, N. Acute effects of static stretching on the shear elastic moduli of the medial and lateral gastrocnemius muscles in young and elderly women. Musculoskelet. Sci. Pract. 2017, 32, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, S.P.; Simonsen, E.B.; Aagaard, P.; Gleim, G.W.; McHugh, M.P.; Kjaer, M. Viscoelastic response to repeated static stretching in human skeletal muscle. Scand. J. Med. Sci. Sport 1995, 5, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Osternig, L.R.; Robertson, R.N.; Troxel, R.K.; Hansen, P. Differential responses to proprioceptive neuromuscular facilitation (PNF) stretch techniques. Med. Sci. Sports Exerc. 1990, 22, 106–111. [Google Scholar] [CrossRef]
- Magnusson, S.P.; Simonsen, E.B.; Dyhre-Poulsen, P.; Aagaard, P.; Mohr, T.; Kjaer, M. Viscoelastic stress relaxation during static stretch in human skeletal muscle in the absence of EMG activity. Scand. J. Med. Sci. Sports 1996, 6, 323–328. [Google Scholar] [CrossRef]
- Chaouachi, A.; Padulo, J.; Kasmi, S.; Ben Othmen, A.; Chatra, M.; Behm, D.G. Unilateral static and dynamic hamstrings stretching increases contralateral hip flexion range of motion. Clin. Physiol. Funct. Imaging 2015, 37, 23–29. [Google Scholar] [CrossRef]
- Behm, D.G.; Cavanaugh, T.; Quigley, P.; Reid, J.C.; Nardi, P.S.M.; Marchetti, P.H. Acute bouts of upper and lower body static and dynamic stretching increase non-local joint range of motion. Graefe’s Arch. Clin. Exp. Ophthalmol. 2016, 116, 241–249. [Google Scholar] [CrossRef]
- Wilke, J.; Niederer, D.; Vogt, L.; Banzer, W. Remote effects of lower limb stretching: Preliminary evidence for myofascial connectivity? J. Sports Sci. 2016, 34, 1–4. [Google Scholar] [CrossRef]
- Cruz-Montecinos, C.; González Blanche, A.; López Sánchez, D.; Cerda, M.; Sanzana-Cuche, R.; Cuesta-Vargas, A. In vivo relationship between pelvis motion and deep fascia displacement of the medial gastrocnemius: Anatomical and functional implications. J. Anat. 2015, 227, 665–672. [Google Scholar] [CrossRef]
- Andrade, R.J.; Freitas, S.R.; Vaz, J.R.; Bruno, P.M.; Pezarat-Correia, P. Provocative mechanical tests of the peripheral nervous system affect the joint torque-angle during passive knee motion. Scand. J. Med. Sci. Sports 2014, 25, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Pavan, P.G.; Stecco, A.; Stern, R.; Stecco, C. Painful Connections: Densification Versus Fibrosis of Fascia. Curr. Pain Headache Rep. 2014, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cowman, M.K.; Schmidt, T.A.; Raghavan, P.; Stecco, A. Viscoelastic Properties of Hyaluronan in Physiological Conditions. F1000Research 2015, 4, 622. [Google Scholar] [CrossRef] [PubMed]
- Fede, C.; Angelini, A.; Stern, R.; Macchi, V.; Porzionato, A.; Ruggieri, P.; De Caro, R.; Stecco, C. Quantification of hyaluronan in human fasciae: Variations with function and anatomical site. J. Anat. 2018, 233, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Stecco, C.; Hammer, W.; Vleeming, A.; De Caro, R. Functional Atlas of the Human Fascial System; Elsevier Health Sciences: London, UK, 2015. [Google Scholar]
- Yucesoy, C.A.; Baan, G.C.; Huijing, P.A. Substantial inter-antagonistic epimuscular myofascial force transmission occurs in the rat between the deep flexor muscles and the muscles of the anterior crural and peroneal compartments. J. Electromyogr. Kinesiol. 2010, 20, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Stecco, C.; Stern, R.; Porzionato, A.; Macchi, V.; Masiero, S.; Stecco, A.; De Caro, R. Hyaluronan within fascia in the etiology of myofascial pain. Surg. Radiol. Anat. 2011, 33, 891–896. [Google Scholar] [CrossRef]
- Krause, F.; Wilke, J.; Vogt, L.; Banzer, W. Intermuscular force transmission along myofascial chains: A systematic review. J. Anat. 2016, 228, 910–918. [Google Scholar] [CrossRef]
