Recurrent Water Deficit and Epigenetic Memory in Medicago sativa L. Varieties



 , ,
, ,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Calculation of Substrate Pot Capacity
2.3. Shoot and Root Dry Weight and Shoot and Root Water Content Determination
2.4. Sampling and DNA Extraction
2.5. MSAP Procedure
2.6. Data Collection and Statistical Analysis
3. Results
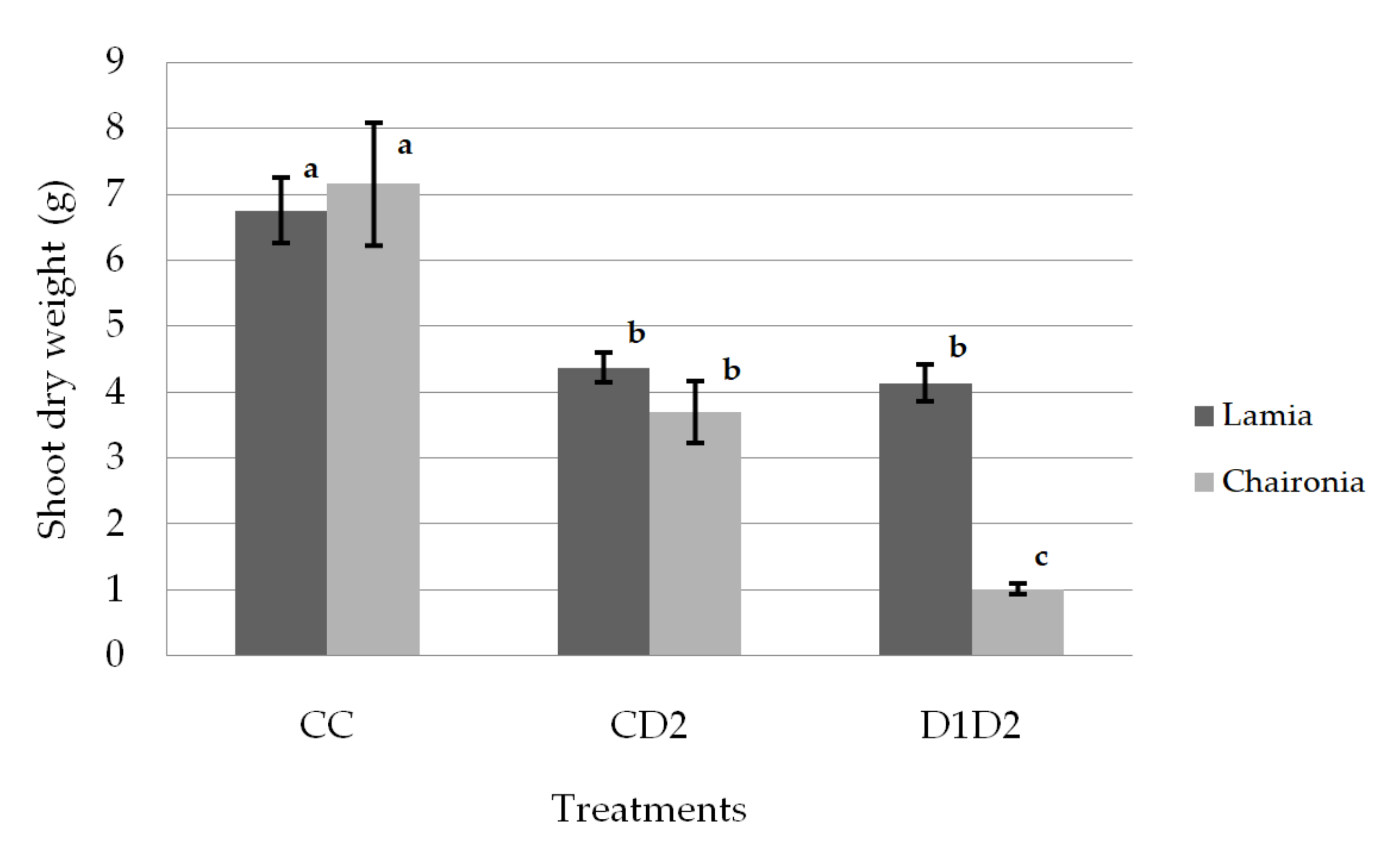
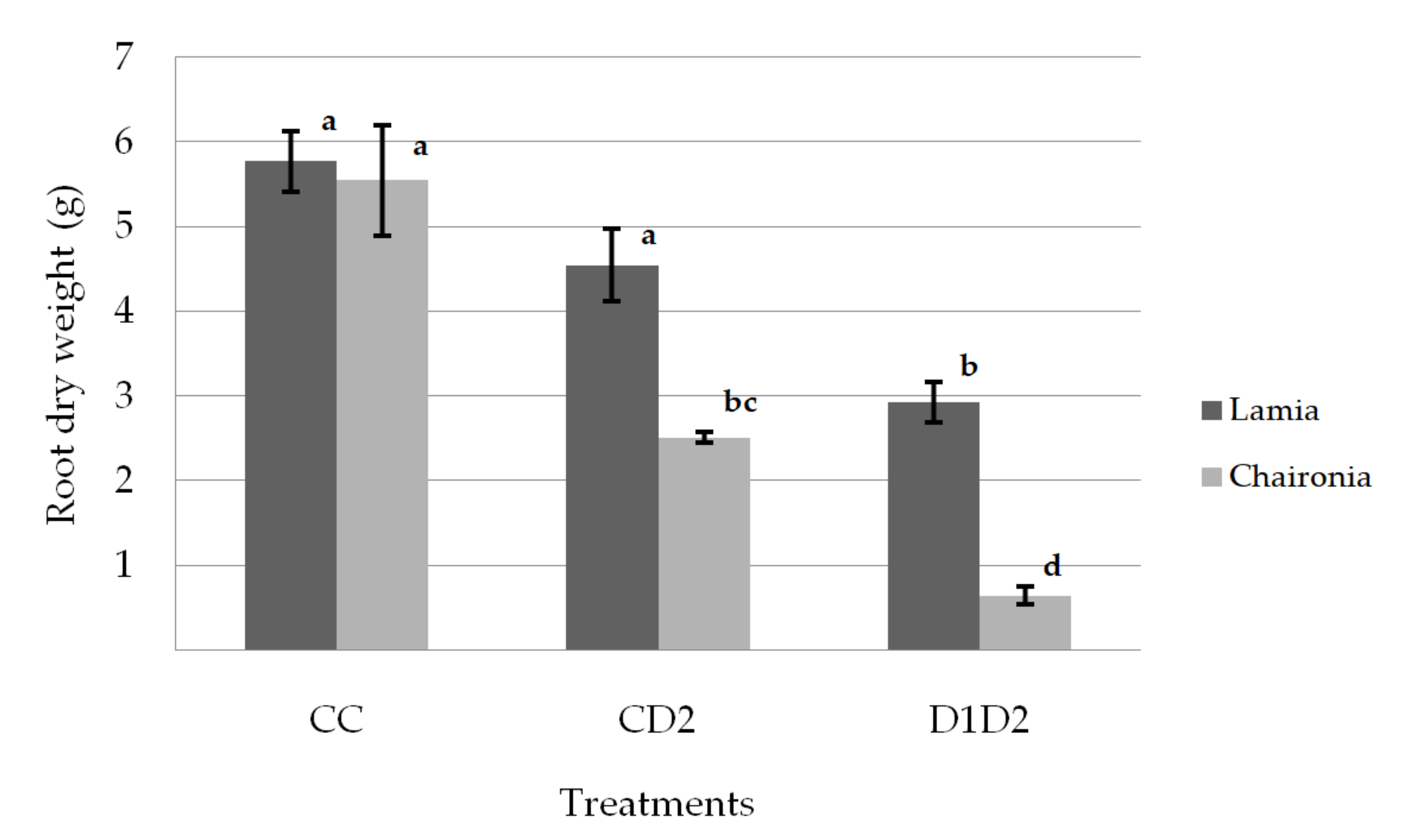
3.1. Shoot and Root Dry Weight
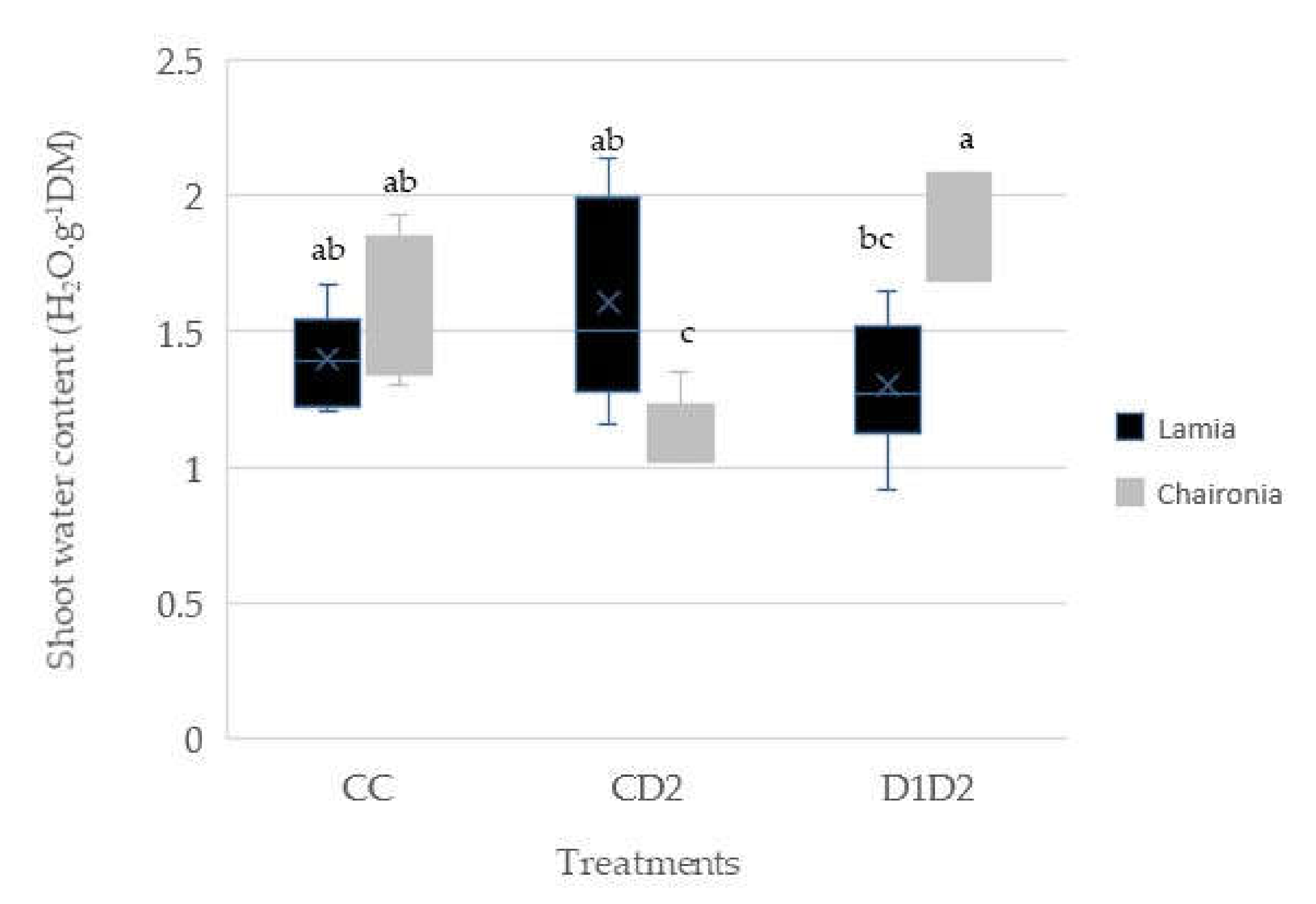
