Modelling of Red Blood Cell Morphological and Deformability Changes during In-Vitro Storage




Abstract
Featured Application
Abstract
1. Introduction
2. Methods
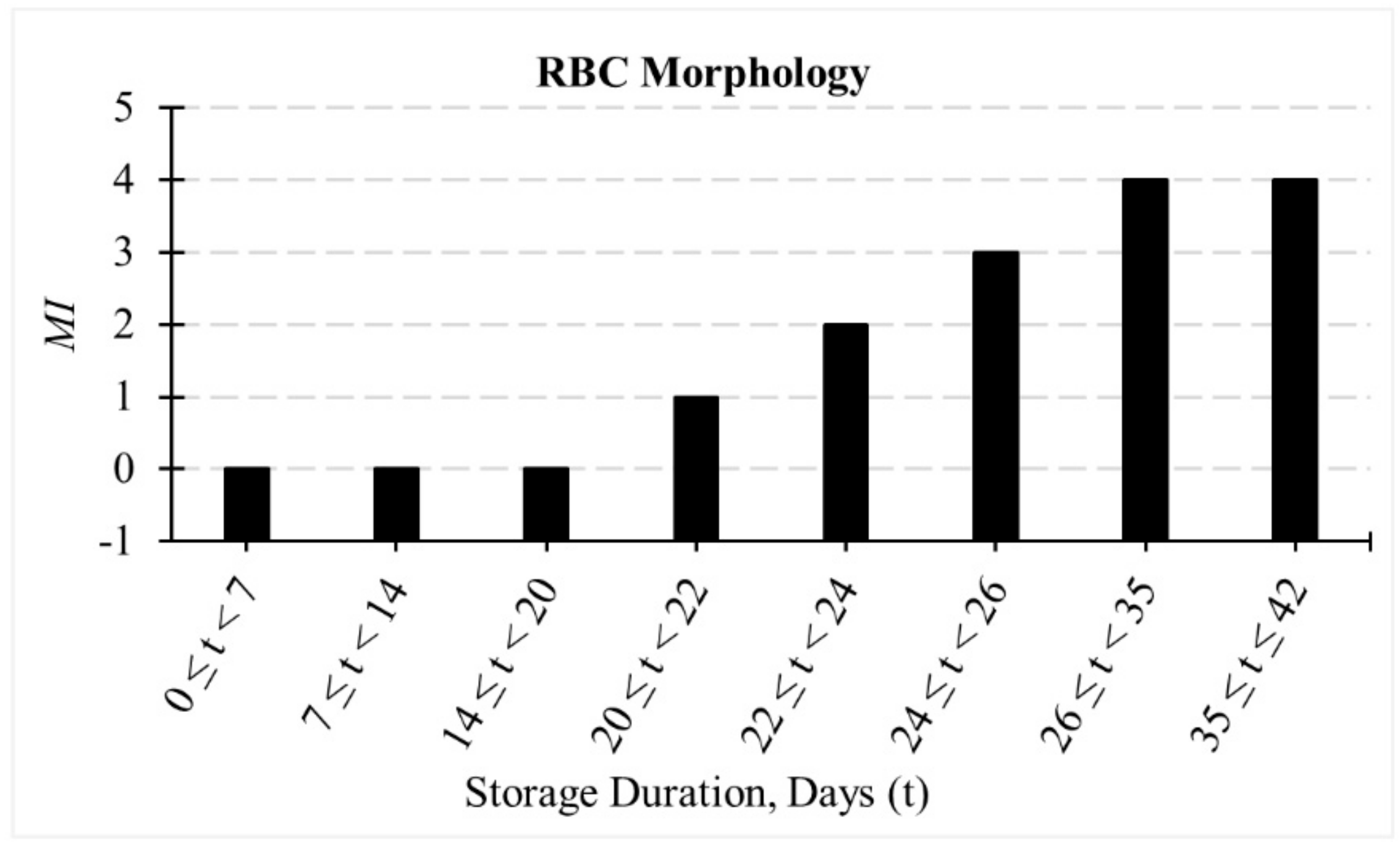
2.1. Evolution of RBC Morphology during In-Vitro Storage
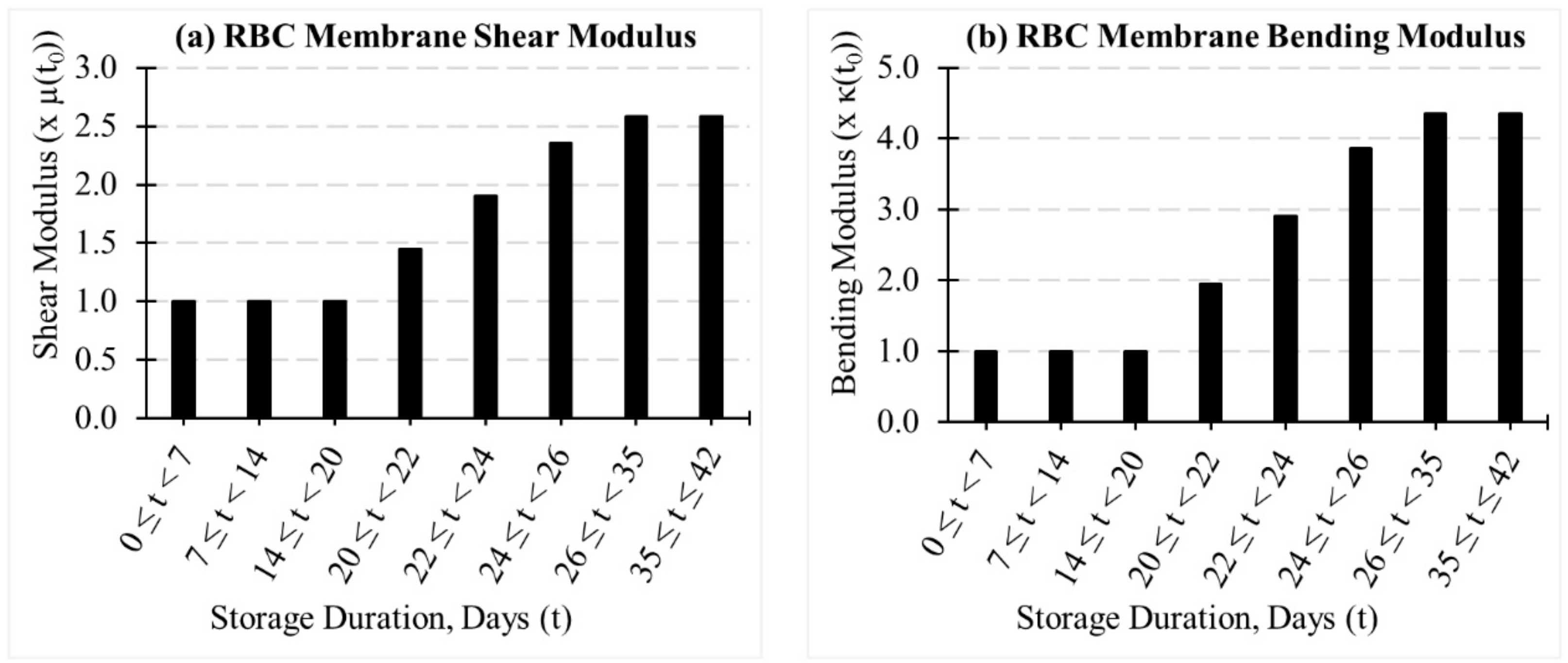
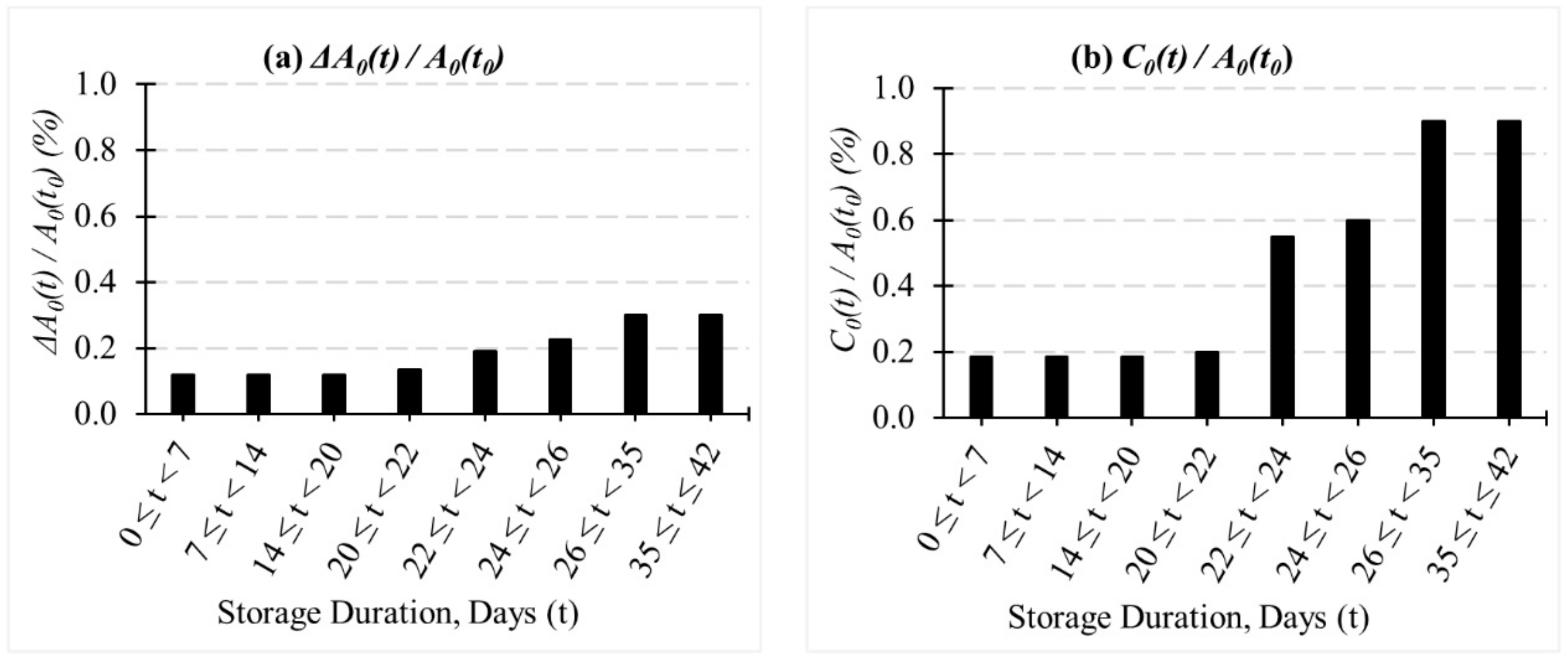
2.2. Evolution of Coarse-Grained (CG)-RBC Membrane Model Parameters during In-Vitro Storage
2.3. CG-RBC Membrane Model-Predicted RBC Morphologies during In-Vitro Storage
3. Results and Discussion
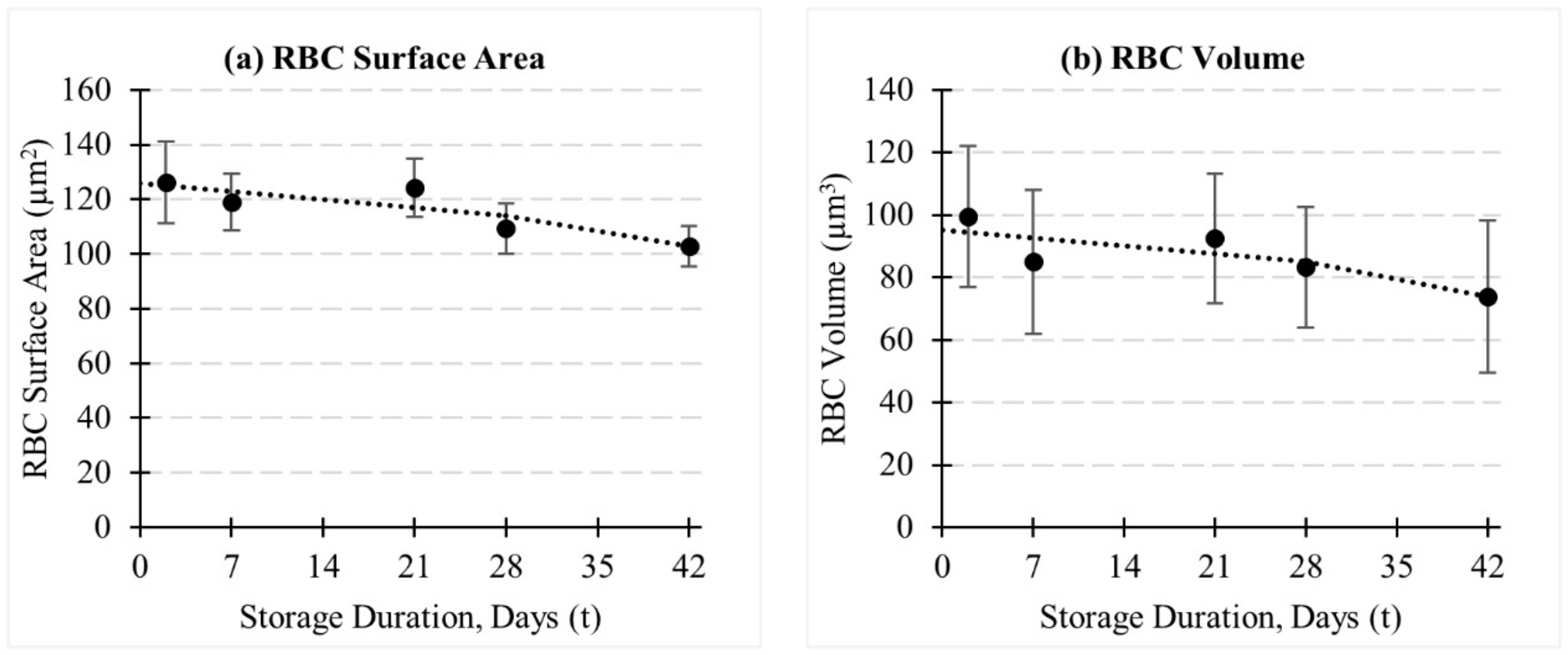
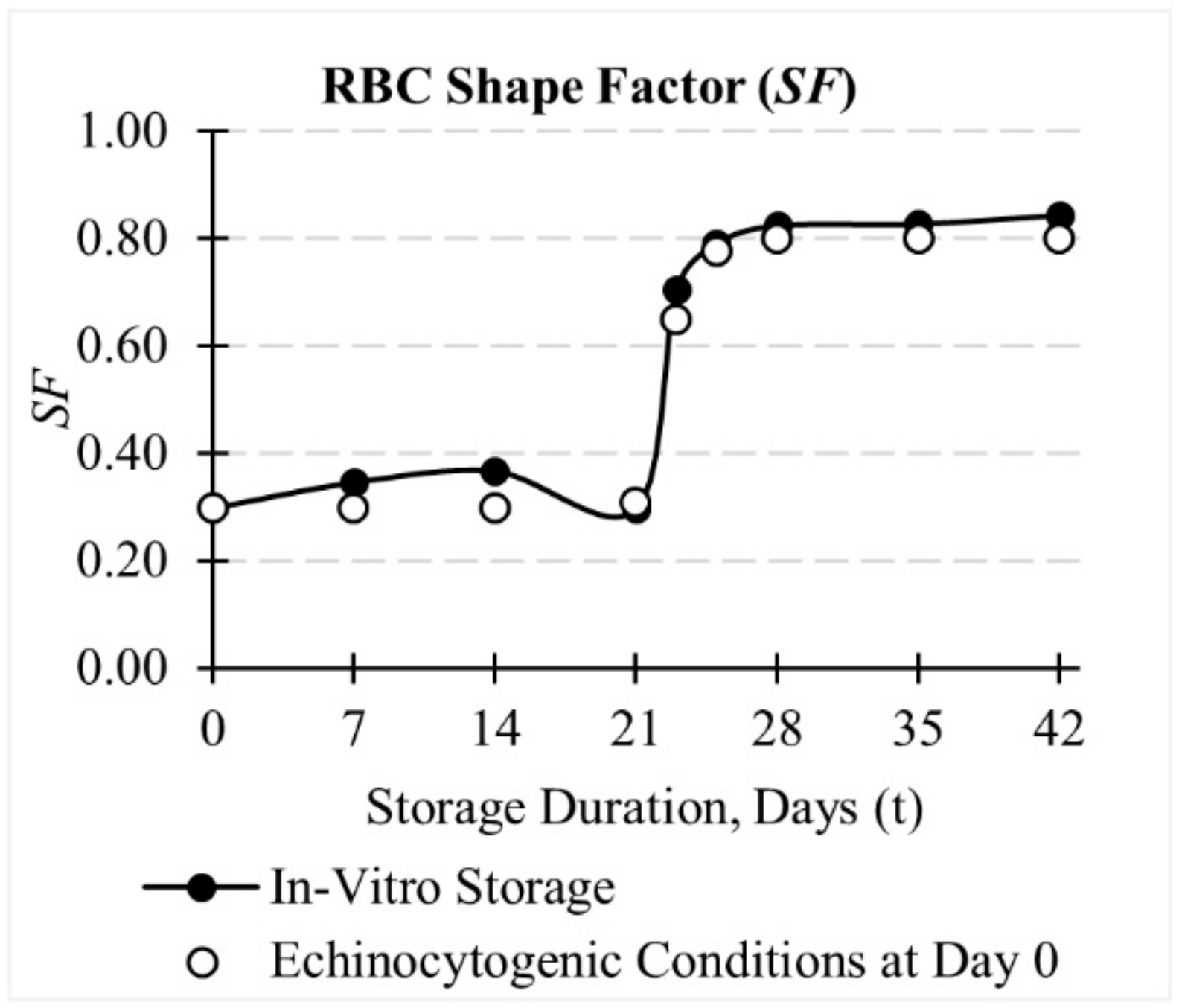
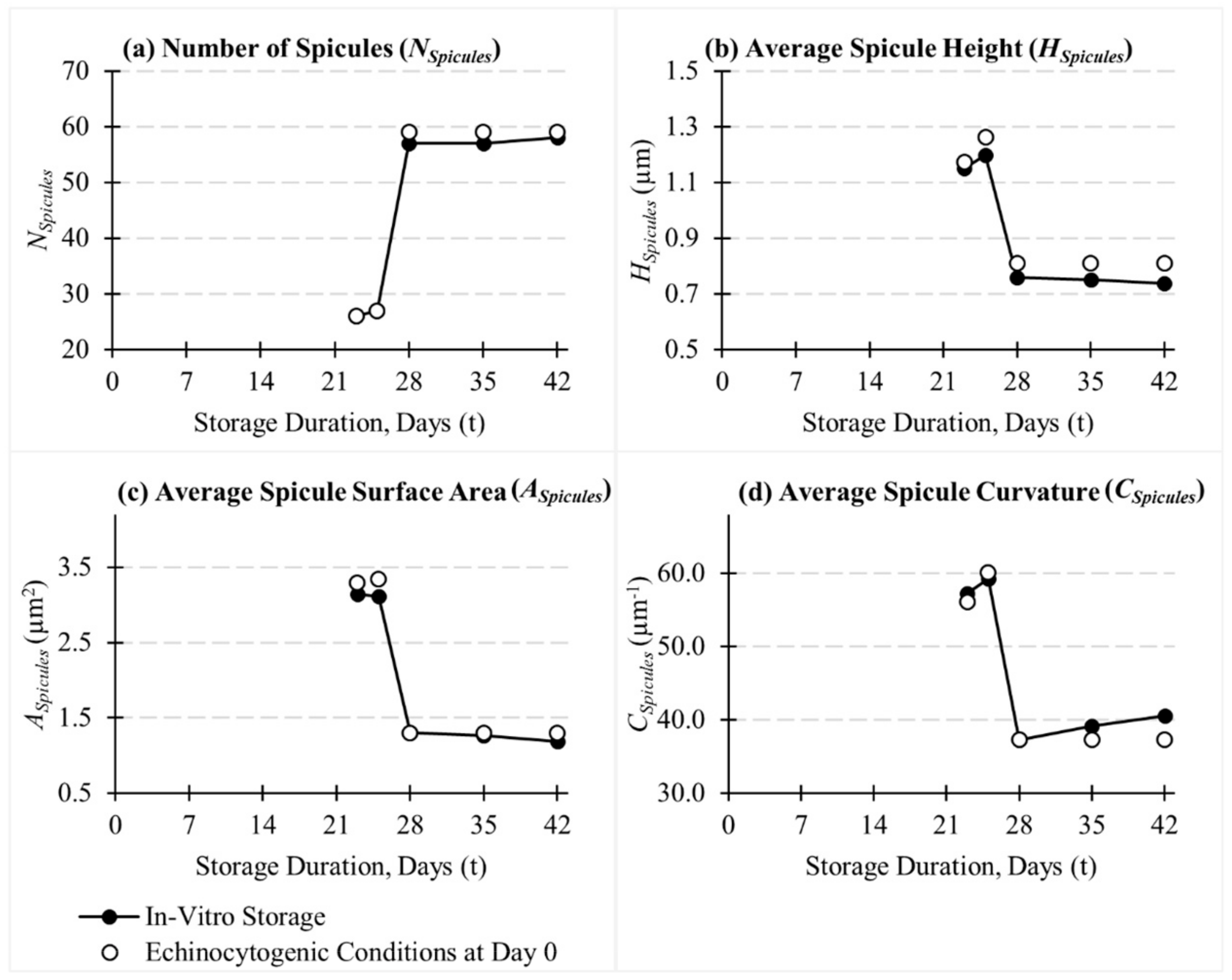
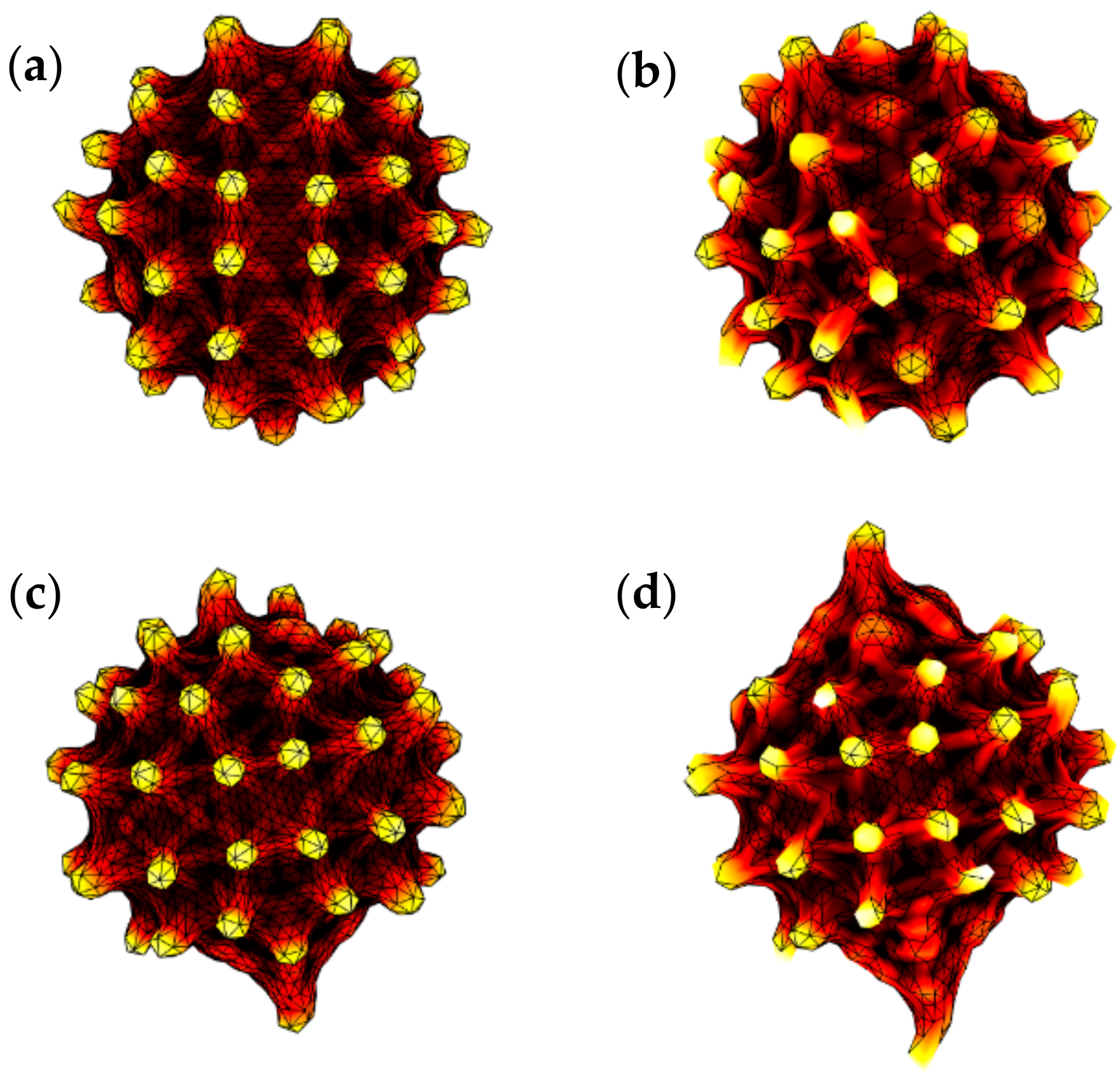
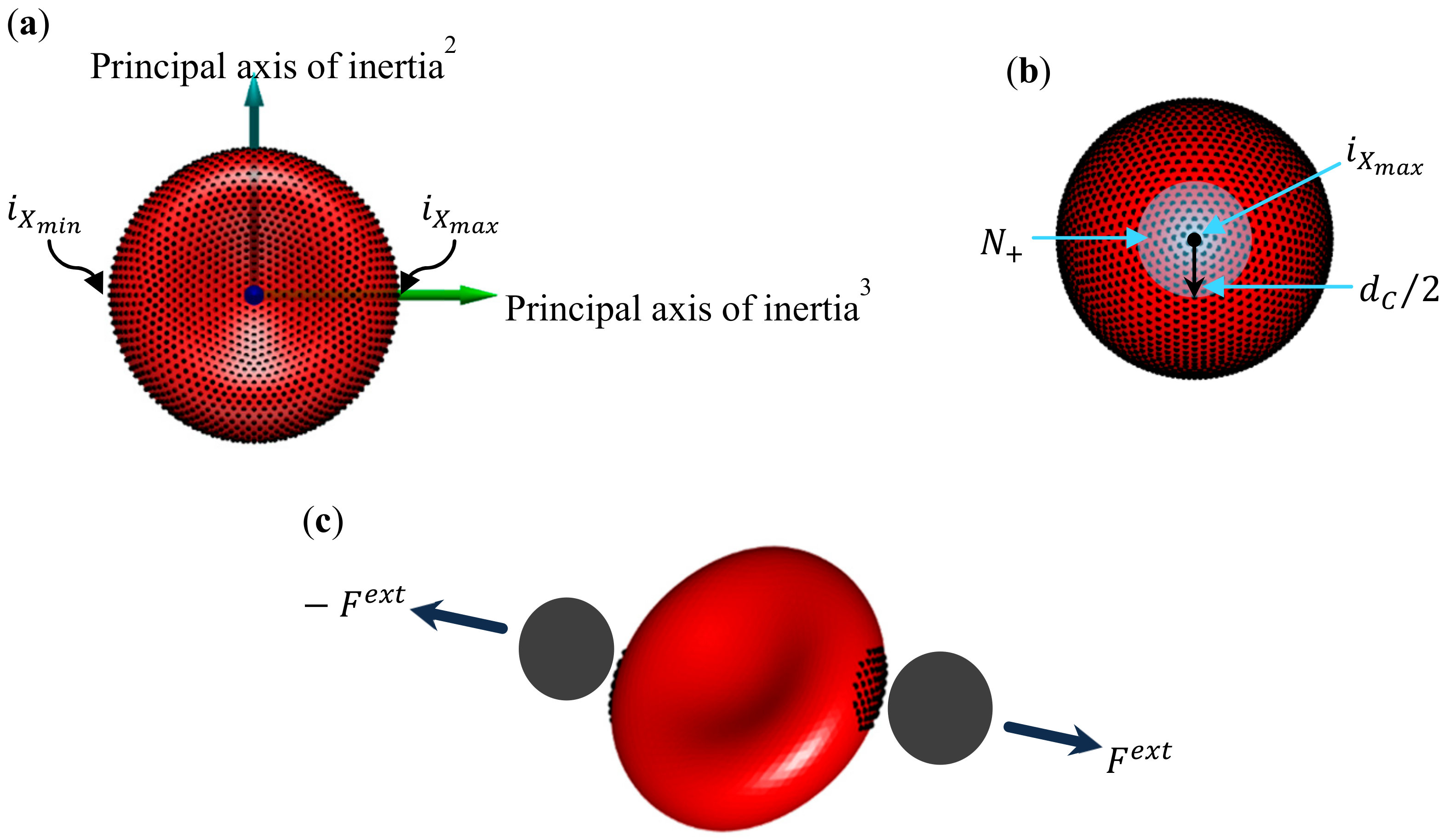
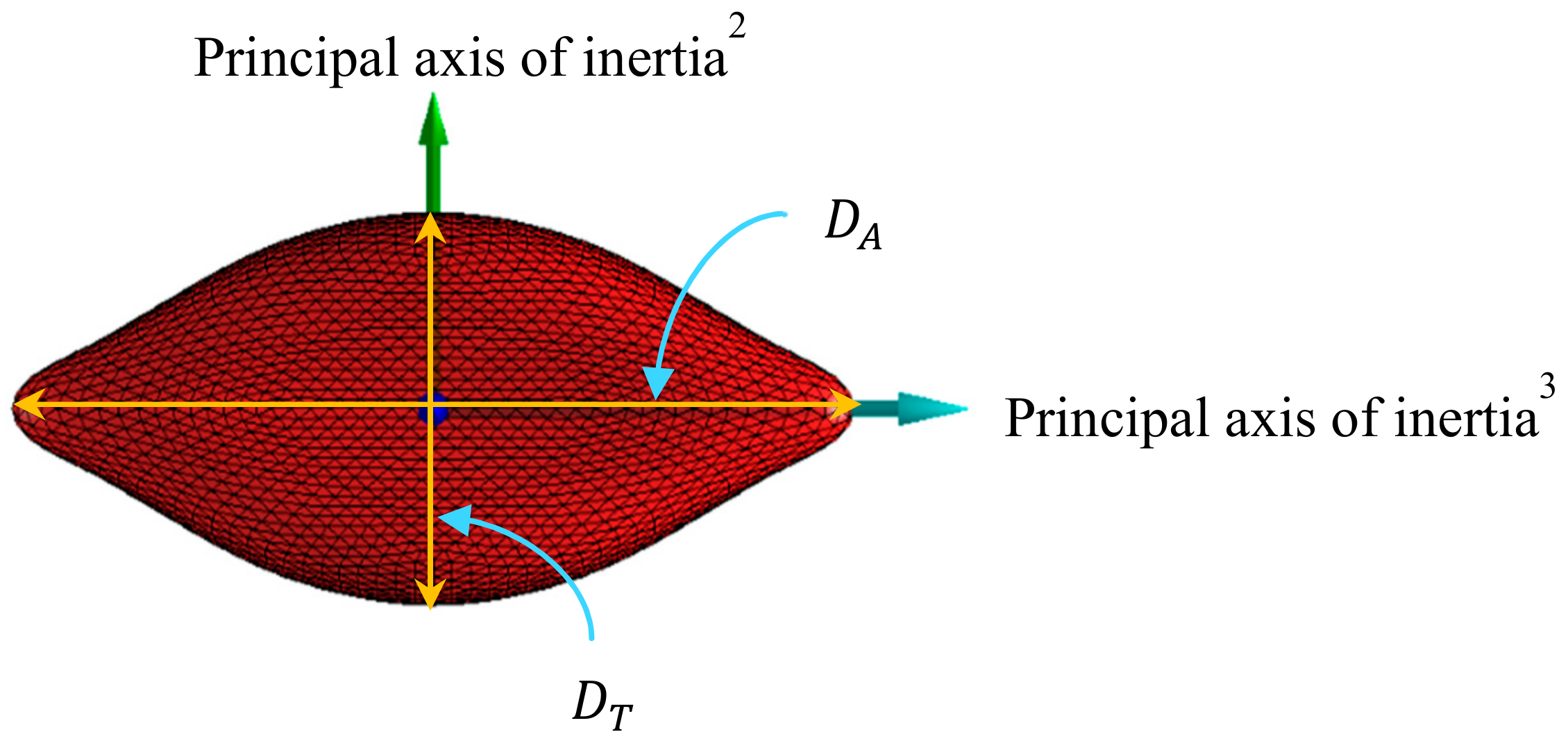
3.1. Evolution of RBC Morphology Characteristics during In-Vitro Storage
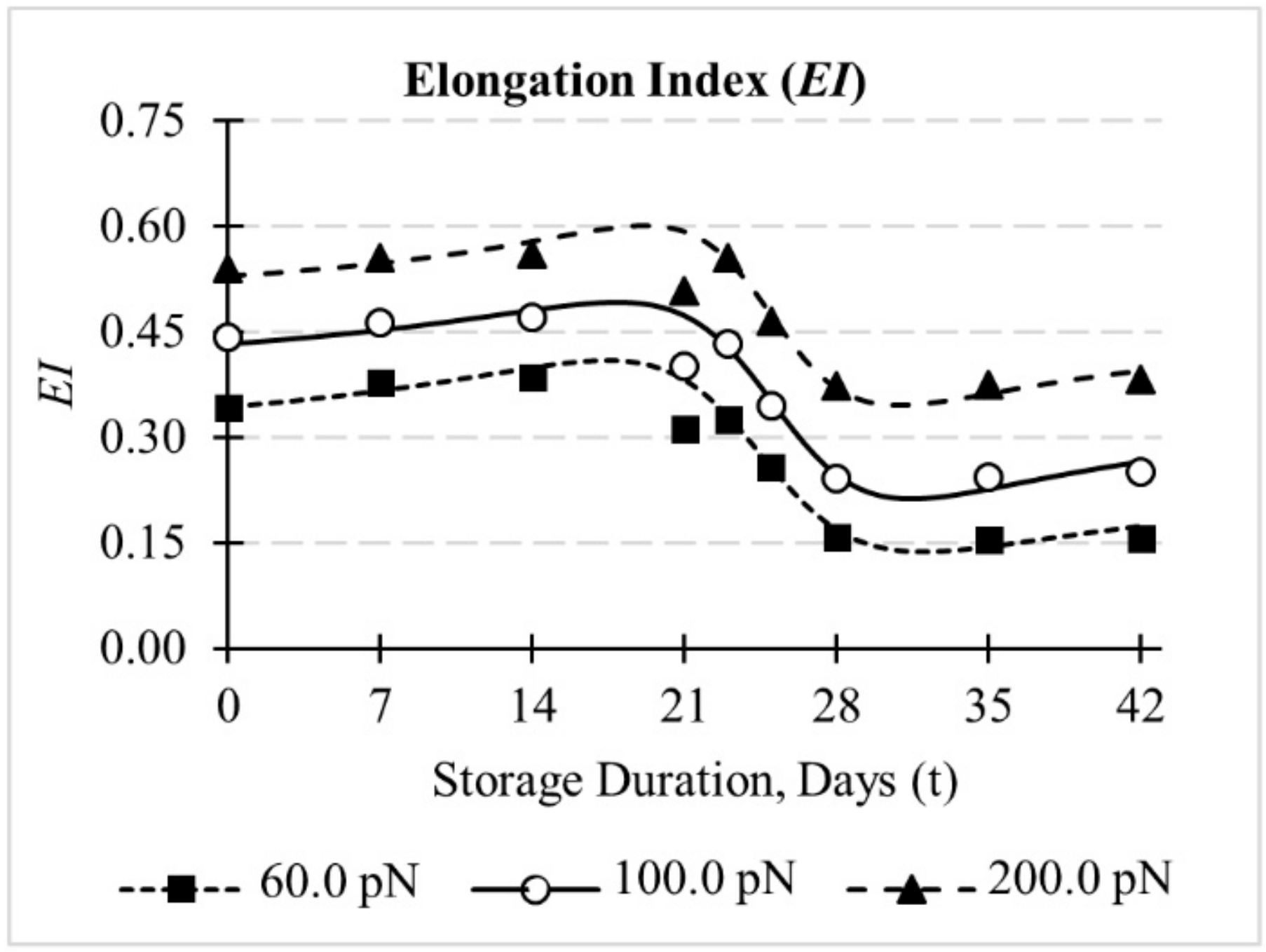
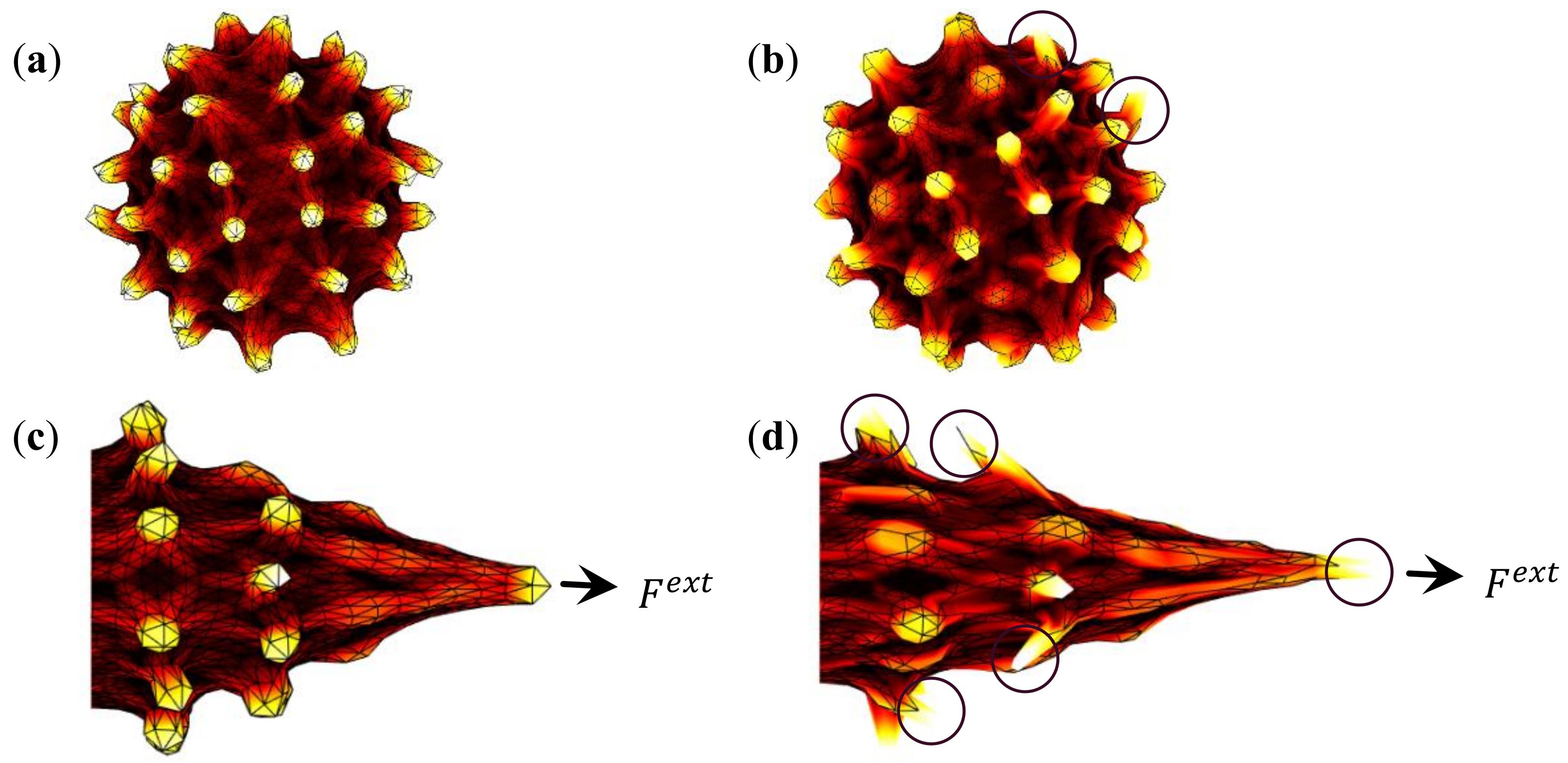
3.2. Evolution of Deformation Behaviour of RBCs during In-Vitro Storage
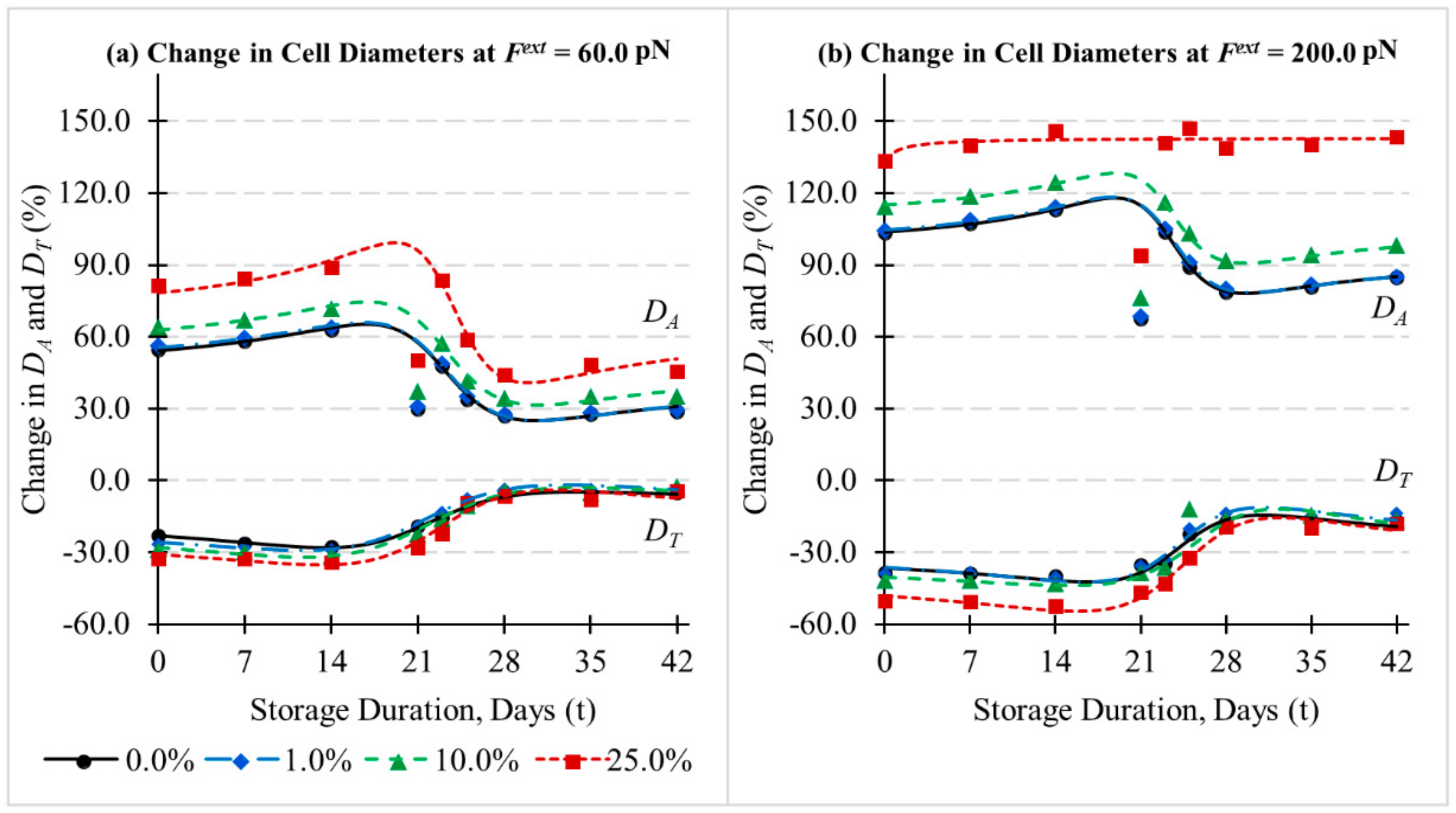
3.3. Influence of Cytoskeletal Actin Junctional Defects during In-Vitro Storage on RBC Morphology and Deformability
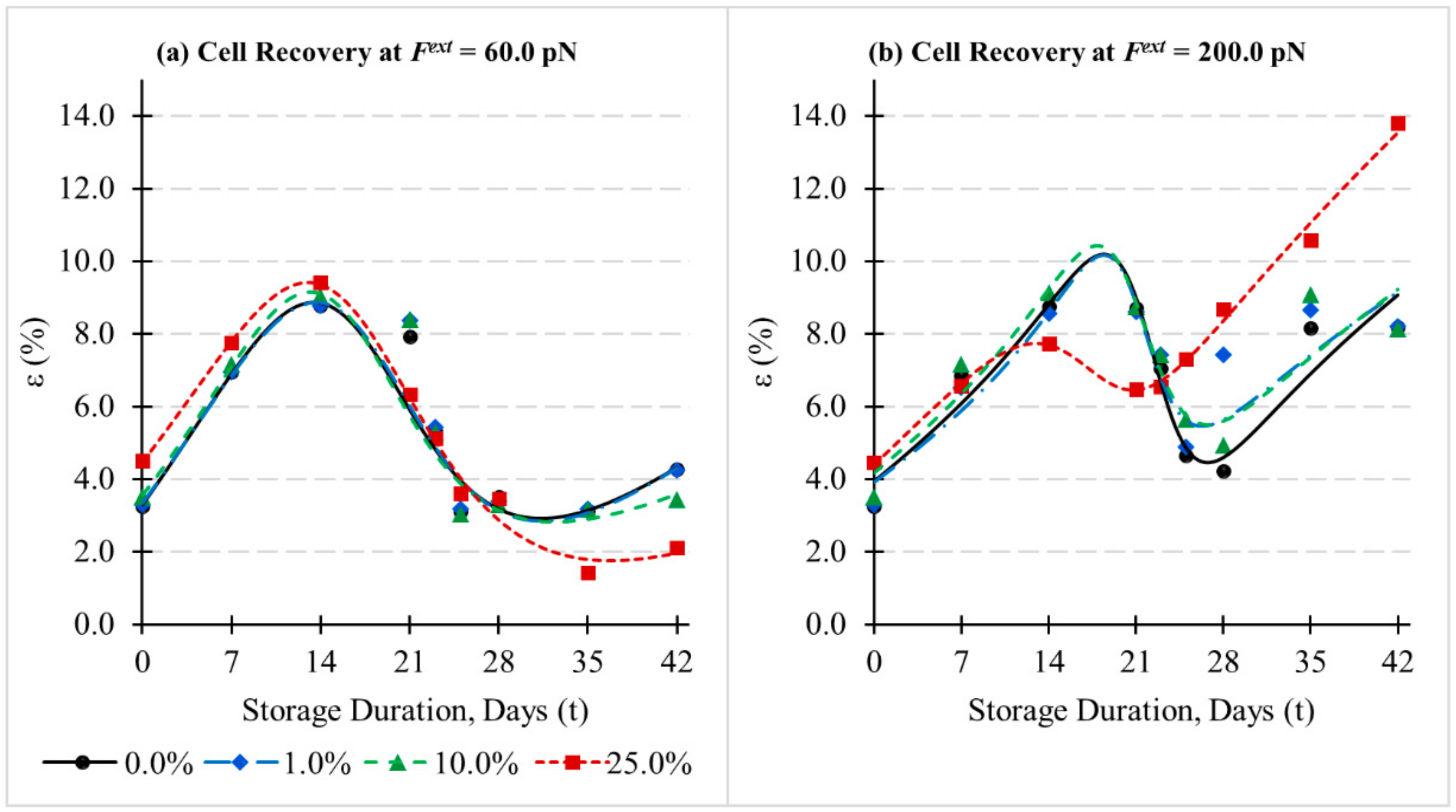
3.4. Influence of Cytoskeletal Actin Junctional Defects during In-Vitro Storage on Cell Recovery following Deformations
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. CG-RBC Membrane Model
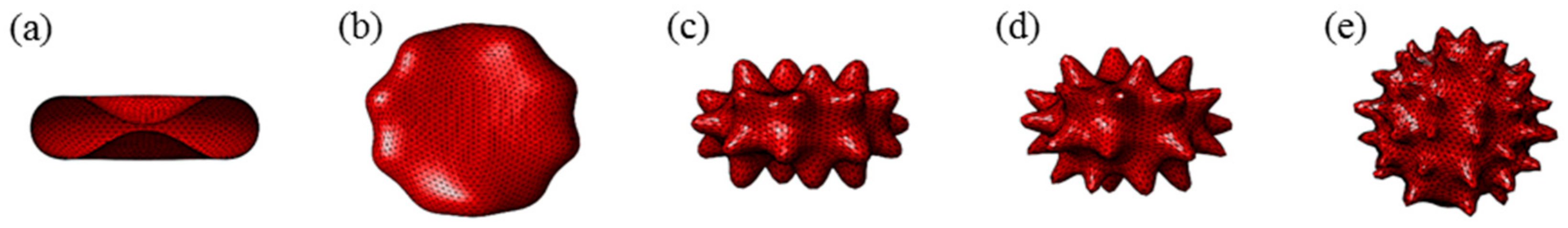
Appendix B. Prediction of Discocyte, Echinocyte I, Echinocyte II, Echinocyte III and Sphero-Echinocyte RBC Morphologies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Value | Source |
|---|---|---|---|
| Reference RBC membrane surface area | 140.0 µm2 | [62,76,93] | |
| Reference total-membrane-curvature for discocyte | 1.85 10−3 m2 | [76] | |
| Reference total-membrane-curvature for echinocyte I | 2.00 10−3 m2 | [76] | |
| Reference total-membrane-curvature for echinocyte II | 5.50 10−3 m2 | [76] | |
| Reference total-membrane-curvature for echinocyte III | 6.00 10−3 m2 | [76] | |
| Reference total-membrane-curvature for sphero-echinocyte | 9.00 10−3 m2 | [76] | |
| Monolayer thickness of the lipid-bilayer | 2.0 nm | [75,76] | |
| Number of links in CG-RBC membrane model | 7680 | [76] | |
| Number of vertices in CG-RBC membrane model | 2562 | [76] | |
| Number of triangles in CG-RBC membrane model | 5120 | [76] | |
| Reference RBC volume | 93.48 µm3 | [62,76,93] | |
| Constraint coefficient for total membrane surface area | 1.0 10−3 Nm−1 | [76] | |
| Constraint coefficient for bilayer-leaflet-area-difference | 300.0 Nm | [76] | |
| Constraint coefficient for total-membrane-curvature | 100.0 Nm | [76] | |
| Constraint coefficient for cell volume | 100.0 Nm−2 | [76] | |
| Constraint coefficient for local membrane surface area | 5.0 10−5 Nm−1 | [76] | |
| Mass of the CG-RBC membrane vertex | 1.0 10−9 kg | [76] | |
| Extensibility of spectrin links | 0.45 | [58,76] | |
| Membrane shear modulus | 4.0 µNm−1 | [58,68,76] | |
| Reference total-membrane-curvature for discocyte | 1.20 10−3 m2 | [76] | |
| Reference total-membrane-curvature for echinocyte I | 1.35 10−3 m2 | [76] | |
| Reference total-membrane-curvature for echinocyte II | 1.90 10−3 m2 | [76] | |
| Reference total-membrane-curvature for echinocyte III | 2.25 10−3 m2 | [76] | |