- Huijing, P.A. Muscle as a collagen fiber reinforced composite material: Force transmission in muscle and whole limbs. J. Biomech. 1999, 32, 329–345. [Google Scholar] [CrossRef]
- Huijing, P.A. Muscular force transmission: A unified, dual or multiple sytem? A review and some explorative experimental results. Arch. Physiol. Biochem. 1999, 170, 292–311. [Google Scholar]
- Huijing, P.; Baan, G. Myofascial Force Transmission Causes Interaction between Adjacent Muscles and Connective Tissue: Effects of Blunt Dissection and Compartmental Fasciotomy on Length Force Characteristics of Rat Extensor Digitorum Longus Muscle. Arch. Physiol. Biochem. 2001, 109, 97–109. [Google Scholar] [CrossRef]
- Maas, H.; Meijer, H.J.M.; Huijing, P.A. Intermuscular Interaction between Synergists in Rat Originates from both Intermuscular and Extramuscular Myofascial Force Transmission. Cells Tissues Organs 2005, 181, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Yucesoy, C.A.; Koopman, H.J.F.M.; Baan, G.C.; Grootenboer, H.J.; Huijing, P.A. Extramuscular myofascial force transmis-sion: Experiments and finite element modeling. Arch. Physiol. Biochem. 2003, 111, 377–388. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yucesoy, C.A.; Baan, G.C.; Koopman, B.H.F.J.M.; Grootenboer, H.J.; Huijing, P.A. Pre-Strained Epimuscular Connections Cause Muscular Myofascial Force Transmission to Affect Properties of Synergistic EHL and EDL Muscles of the Rat. J. Biomech. Eng. 2005, 127, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.S.; Bilgili, F.; Akalan, N.E.; Temelli, Y.; Ates, F.; Yucesoy, C.A. Intraoperative experiments combined with gait analyses indicate that active state rather than passive dominates the spastic gracilis muscle’s joint movement limiting effect in cerebral palsy. Clin. Biomech. 2019, 68, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Stecco, A.; Gilliar, W.; Hill, R.; Fullerton, B.; Stecco, C. The anatomical and functional relation between gluteus maximus and fascia lata. J. Bodyw. Mov. Ther. 2013, 17, 512–517. [Google Scholar]
- Stecco, C.; Gagey, O.; Macchi, V.; Porzionato, A.; De Caro, R.; Aldegheri, R.; Delmas, V. Tendinous muscular insertions onto the deep fascia of the upper limb. First part: Anatomical study. Morphology 2007, 91, 29–37. [Google Scholar] [CrossRef]
- Berthier, C.; Blaineau, S. Supramolecular organization of the subsarcolemmal cytoskeleton of adult skeletal muscle fibers. A review. Biol. Cell 1997, 89, 413–434. [Google Scholar] [CrossRef]
- Nishimura, T.; Ojima, K.; Liu, A.; Hattori, A.; Takahashi, K. Structural changes in the intramuscular connective tissue during development of bovine semitendinosus muscle. Tissue Cell 1996, 28, 527–536. [Google Scholar] [CrossRef]
- Passerieux, E.; Rossignol, R.; Letellier, T.; Delage, J.P. Physical continuity of the perimysium from myofibers to tendons: In-volvement in lateral force transmission in skeletal muscle. J. Struct. Biol. 2007, 159, 19–28. [Google Scholar] [CrossRef]
- Street, S.F.; Ramsey, R.W. Sarcolemma: Transmitter of Active Tension in Frog Skeletal Muscle. Science 1965, 149, 1379–1380. [Google Scholar] [CrossRef]
- Stecco, L. Il Dolore e le Sequenze Neuromiofasciali; IPSA Editore: Palermo, Italy, 1990. [Google Scholar]
- Nordez, A.; Gross, R.; Andrade, R.; Le Sant, G.; Freitas, S.; Ellis, R.; McNair, P.J.; Hug, F. Non-Muscular Structures Can Limit the Maximal Joint Range of Motion during Stretching. Sports Med. 2017, 47, 1925–1929. [Google Scholar] [CrossRef] [PubMed]
- Behm, D.G. The Science and Physiology of Flexibility and Stretching; Routledge: Abingdon, UK, 2018. [Google Scholar]
- Korhonen Rami, K.; Saarakkala, S. Biomechanics and modeling of skeletal soft tissues. In Theoretical Bio-Mechanics; Klika, V., Ed.; IntechOpen: London, UK, 2011. [Google Scholar]