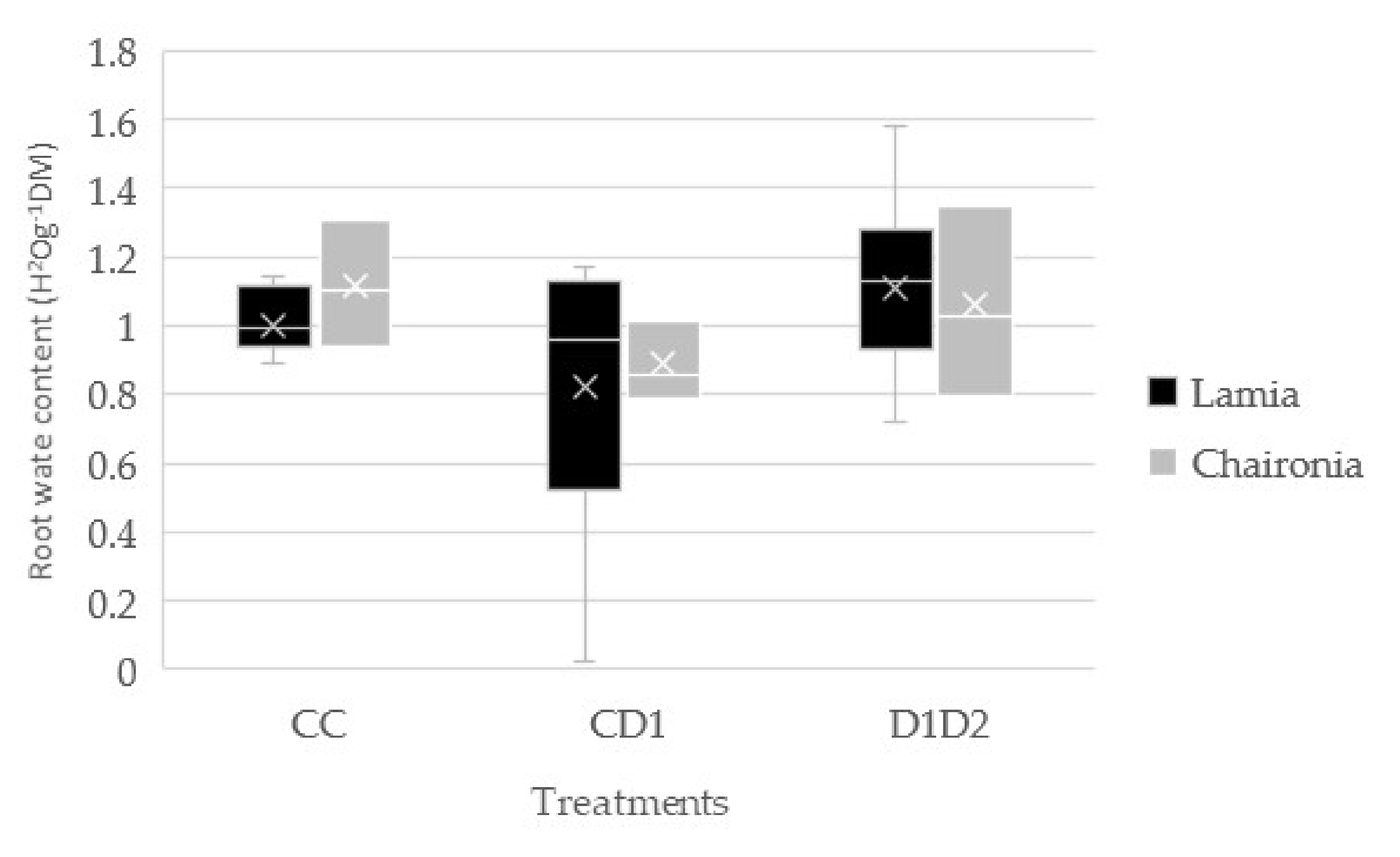
3.2. Shoot and Root Water Content
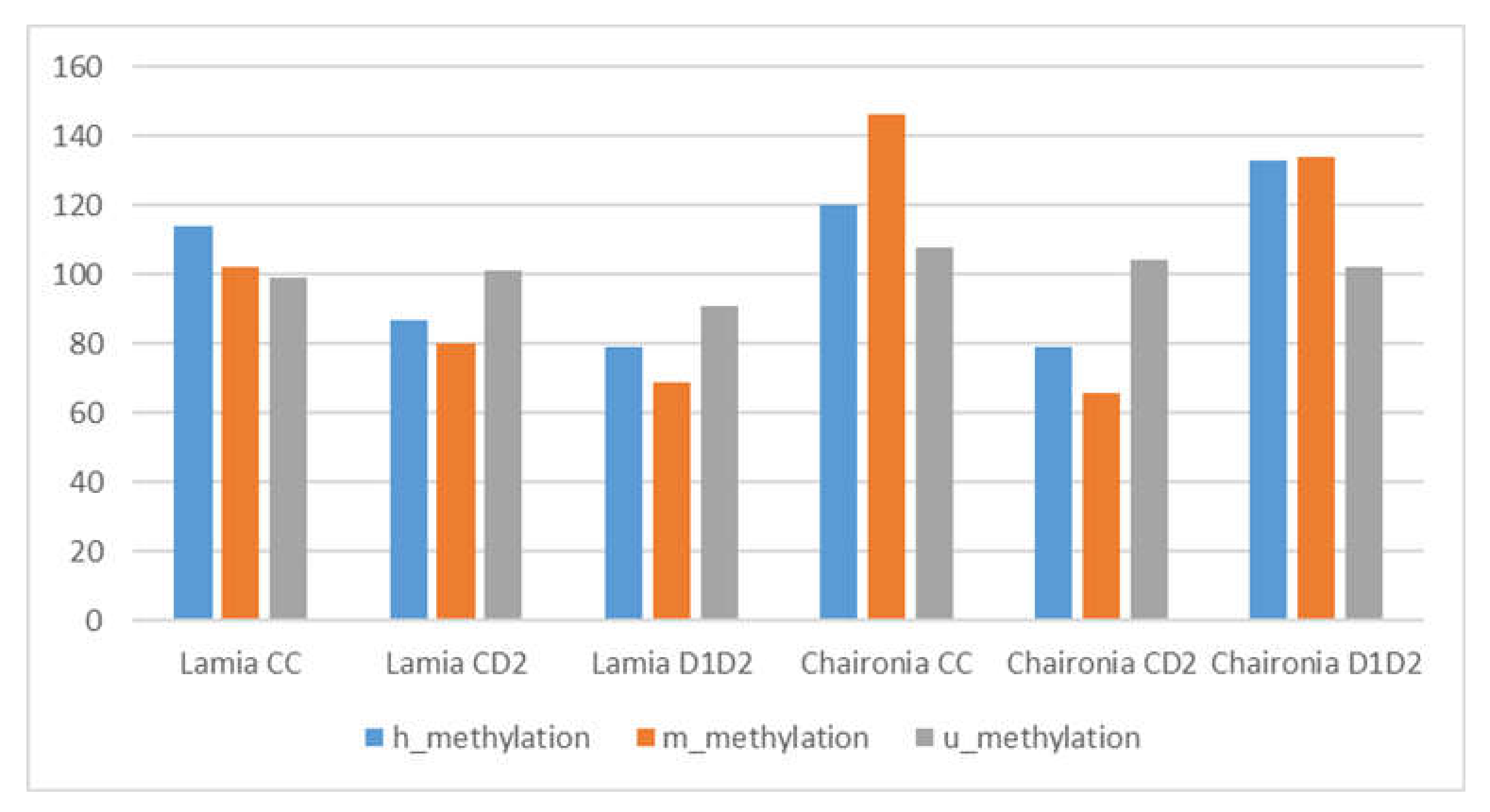
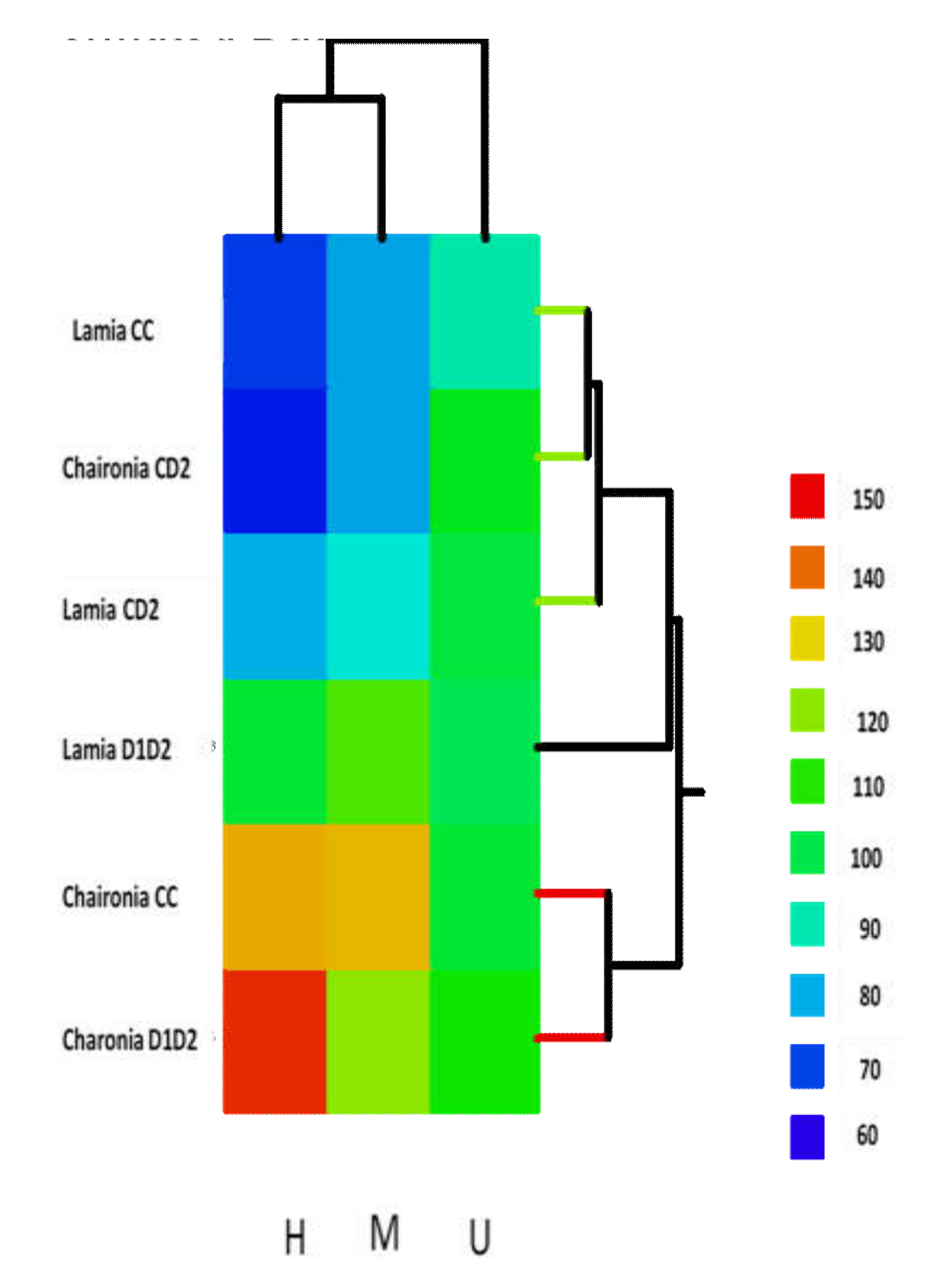
3.3. MSAP Results
- (a)
- For h alleles, no significant differences were found between Lamia treatments, while for Chaironia significant differences were found between CC/CD2, and CD2/D1D2 treatments.