| Reference total-membrane-curvature for sphero-echinocyte | 3.00 10−3 m2 | [76] | |
| Absolute temperature | 296.15 K | [76] | |
| Membrane viscosity | 1.0 10−7 Nsm−1 | [76] | |
| Exponent of | 2 | [58,76] | |
| Membrane bending modulus | 2.5 10−19 Nm | [57,68,76] |

Appendix C. Measurements of Cell Surface Area and Volume of RBC during In-Vitro Storage—Experimental Observations
| RBC Surface Area (µm2) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Day # | Discocyte | Echinocyte I | Echinocyte II | Echinocyte III | RBC Mean | ||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | ||
| 2 | 130.97 | 10.13 | 128.33 | 16.59 | 135.11 | 5.34 | 112.09 | 12.09 | 126.18 ± 15.08 |
| 7 | 111.80 | 7.33 | 122.26 | 13.27 | 118.58 | 7.27 | 123.05 | 4.78 | 118.87 ± 10.33 |
| 21 | 127.02 | 4.44 | 126.19 | 11.15 | 117.71 | 12.11 | 118.88 | 13.68 | 124.29 ± 10.69 |
| 28 | 106.48 | 10.29 | 106.41 | 7.59 | 115.55 | 2.19 | 112.89 | 10.30 | 109.33 ± 9.26 |
| 48 | 99.41 | 3.64 | 105.86 | 8.82 | 98.66 | 2.69 | 105.12 | 7.56 | 102.88 ± 7.48 |
| RBC Volume (µm3) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Day # | Discocyte | Echinocyte I | Echinocyte II | Echinocyte III | RBC Mean | ||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | ||
| 2 | 106.98 | 24.19 | 96.40 | 22.27 | 113.27 | 17.74 | 84.78 | 13.73 | 99.51 ± 22.68 |
| 7 | 77.15 | 19.88 | 86.39 | 21.73 | 81.69 | 22.75 | 97.06 | 23.51 | 85.03 ± 22.98 |
| 21 | 89.95 | 19.88 | 91.62 | 18.73 | 92.62 | 24.61 | 99.38 | 20.72 | 92.47 ± 20.78 |
| 28 | 77.33 | 18.31 | 79.96 | 15.96 | 90.53 | 24.04 | 87.18 | 16.38 | 83.25 ± 19.24 |
| 48 | 76.20 | 20.38 | 69.32 | 25.61 | 69.97 | 22.96 | 82.00 | 24.92 | 73.87 ± 24.39 |
Appendix D. Implementation of Optical Tweezers Stretching


References
- Park, H.; Lee, S.; Ji, M.; Kim, K.; Son, Y.; Jang, S.; Park, Y. Measuring cell surface area and deformability of individual human red blood cells over blood storage using quantitative phase imaging. Sci. Rep. 2016, 6, 34257. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; Culp-Hill, R.; Reisz, J.A.; Anderson, M.; Fu, X.; Nemkov, T.; Gehrke, S.; Zheng, C.; Kanias, T.; Guo, Y.; et al. Heterogeneity of blood processing and storage additives in different centers impacts stored red blood cell metabolism as much as storage time: Lessons from REDS-III—Omics. Transfusion 2019, 59, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Bosman, G.J.C.G.M.; Werre, J.M.; Willekens, F.L.A.; Novotný, V.M.J. Erythrocyte ageing in vivo and in vitro: Structural aspects and implications for transfusion. Transfus. Med. 2008, 18, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, R.L. Time to revisit red blood cell additive solutions and storage conditions: A role for “omics” analyses. Blood Transfus. 2012, 10, 7–11. [Google Scholar]
- Mustafa, I.; al Marwani, A.; Nasr, K.M.; Kano, N.A.; Hadwan, T. Time dependent assessment of morphological changes: Leukodepleted packed red blood cells stored in SAGM. Biomed. Res. Int. 2016, 2016, 4529434. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; Liumbruno, G.M. Red blood cell storage and clinical outcomes: New insights. Blood Transfus. 2017, 15, 101–103. [Google Scholar] [PubMed]
- D’Alessandro, A.; Gray, A.D.; Sczcepiorkowski, Z.M.; Hansen, K.; Herschel, L.H.; Dumont, L.J. Red blood cell metabolic responses to refrigerated storage, rejuvenation, and frozen storage. Transfusion 2017, 57, 1019–1030. [Google Scholar] [CrossRef]