- Pavan, P.G.; Stecco, C.; Darwish, S.; Natali, A.N.; De Caro, R. Investigation of the mechanical properties of the plantar apo-neurosis. Surg. Radiol. Anat. 2011, 33, 905–911. [Google Scholar] [CrossRef] [PubMed]
- Stecco, C.; Pavan, P.; Pachera, P.; De Caro, R.; Natali, A. Investigation of the mechanical properties of the human crural fascia and their possible clinical implications. Surg. Radiol. Anat. 2013, 36, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Pavan, P.G.; Pachera, P.; Stecco, C.; Natali, A.N. Biomechanical behavior of human crural fascia in anterior and posterior regions of the lower limb. Med. Biol. Eng. Comput. 2015, 53, 951–959. [Google Scholar] [CrossRef]
- Bilston, L.E.; Tan, K. Measurement of Passive Skeletal Muscle Mechanical Properties In Vivo: Recent Progress, Clinical Applications, and Remaining Challenges. Ann. Biomed. Eng. 2015, 43, 261–273. [Google Scholar] [CrossRef]
- Meyers, S.A.; Seaber, A.V.; Glisson, R.R.; Nunley, J.A. Effect of hyaluronic acid/chondroitin sulfate on healing of full-thickness tendon lacerations in rabbits. J. Orthop. Res. 1989, 7, 683–689. [Google Scholar] [CrossRef]
- Tamura, A.; Hongu, J.-I.; Matsumoto, T. Theoretical elastic tensile behavior of muscle fiber bundles in traumatic loading events. Clin. Biomech. 2019, 69, 184–190. [Google Scholar] [CrossRef]
- Schleifenbaum, S.; Schmidt, M.; Möbius, R.; Wolfskämpf, T.; Schröder, C.; Grunert, R.; Hammer, N.; Prietzel, T. Load and failure behavior of human muscle samples in the context of proximal femur replacement. BMC Musculoskelet. Disord. 2016, 17, 149. [Google Scholar] [CrossRef]
- Tamura, A.; Hayashi, S.; Matsumoto, T. Effect of Loading Rate on Viscoelastic Properties and Local Mechanical Heterogeneity of Freshly Isolated Muscle Fiber Bundles Subjected tTo Uniaxial Stretching. J. Mech. Med. Biol. 2016, 16, 1650086. [Google Scholar] [CrossRef]
- Malanga, G.A.; Ibrahim, V. Regenerative Treatments in Sports and Orthopedic Medicine; Demosmedical: New York, NY, USA, 2018. [Google Scholar]
- Morales-Orcajo, E.; Bayod, J.; Casas, E.B.D.L. Computational Foot Modeling: Scope and Applications. Arch. Comput. Methods Eng. 2015, 23, 389–416. [Google Scholar] [CrossRef]
- Soderberg, G.L. Kinesiology: Application to Pathological Motion; Williams & Wilkins: Baltimore, MD, USA, 1986. [Google Scholar]
- Bennett, M.B.; Ker, R.F.; Imery, N.J.; Alexander, R.M. Mechanical properties of various mammalian tendons. J. Zool. 1986, 209, 537–548. [Google Scholar] [CrossRef]
- Ker, R.F. Dynamic tensile properties of the plantaris tendon of sheep (Ovis aries). J. Exp. Biol. 1981, 93, 283–302. [Google Scholar] [PubMed]
- Kubo, K.; Kanehisa, H.; Fukunaga, T. Effect of stretching training on the viscoelastic properties of human tendon structures in vivo. J. Appl. Physiol. 2002, 92, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Duenwald, S.E.; Vanderby, R., Jr.; Lakes, R.S. Stress relaxation and recovery in tendon and ligament: Experiment and modeling. Biorheology 2010, 47, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Proske, U. Exercise, fatigue and proprioception: A retrospective. Exp. Brain Res. 2019, 237, 2447–2459. [Google Scholar] [CrossRef]
- Smith, L.K.; Weiss, E.L.; Lehmkuhl, L.D. Brunnstrom’s Clinical Kinesiology, 5th ed.; F.A. Davis: Philadelphia, PA, USA, 1996; pp. 138–142. [Google Scholar]
- Kjaer, M. Role of Extracellular Matrix in Adaptation of Tendon and Skeletal Muscle to Mechanical Loading. Physiol. Rev. 2004, 84, 649–698. [Google Scholar] [CrossRef]
- Dintenfass, L. Lubrication in Synovial Joints: A Theoretical Analysis. J. Bone Jt. Surg. Am. 1963, 45, 1241–1256. [Google Scholar] [CrossRef]
- Chytil, M.; Strand, S.; Christensen, B.E.; Pekař, M. Calorimetric and light scattering study of interactions and macromolecular properties of native and hydrophobically modified hyaluronan. Carbohydr. Polym. 2010, 81, 855–863. [Google Scholar] [CrossRef]