- (b)
- For m alleles, significant differences were found for Lamia CC/Lamia D1D2 and for Chaironia CC/Chaironia CD2, Chaironia CD2/Chaironia D1D2.
- (c)
- For total methylation (h+m), significant differences were found between Lamia CC/Lamia D1D2, Chaironia CC/Chaironia CD2, Chaironia CD2/Chaironia D1D2.
3.4. Among Different Treatments and between Varieties
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Matiu, M.; Ankerst, D.P.; Menzel, A. Interactions between temperature and drought in global and regional crop yield variability during 1961–2014. PLoS ONE 2017, 12, e0178339. [Google Scholar] [CrossRef]
- Singer, S.D.; Hannoufa, A.; Acharya, S. Molecular improvement of alfalfa for enhanced productivity and adaptability in a changing environment. Plant Cell Environ. 2018, 41, 1955–1971. [Google Scholar] [CrossRef]
- Gallusci, P.; Dai, Z.; Génard, M.; Gauffretau, A.; Leblanc-Fournier, N.; Richard-Molard, C.; Vile, D.; Brunel-Muguet, S. Epigenetics for Plant Improvement: Current Knowledge and Modeling Avenues. Trends Plant Sci. 2017, 22, 610–623. [Google Scholar] [CrossRef]
- Thiebaut, F.; Hemerly, A.S.; Ferreira, P.C.G. A Role for Epigenetic Regulation in the Adaptation and Stress Responses of Non-model Plants. Trends Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Springer, N.M. Epigenetics and crop improvement. Trends Genet. 2013, 29, 241–247. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Li, Z. Epigenetic Environmental Memories in Plants: Establishment, Maintenance, and Reprogramming. Trends Genet. 2018, 34, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Kapazoglou, A.; Ganopoulos, I.; Tani, E.; Tsaftaris, A. Chapter Nine—Epigenetics, Epigenomics and Crop Improvement. In Advances in Botanical Research; Kuntz, M., Ed.; Academic Press: Cambridge, MA, USA, 2018; Volume 86, pp. 287–324. [Google Scholar]
- Mozgová, I.; Mikulski, P.; Pecinka, A.; Farrona, S. Epigenetic Mechanisms of Abiotic Stress Response and Memory in Plants. In Epigenetics in Plants of Agronomic Importance: Fundamentals and Applications; Alvarez-Venegas, R., De-la-Peña, C., Casas-Mollano, J., Eds.; Springer: Cham, Switzerland, 2019; pp. 1–64. [Google Scholar]
- Saraswat, S.; Yadav, A.K.; Sirohi, P.; Singh, N.K. Role of epigenetics in crop improvement: Water and heat stress. J. Plant Biol. 2017, 60, 231–240. [Google Scholar] [CrossRef]
- Zhang, H.; Lang, Z.; Zhu, J.-K. Dynamics and function of DNA methylation in plants. Nat. Rev. Mol. Cell Biol. 2018, 19, 489–506. [Google Scholar] [CrossRef]
- Iqbal, K.; Jin, S.-G.; Pfeifer, G.P.; Szabó, P.E. Reprogramming of the paternal genome upon fertilization involves genome-wide oxidation of 5-methylcytosine. Proc. Natl. Acad. Sci. USA 2011, 108, 3642–3647. [Google Scholar] [CrossRef]
- Tang, X.-M.; Tao, X.; Wang, Y.; Ma, D.-W.; Li, D.; Yang, H.; Ma, X.-R. Analysis of DNA methylation of perennial ryegrass under drought using the methylation-sensitive amplification polymorphism (MSAP) technique. Mol. Genet. Genom. MGG 2014, 289, 1075–1084. [Google Scholar]
- Yaish, M.; Peng, M.; Rothstein, S. Global DNA Methylation Analysis Using Methyl-Sensitive Amplification Polymorphism (MSAP). Methods Mol. Biol. 2014, 1062, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Crisp, P.A.; Ganguly, D.; Eichten, S.R.; Borevitz, J.O.; Pogson, B.J. Reconsidering plant memory: Intersections between stress recovery, RNA turnover, and epigenetics. Sci. Adv. 2016, 2, e1501340. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-S.; Pan, Y.-J.; Zhao, X.-Q.; Dwivedi, D.; Zhu, L.-H.; Ali, J.; Fu, B.-Y.; Li, Z.-K. Drought-induced site-specific DNA methylation and its association with drought tolerance in rice (Oryza sativa L.). J. Exp. Bot. 2011, 62, 1951–1960. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Fromm, M.; Avramova, Z. Multiple exposures to drought ’train’ transcriptional responses in Arabidopsis. Nat. Commun. 2012, 3, 740. [Google Scholar] [CrossRef] [PubMed]