- Kozlova, E.; Chernysh, A.; Moroz, V.; Sergunova, V.; Gudkova, O.; Manchenko, E. Morphology, membrane nanostructure and stiffness for quality assessment of packed red blood cells. Sci. Rep. 2017, 7, 7846. [Google Scholar] [CrossRef]
- Roussel, C.; Dussiot, M.; Marin, M.; Morel, A.; Ndour, P.A.; Duez, J.; Kim, C.L.V.; Hermine, O.; Colin, Y.; Buffet, P.A.; et al. Spherocytic shift of red blood cells during storage provides a quantitative whole cell–based marker of the storage lesion. Transfusion 2017, 57, 1007–1018. [Google Scholar] [CrossRef]
- Sierra, F.D.A.; Melzak, K.A.; Janetzko, K.; Klueter, H.; Suhr, H.; Bieback, K.; Wiedemann, P. Flow morphometry to assess the red blood cell storage lesion. Cytom. Part A 2017, 91, 874–882. [Google Scholar] [CrossRef]
- Tuo, W.W.; Wang, D.; Liang, W.J.; Huang, Y.X. How cell number and cellular properties of blood-banked red blood cells of different cell ages decline during storage. PLoS ONE 2014, 9, 105692. [Google Scholar] [CrossRef]
- Holme, S. Current issues related to the quality of stored RBCs. Transfus. Apher. Sci. 2005, 33, 55–61. [Google Scholar] [CrossRef]
- Zehnder, L.; Schulzki, T.; Goede, J.S.; Hayes, J.; Reinhart, W.H. Erythrocyte storage in hypertonic (SAGM) or isotonic (PAGGSM) conservation medium: Influence on cell properties. Vox Sang. 2008, 95, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Bhaduri, B.; Kandel, M.; Brugnara, C.; Tangella, K.; Popescu, G. Optical assay of erythrocyte function in banked blood. Sci. Rep. 2014, 4, 6211. [Google Scholar] [CrossRef] [PubMed]
- Antonelou, M.H.; Kriebardis, A.G.; Stamoulis, K.E.; Economou-Petersen, E.; Margaritis, L.H.; Papassideri, I.S. Red blood cell aging markers during storage in citrate-phosphate-dextrose-saline-adenine-glucose-mannitol. Transfusion 2010, 50, 376–389. [Google Scholar] [CrossRef] [PubMed]
- Tzounakas, V.L.; Kriebardis, A.G.; Seghatchian, J.; Papassideri, I.S.; Antonelou, M.H. Unraveling the Gordian knot: Red blood cell storage lesion and transfusion outcomes. Blood Transfus. 2017, 15, 126–130. [Google Scholar] [PubMed]
- Xu, Z.; Pu, H.; Xie, S.; Wang, C.; Sun, Y. Microfluidic measurement of RBC bending stiffness changes in blood storage. In Proceedings of the 19th International Conference on Solid-State Sensors, Actuators and Microsystems (TRANSDUCERS), Kaohsiung, Taiwan, 18–22 June 2017; pp. 734–737. [Google Scholar]
- Xu, Z.; Zheng, Y.; Wang, X.; Shehata, N.; Wang, C.; Sun, Y. Stiffness Increase of red blood cells during storage. Microsyst. Nanoeng. 2018, 4, 17103. [Google Scholar] [CrossRef]
- Yoshida, T.; Prudent, M.; D’Alessandro, A. Red blood cell storage lesion: Causes and potential clinical consequences. Blood Transfus. 2019, 17, 27–52. [Google Scholar]
- Frank, S.M.; Abazyan, B.; Ono, M.; Hogue, C.W.; Cohen, D.B.; Berkowitz, D.E.; Ness, P.M.; Barodka, V.M. Decreased erythrocyte deformability after transfusion and the effects of erythrocyte storage duration. Anesth. Analg. 2013, 116, 975–981. [Google Scholar] [CrossRef]
- Obrador, R.; Musulin, S.; Hansen, B. Red blood cell storage lesion. J. Vet. Emerg. Crit. Care 2015, 25, 187–199. [Google Scholar] [CrossRef]