- Tadmor, R.; Chen, N.; Israelachvili, J.N. Thin film rheology and lubricity of hyaluronic acid solutions at a normal physiological concentration. J. Biomed. Mater. Res. 2002, 61, 514–523. [Google Scholar] [CrossRef]
- Matteini, P.; Dei, L.; Carretti, E.; Volpi, N.; Goti, A.; Pini, R. Structural Behavior of Highly Concentrated Hyaluronan. Biomacromolecules 2009, 10, 1516–1522. [Google Scholar] [CrossRef]
- Tømmeraas, K.; Melander, C. Kinetics of Hyaluronan Hydrolysis in Acidic Solution at Various pH Values. Biomacromolecules 2008, 9, 1535–1540. [Google Scholar] [CrossRef] [PubMed]
- Gatej, I.; Popa, M.; Rinaudo, M. Role of the pH on Hyaluronan Behavior in Aqueous Solution. Biomacromolecules 2005, 6, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Juel, C.; Klarskov, C.; Nielsen, J.J.; Krustrup, P.; Mohr, M.; Bangsbo, J. Effect of high-intensity intermittent training on lactate and H+ release from human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E245–E251. [Google Scholar] [CrossRef] [PubMed]
- Karakuzu, A.; Pamuk, U.; Ozturk, C.; Acar, B.; Yucesoy, C.A. Magnetic resonance and diffusion tensor imaging analyses indicate heterogeneous strains along human medial gastrocnemius fascicles caused by submaximal plantar-flexion activity. J. Biomech. 2017, 57, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Perret, C.; Poiraudeau, S.; Fermanian, J.; Colau, M.M.L.; Benhamou, M.A.M.; Revel, M. Validity, reliability, and responsiveness of the fingertip-to-floor test. Arch. Phys. Med. Rehabil. 2001, 82, 1566–1570. [Google Scholar] [CrossRef] [PubMed]
- Konrad, A.; Stafilidis, S.; Tilp, M. Effects of acute static, ballistic, and PNF stretching exercise on the muscle and tendon tissue properties. Scand. J. Med. Sci. Sports 2016, 27, 1070–1080. [Google Scholar] [CrossRef]
- Roser, M.; Cameron, A.; Hannah, R. “Human Height”. Available online: OurWorldInData.org (accessed on 7 October 2020).
- Herda, T.J.; Cramer, J.T.; Ryan, E.D.; McHugh, M.P.; Stout, J. Acute Effects of Static versus Dynamic Stretching on Isometric Peak Torque, Electromyography, and Mechanomyography of the Biceps Femoris Muscle. J. Strength Cond. Res. 2008, 22, 809–817. [Google Scholar] [CrossRef]
- Nordez, A.; McNair, P.; Casari, P.; Cornu, C. The effect of angular velocity and cycle on the dissipative properties of the knee during passive cyclic stretching: A matter of viscosity or solid friction. Clin. Biomech. 2009, 24, 77–81. [Google Scholar] [CrossRef]
- Bishop, D.J. Warm up I: Potential mechanisms and the effects of passive warm up on exercise performance. Sports Med. 2003, 33, 439–454. [Google Scholar] [CrossRef]
- Buchthal, F.; Kaiser, E.; Knappeis, G.G. Elasticity, Viscosity and Plasticity in the Cross Striated Muscle Fibre. Acta Physiol. Scand. 2008, 8, 16–37. [Google Scholar] [CrossRef]
- Hill, A.V. The heat of shortening and the dynamic constants of muscle. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2006, 126, 136–195. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stecco, C.; Pirri, C.; Fede, C.; Yucesoy, C.A.; De Caro, R.; Stecco, A. Fascial or Muscle Stretching? A Narrative Review. Appl. Sci. 2021, 11, 307. https://doi.org/10.3390/app11010307
Stecco C, Pirri C, Fede C, Yucesoy CA, De Caro R, Stecco A. Fascial or Muscle Stretching? A Narrative Review. Applied Sciences. 2021; 11(1):307. https://doi.org/10.3390/app11010307
Chicago/Turabian StyleStecco, Carla, Carmelo Pirri, Caterina Fede, Can A. Yucesoy, Raffaele De Caro, and Antonio Stecco. 2021. "Fascial or Muscle Stretching? A Narrative Review" Applied Sciences 11, no. 1: 307. https://doi.org/10.3390/app11010307
APA StyleStecco, C., Pirri, C., Fede, C., Yucesoy, C. A., De Caro, R., & Stecco, A. (2021). Fascial or Muscle Stretching? A Narrative Review. Applied Sciences, 11(1), 307. https://doi.org/10.3390/app11010307