- Tricker, P.; Rodríguez López, C.; Hadley, P.; Wagstaff, C.; Wilkinson, M. Pre-conditioning the epigenetic response to high vapor pressure deficit increases the drought tolerance of Arabidopsis thaliana. Plant Signal. Behav. 2013, 8, e25974. [Google Scholar] [CrossRef]
- Tricker, P.J.; López, C.M.R.; Gibbings, G.; Hadley, P.; Wilkinson, M.J. Transgenerational, Dynamic Methylation of Stomata Genes in Response to Low Relative Humidity. Int. J. Mol. Sci. 2013, 14, 6674–6689. [Google Scholar] [CrossRef]
- Forestan, C.; Farinati, S.; Zambelli, F.; Pavesi, G.; Rossi, V.; Varotto, S. Epigenetic signatures of stress adaptation and flowering regulation in response to extended drought and recovery in Zea mays. Plant Cell Environ. 2020, 43, 55–75. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Barrett, B.; Brummer, E.C.; Julier, B.; Marshall, A.H. Achievements and Challenges in Improving Temperate Perennial Forage Legumes. Crit. Rev. Plant Sci. 2015, 34, 327–380. [Google Scholar] [CrossRef]
- Osborn, T.C.; Brouwer, D.J.; Kidwell, K.K.; Tavoletti, S.; Bingham, E.T. Molecular Marker Applications to Genetics and Breeding of Alfalfa. In Molecular and Cellular Technologies for Forage Improvement; Brummer, E.C., Hill, N.S., Roberts, C.A., Eds.; Crop Science Society of America: Madison, WI, USA, 1998; pp. 25–31. [Google Scholar] [CrossRef]
- Kumar, T.; Bao, A.K.; Bao, Z.; Wang, F.; Gao, L.; Wang, S.M. The progress of genetic improvement in alfalfa (Medicago sativa L.). Czech J. Genet. Plant Breed. 2018, 54. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Scotti, C.; Carelli, M.; Pecetti, L. Questions and avenues for lucerne improvement. Czech J. Genet. Plant Breed. 2010, 46, 1–13. [Google Scholar] [CrossRef]
- Song, Y.; Lv, J.; Ma, Z.; Dong, W. The mechanism of alfalfa (Medicago sativa L.) response to abiotic stress. Plant Growth Regul. 2019, 89, 239–249. [Google Scholar] [CrossRef]
- Al-Lawati, A.; Al-Bahry, S.; Victor, R.; Al Lawati, A.; Yaish, M. Salt stress alters DNA methylation levels in alfalfa (Medicago spp.). Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Kontsiotou, E. Alfalfa: Cultivation and Use; Agrotypos: Marousi, Greece, 2005; p. 168. [Google Scholar]
- Rinehart, T.A. AFLP analysis using GeneMapper® software and an Excel® macro that aligns and converts output to binary. Biotechniques 2004, 37, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Vekemans, X.; Beauwens, T.; Lemaire, M.; Roldán-Ruiz, I. Data from amplified fragment length polymorphism (AFLP) markers show indication of size homoplasy and of a relationship between degree of homoplasy and fragment size. Mol. Ecol. 2002, 11, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Eckstein, R.L.; Durka, W. Scoring and analysis of methylation-sensitive amplification polymorphisms for epigenetic population studies. Mol. Ecol. Resour. 2013, 13, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Orlóci, L. Multivariate Analysis in Vegetation Research; Springer: Berlin/Heidelberg, Germany, 1978. [Google Scholar] [CrossRef]
- Wilkinson, L. SYSTAT. WIREs Comput. Stat. 2010, 2, 256–257. [Google Scholar] [CrossRef]
- Ruiz-Sánchez, M.C.; Domingo, R.; Torrecillas, A.; Pérez-Pastor, A. Water stress preconditioning to improve drought resistance in young apricot plants. Plant Sci. 2000, 156, 245–251. [Google Scholar] [CrossRef]
- Li, X.; Zhang, L.; Li, Y. Preconditioning Alters Antioxidative Enzyme Responses in Rice Seedlings to Water Stress. Procedia Environ. Sci. 2011, 11, 1346–1351. [Google Scholar] [CrossRef]
- Li, Z.; Shi, P.; Peng, Y. Improved drought tolerance through drought preconditioning associated with changes in antioxidant enzyme activities, gene expression and osmoregulatory solutes accumulation in white clover (Trifolium repens L.). Plant OMICS 2013, 6, 481–489. [Google Scholar]