- Antonelou, M.H.; Seghatchian, J. Insights into red blood cell storage lesion: Toward a new appreciation. Transfus. Apher. Sci. 2016, 55, 292–301. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A. Red blood cell storage lesion. ISBT Sci. Ser. 2017, 12, 207–213. [Google Scholar] [CrossRef]
- Matthews, K.; Myrand-Lapierre, M.-E.; Ang, R.R.; Duffy, S.P.; Scott, M.D.; Ma, H. Microfluidic deformability analysis of the red cell storage lesion. J. Biomech. 2015, 48, 4065–4072. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Liu, Y.; Zhang, B.; Tian, K.; Zhu, P.; Lu, H.; Tang, Q. Study of in vitro RBCs membrane elasticity with AOD scanning optical tweezers. Biomed. Opt. Express 2017, 8, 384–394. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Chen, J.; Cui, T.; Shehata, N.; Wang, C.; Sun, Y. Characterization of red blood cell deformability change during blood storage. Lab Chip 2014, 14, 577–583. [Google Scholar] [CrossRef]
- Dao, M.; Lim, C.T.; Suresh, S. Mechanics of the human red blood cell deformed by optical tweezers. J. Mech. Phys. Solids 2003, 51, 2259–2280. [Google Scholar] [CrossRef]
- Kuzman, D.; Svetina, S.; Waugh, R.E.; Žekš, B. Elastic properties of the red blood cell membrane that determine echinocyte deformability. Eur. Biophys. J. 2004, 33, 1–15. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, K.; Park, Y.K. Measurement techniques for red blood cell deformability: Recent advances. In Blood Cell—An Overview of Studies in Hematology; Moschandreou, T.E., Ed.; InTech: New York, NY, USA, 2012; Volume 10, pp. 167–194. [Google Scholar]
- Chang, H.-Y.; Li, X.; Li, H.; Karniadakis, G.E. MD/DPD Multiscale Framework for Predicting Morphology and Stresses of Red Blood Cells in Health and Disease. PLoS Comput. Biol. 2016, 12, 1005173. [Google Scholar] [CrossRef]
- Li, X.; Dao, M.; Lykotrafitis, G.; Karniadakis, G.E. Biomechanics and biorheology of red blood cells in sickle cell anemia. J. Biomech. 2016, 50, 34–41. [Google Scholar] [CrossRef]
- Li, H.; Lu, L.; Li, X.; Buffet, P.A.; Dao, M.; Karniadakis, G.E.; Suresh, S. Mechanics of diseased red blood cells in human spleen and consequences for hereditary blood disorders. Proc. Natl. Acad. Sci. USA 2018, 115, 9574–9579. [Google Scholar] [CrossRef]
- Wang, Y.; You, G.; Chen, P.; Li, J.; Chen, G.; Wang, B.; Li, P.; Han, D.; Zhou, H.; Zhao, L. The mechanical properties of stored red blood cells measured by a convenient microfluidic approach combining with mathematic model. Biomicrofluidics 2016, 10, 024104. [Google Scholar] [CrossRef] [PubMed]
- Card, R.T.; Mohandas, N.; Mollison, P.L. Relationship of post-transfusion viability to deformability of stored red-cells. Br. J. Haematol. 1983, 53, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Van de Watering, L. More data on red blood cell storage could clarify confusing clinical outcomes. Transfusion 2014, 54, 501–502. [Google Scholar] [CrossRef] [PubMed]
- Huruta, R.R.; Barjas-Castro, M.L.; Saad, S.T.O.; Costa, F.F.; Fontes, A.; Barbosa, L.C.; Cesar, C.L. Mechanical properties of stored red blood cells using optical tweezers. Blood 1998, 92, 2975–2977. [Google Scholar] [CrossRef]
- Ramirez-Arcos, S.; Marks, D.C.; Acker, J.P.; Sheffield, W.P. Quality and safety of blood products. J. Blood Transfus. 2016, 2016, 2482157. [Google Scholar] [CrossRef]
- Czerwinska, J.; Wolf, S.M.; Mohammadi, H.; Jeney, S. Red blood cell aging during storage, studied using optical tweezers experiment. Cell. Mol. Bioeng. 2015, 8, 258–266. [Google Scholar] [CrossRef]
- Dao, M.; Li, J.; Suresh, S. Molecularly based analysis of deformation of spectrin network and human erythrocyte. Mater. Sci. Eng. C 2006, 26, 1232–1244. [Google Scholar] [CrossRef]
- Lim, C.T.; Dao, M.; Suresh, S.; Sow, C.H.; Chew, K.T. Large deformation of living cells using laser traps. Acta Mater. 2004, 52, 1837–1845. [Google Scholar] [CrossRef]
- Mills, J.P.; Qie, L.; Dao, M.; Lim, C.T.; Suresh, S. Nonlinear elastic and viscoelastic deformation of the human red blood cell with optical tweezers. Mech. Chem. Biosyst. 2004, 1, 169–180. [Google Scholar]
- Fedosov, D.A.; Dao, M.; Karniadakis, G.E.; Suresh, S. Computational biorheology of human blood flow in health and disease. Ann. Biomed. Eng. 2014, 42, 368–387. [Google Scholar] [CrossRef]
- Fedosov, D.A.; Noguchi, H.; Gompper, G. Multiscale modeling of blood flow: From single cells to blood rheology. Biomech. Model. Mechanobiol. 2014, 13, 239–258. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.-G.; Wu, H.-A.; Zhou, X.-Z.; Wang, X.-X. Coarse-grained molecular dynamics simulation of a red blood cell. Chin. Phys. Lett. 2010, 27, 028704. [Google Scholar]
- Polwaththe-Gallage, H.N.; Saha, S.C.; Gu, Y. Deformation of a three-dimensional red blood cell in a stenosed microcapillary. In Proceedings of the 8th Australasian Congress on Applied Mechanics (ACAM-8), Melbourne, Australia, 23–26 November 2014. [Google Scholar]
- Yazdani, A.; Li, X.; Karniadakis, G.E. Dynamic and rheological properties of soft biological cell suspensions. Rheol. Acta 2016, 55, 433–449. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Park, Y.; Best, C.A.; Badizadegan, K.; Dasari, R.R.; Feld, M.S.; Kuriabova, T.; Henle, M.L.; Levine, A.J.; Popescu, G. Measurement of red blood cell mechanics during morphological changes. Proc. Natl. Acad. Sci. USA 2010, 107, 6731–6736. [Google Scholar] [CrossRef] [PubMed]
- Israelachvili, J.N. (Ed.) Intermolecular and Surface Forces, 3rd ed.; Academic Press: San Diego, CA, USA, 2011. [Google Scholar]
- Iglič, A.; Kralj-Iglič, V.; Hägerstrand, H. Amphiphile induced echinocyte-spheroechinoeyte transformation of red blood cell shape. Eur. Biophys. J. 1998, 27, 335–339. [Google Scholar] [PubMed]
- Mukhopadhyay, R.; Lim, G.; Wortis, M. Echinocyte shapes: Bending, stretching, and shear determine spicule shape and spacing. Biophys. J. 2002, 82, 1756–1772. [Google Scholar] [CrossRef]
- Rudenko, S.V. Erythrocyte morphological states, phases, transitions and trajectories. Biochim. Biophys. Acta (BBA)—Biomembr. 2010, 1798, 1767–1778. [Google Scholar] [CrossRef]
- Rudenko, S.V.; Saeid, M.K. Reconstruction of erythrocyte shape during modified morphological response. Biochemistry (Moscow) 2010, 75, 1025–1031. [Google Scholar] [CrossRef]
- Etcheverry, S.; Gallardo, M.J.; Solano, P.; Suwalsky, M.; Mesquita, O.N.; Saavedra, C. Real-time study of shape and thermal fluctuations in the echinocyte transformation of human erythrocytes using defocusing microscopy. J. Biomed. Opt. 2012, 17, 106013. [Google Scholar] [CrossRef]
- Chen, M.; Boyle, F.J. An enhanced spring-particle model for red blood cell structural mechanics: Application to the stomatocyte–discocyte–echinocyte transformation. J. Biomech. Eng. 2017, 139, 121009. [Google Scholar] [CrossRef]
- Wong, P. A basis of echinocytosis and stomatocytosis in the disc–sphere transformations of the erythrocyte. J. Theor. Biol. 1999, 196, 343–361. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Asaro, R.J.; Zhu, Q. Multiscale simulation of erythrocyte membranes. Phys. Rev. E 2010, 81, 031904. [Google Scholar] [CrossRef] [PubMed]
- Fedosov, D.A.; Caswell, B.; Karniadakis, G.E. Coarse-grained red blood cell model with accurate mechanical properties, rheology and dynamics. In Proceedings of the Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Minneapolis, MN, USA, 2–6 September 2009; Volume 1–20, pp. 4266–4269. [Google Scholar]
- Fedosov, D.A.; Caswell, B.; Karniadakis, G.E. Systematic coarse-graining of spectrin-level red blood cell models. Comput. Methods Appl. Mech. Eng. 2010, 199, 1937–1948. [Google Scholar] [CrossRef] [PubMed]
- Fedosov, D.A.; Caswell, B.; Karniadakis, G.E. A multiscale red blood cell model with accurate mechanics, rheology, and dynamics. Biophys. J. 2010, 98, 2215–2225. [Google Scholar] [CrossRef]
- Peng, Z.; Asaro, R.J.; Zhu, Q. Multiscale modelling of erythrocytes in Stokes flow. J. Fluid Mech. 2011, 686, 299–337. [Google Scholar] [CrossRef]
- Peng, Z.; Li, X.; Pivkin, I.V.; Dao, M.; Karniadakis, G.E.; Suresh, S. Lipid bilayer and cytoskeletal interactions in a red blood cell. Proc. Natl. Acad. Sci. USA 2013, 110, 13356–13361. [Google Scholar] [CrossRef]
- Tomaiuolo, G. Biomechanical properties of red blood cells in health and disease towards microfluidics. Biomicrofluidics 2014, 8, 051501. [Google Scholar] [CrossRef]
- Ju, M.; Ye, S.S.; Namgung, B.; Cho, S.; Low, H.T.; Leo, H.L.; Kim, S. A review of numerical methods for red blood cell flow simulation. Comput. Methods Biomech. Biomed. Eng. 2015, 18, 130–140. [Google Scholar] [CrossRef]
- Monzel, C.; Sengupta, K. Measuring shape fluctuations in biological membranes. J. Phys. D—Appl. Phys. 2016, 49, 243002. [Google Scholar] [CrossRef]
- Bento, D.; Rodrigues, R.; Faustino, V.; Pinho, D.; Fernandes, C.; Pereira, A.; Garcia, V.; Miranda, J.; Lima, R. Deformation of red blood cells, air bubbles, and droplets in microfluidic devices: Flow visualizations and measurements. Micromachines 2018, 9, 151. [Google Scholar] [CrossRef]
- Polwaththe-Gallage, H.N.; Saha, S.C.; Sauret, E.; Flower, R.; Senadeera, W.; Gu, Y. SPH-DEM approach to numerically simulate the deformation of three-dimensional RBCs in non-uniform capillaries. Biomed. Eng. Online 2016, 15, 354–370. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Barns, S.; Balanant, M.A.; Sauret, E.; Flower, R.; Saha, S.; Gu, Y. Investigation of red blood cell mechanical properties using AFM indentation and coarse-grained particle method. Biomed. Eng. Online 2017, 16, 140. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, H.; Chang, H.-Y.; Lykotrafitis, G.; Karniadakis, G.E. Computational biomechanics of human red blood cells in hematological disorders. J. Biomech. Eng. 2017, 139, 021008. [Google Scholar] [CrossRef] [PubMed]
- Ademiloye, A.S.; Zhang, L.W.; Liew, K.M. A multiscale framework for large deformation modeling of RBC membranes. Comput. Methods Appl. Mecha. Eng. 2018, 329, 144–167. [Google Scholar] [CrossRef]
- Ademiloye, A.S.; Zhang, L.W.; Liew, K.M. Atomistic–continuum model for probing the biomechanical properties of human erythrocyte membrane under extreme conditions. Comput. Methods Appl. Mech. Eng. 2017, 325, 22–36. [Google Scholar] [CrossRef]
- Hénon, S.; Lenormand, G.; Richert, A.; Gallet, F. A new determination of the shear modulus of the human erythrocyte membrane using optical tweezers. Biophys. J. 1999, 76, 1145–1151. [Google Scholar] [CrossRef]
- Liang, Y.; Xiang, Y.; Lamstein, J.; Bezryadina, A.; Chen, Z. Cell deformation and assessment with tunable “tug-of-war” optical tweezers. In Conference on Lasers and Electro-Optics; Optical Society of America: San Jose, CA, USA, 2019. [Google Scholar]
- Imai, Y.; Omori, T.; Shimogonya, Y.; Yamaguchi, T.; Ishikawa, T. Numerical methods for simulating blood flow at macro, micro, and multi scales. J. Biomech. 2016, 49, 2221–2228. [Google Scholar] [CrossRef]
- Lim, G.H.W.; Wortis, M.; Mukhopadhyay, R. Stomatocyte–discocyte–echinocyte sequence of the human red blood cell: Evidence for the bilayer– couple hypothesis from membrane mechanics. Proc. Natl. Acad. Sci. USA 2002, 99, 16766–16769. [Google Scholar] [CrossRef]
- Lim, G.H.W.; Wortis, M.; Mukhopadhyay, R. Red blood cell shapes and shape transformations: Newtonian mechanics of a composite membrane: Sections 2.5–2.8. Soft Matter 2009, 4, 139–204. [Google Scholar]
- Geekiyanage, N.M.; Balanant, M.A.; Sauret, E.; Saha, S.; Flower, R.; Lim, C.T.; Gu, Y. A coarse-grained red blood cell membrane model to study stomatocyte-discocyte-echinocyte morphologies. PLoS ONE 2019, 14, 0215447. [Google Scholar] [CrossRef]
- Bessis, M. Red cell shapes. An illustrated classification and its rationale. In Red Cell Shape; Springer: Berlin/Heidelberg, Germany, 1973; Volume 12, pp. 721–746, no. 6. [Google Scholar]
- Jaferzadeh, K.; Moon, I. Quantitative investigation of red blood cell three-dimensional geometric and chemical changes in the storage lesion using digital holographic microscopy. J. Biomed. Opt. 2015, 20, 111218. [Google Scholar] [CrossRef] [PubMed]
- Veale, M.F.; Healey, G.; Sparrow, R.L. Effect of additive solutions on red blood cell (RBC) membrane properties of stored RBCs prepared from whole blood held for 24 hours at room temperature. Transfusion 2011, 51, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.; Edeskär, T.; Knutsson, S. Particle shape quantities and measurement techniques: A review. Electron. J. Geotech. Eng. 2013, 18, 169–198. [Google Scholar]
- Brecher, G.; Bessis, M. Present status of spiculed red cells and their relationship to the discocyte-echinocyte transformation: A critical review. Blood 1972, 40, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lykotrafitis, G. Vesiculation of healthy and defective red blood cells. Phys. Rev. E 2015, 92, 012715. [Google Scholar] [CrossRef]
- Li, Y.; Wen, C.; Xie, H.; Ye, A.; Yin, Y. Mechanical property analysis of stored red blood cell using optical tweezers. Colloids Surf. B Biointerfaces 2009, 70, 169–173. [Google Scholar] [CrossRef]
- Li, H.; Lykotrafitis, G. Erythrocyte membrane model with explicit description of the lipid bilayer and the spectrin network. Biophys. J. 2014, 107, 642–653. [Google Scholar] [CrossRef]
- Geekiyanage, N.M.; Sauret, E.; Saha, S.C.; Flower, R.L.; Gu, Y.T. Deformation behaviour of stomatocyte, discocyte and echinocyte red blood cell morphologies during optical tweezers stretching. (in eng). Biomech. Model. Mechanobiol. 2020, 1–17. (In English) [Google Scholar] [CrossRef]
- Li, J.; Dao, M.; Lim, C.T.; Suresh, S. Spectrin-level modeling of the cytoskeleton and optical tweezers stretching of the erythrocyte. Biophys. J. 2005, 88, 3707–3719. [Google Scholar] [CrossRef]
- Suresh, S.; Spatz, J.; Mills, J.P.; Micoulet, A.; Dao, M.; Lim, C.T.; Beil, M.; Seufferlein, T. Connections between single-cell biomechanics and human disease states: Gastrointestinal cancer and malaria. Acta Biomater. 2005, 1, 15–30. [Google Scholar] [CrossRef]
- Helfrich, W. Blocked lipid exchange in bilayers and its possible influence on the shape of vesicles. Z. Naturforschung 1974, 29, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Svetina, S.; Žekš, B. Membrane bending energy and shape determination of phospholipid vesicles and red blood cells. Eur. Biophys. J. 1989, 17, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Seifert, U.; Berndl, K.; Lipowsky, R. Shape transformations of vesicles: Phase diagram for spontaneous- curvature and bilayer-coupling models. Phys. Rev. A 1991, 44, 1182–1202. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Seifert, U.; Wortis, M.; Döbereiner, H.-G. Budding transitions of fluid-bilayer vesicles: The effect of area-difference elasticity. Phys. Rev. E 1994, 49, 5389–5407. [Google Scholar] [CrossRef]
- Nakamura, M.; Bessho, S.; Wada, S. Analysis of red blood cell deformation under fast shear flow for better estimation of hemolysis. Int. J. Numer. Methods Biomed. Eng. 2014, 30, 42–54. [Google Scholar] [CrossRef]
- Mohandas, N.; Gallagher, P.G. Red cell membrane: Past, present, and future. Blood 2008, 112, 3939–3948. [Google Scholar] [CrossRef]












| Days (t) | CG-RBC Membrane Model Parameters at | RBC Shape | |||||
|---|---|---|---|---|---|---|---|
(%) | (%) | ||||||
| 0 | 1.000 | 1.000 | 1.000 | 1.000 | 0.120 | 0.185 |  |
| 7 | 1.000 | 1.000 | 0.976 | 0.974 | 0.120 | 0.185 |  |
| 14 | 1.000 | 1.000 | 0.953 | 0.947 | 0.120 | 0.185 |  |
| 21 | 1.452 | 1.956 | 0.929 | 0.921 | 0.135 | 0.200 |  |
| 23 | 1.905 | 2.911 | 0.923 | 0.913 | 0.190 | 0.550 |  |
| 25 | 2.357 | 3.867 | 0.916 | 0.906 | 0.225 | 0.600 |  |
| 28 | 2.583 | 4.345 | 0.906 | 0.895 | 0.300 | 0.900 |  |
| 35 | 2.583 | 4.345 | 0.862 | 0.835 | 0.300 | 0.900 |  |
| 42 | 2.583 | 4.345 | 0.818 | 0.775 | 0.300 | 0.900 |  |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geekiyanage, N.; Sauret, E.; Saha, S.; Flower, R.; Gu, Y. Modelling of Red Blood Cell Morphological and Deformability Changes during In-Vitro Storage. Appl. Sci. 2020, 10, 3209. https://doi.org/10.3390/app10093209
Geekiyanage N, Sauret E, Saha S, Flower R, Gu Y. Modelling of Red Blood Cell Morphological and Deformability Changes during In-Vitro Storage. Applied Sciences. 2020; 10(9):3209. https://doi.org/10.3390/app10093209
Chicago/Turabian StyleGeekiyanage, Nadeeshani, Emilie Sauret, Suvash Saha, Robert Flower, and YuanTong Gu. 2020. "Modelling of Red Blood Cell Morphological and Deformability Changes during In-Vitro Storage" Applied Sciences 10, no. 9: 3209. https://doi.org/10.3390/app10093209
APA StyleGeekiyanage, N., Sauret, E., Saha, S., Flower, R., & Gu, Y. (2020). Modelling of Red Blood Cell Morphological and Deformability Changes during In-Vitro Storage. Applied Sciences, 10(9), 3209. https://doi.org/10.3390/app10093209