- Ben Abdallah, M.; Methenni, K.; Nouairi, I.; Zarrouk, M.; Youssef, N.B. Drought priming improves subsequent more severe drought in a drought-sensitive cultivar of olive cv. Chétoui. Sci. Hortic. 2017, 221, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.; Nagy, L.; Hein, R.; Rascher, U.; Beierkuhnlein, C.; Willner, E.; Jentsch, A. Do plants remember drought? Hints towards a drought-memory in grasses. Environ. Exp. Bot. 2011, 71, 34–40. [Google Scholar] [CrossRef]
- Mousavi, S.; Regni, L.; Bocchini, M.; Mariotti, R.; Cultrera, N.G.M.; Mancuso, S.; Googlani, J.; Chakerolhosseini, M.R.; Guerrero, C.; Albertini, E.; et al. Physiological, epigenetic and genetic regulation in some olive cultivars under salt stress. Sci. Rep. 2019, 9, 1093. [Google Scholar] [CrossRef] [PubMed]
- Neves, D.M.; Almeida, L.A.D.H.; Santana-Vieira, D.D.S.; Freschi, L.; Ferreira, C.F.; Soares Filho, W.D.S.; Costa, M.G.C.; Micheli, F.; Coelho Filho, M.A.; Gesteira, A.D.S. Recurrent water deficit causes epigenetic and hormonal changes in citrus plants. Sci. Rep. 2017, 7, 13684. [Google Scholar] [CrossRef]
- Choi, C.-S.; Sano, H. Abiotic-stress induces demethylation and transcriptional activation of a gene encoding a glycerophosphodiesterase-like protein in tobacco plants. Mol. Genet. Genom. 2007, 277, 589–600. [Google Scholar] [CrossRef]
- Garg, R.; Narayana Chevala, V.V.S.; Shankar, R.; Jain, M. Divergent DNA methylation patterns associated with gene expression in rice cultivars with contrasting drought and salinity stress response. Sci. Rep. 2015, 5, 14922. [Google Scholar] [CrossRef] [PubMed]
- Uthup, T.K.; Ravindran, M.; Bini, K.; Thakurdas, S. Divergent DNA Methylation Patterns Associated with Abiotic Stress in Hevea brasiliensis. Mol. Plant 2011, 4, 996–1013. [Google Scholar] [CrossRef]
- Joel, A.J. Epigenetic responses to drought stress in rice (Oryza sativa L.). Physiol. Mol. Biol. Plants 2013, 19, 379–387. [Google Scholar] [CrossRef]
- Berry, S.; Dean, C. Environmental perception and epigenetic memory: Mechanistic insight through FLC. Plant J. 2015, 83, 133–148. [Google Scholar] [CrossRef]
- Wang, W.; Huang, F.; Qin, Q.; Zhao, X.; Li, Z.; Fu, B. Comparative analysis of DNA methylation changes in two rice genotypes under salt stress and subsequent recovery. Biochem. Biophys. Res. Commun. 2015, 465, 790–796. [Google Scholar] [CrossRef]
- Fleta-Soriano, E.; Munné-Bosch, S. Stress Memory and the Inevitable Effects of Drought: A Physiological Perspective. Front. Plant Sci. 2016, 7, 143. [Google Scholar] [CrossRef] [PubMed]
- Bruce, T.J.A.; Matthes, M.C.; Napier, J.A.; Pickett, J.A. Stressful “memories” of plants: Evidence and possible mechanisms. Plant Sci. 2007, 173, 603–608. [Google Scholar] [CrossRef]
- Backhaus, S.; Kreyling, J.; Grant, K.; Beierkuhnlein, C.; Walter, J.; Jentsch, A. Recurrent Mild Drought Events Increase Resistance Toward Extreme Drought Stress. Ecosystems 2014, 17, 1068–1081. [Google Scholar] [CrossRef]
- Ganguly, D.R.; Crisp, P.A.; Eichten, S.R.; Pogson, B.J. The Arabidopsis DNA Methylome Is Stable under Transgenerational Drought Stress. J. Plant Physiol. 2017, 175, 1893–1912. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Gong, Z.; Zhu, J.-K. Abscisic acid-mediated epigenetic processes in plant development and stress responses. J. Integr. Plant Biol. 2008, 50, 1187–1195. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 5′ to 3′ Sequence | |
|---|---|
| EcoRI adapter | CTCGTAGACTGCGTACC AATTGGTACGCAGTC |
| HpaII/MspI adapter | GACGATGAGTCTCGAT CGATCGAGACTCAT |
| Pre-selective EcoRI primer | GACTGCGTACCAATTC-A |
| Pre-selective HpaII/MspI primer | ATGAGTCTCGATCGG-T |
| Selective EcoRI primers | GACTGCGTACCAATTC+ATG GACTGCGTACCAATTC+ACT GACTGCGTACCAATTC+AAC GACTGCGTACCAATTC+AAG |
| Selective HpaII/MspI primer | ATGAGTCTCGATCGG+TCA |
| Treatment | Shoot Dry Weight (g) | Root Dry Weight (g) | Ratio: Shoot/ Root |
|---|---|---|---|
| CC | 6.95 ± 0.34a | 5.66 ± 0.30a | 1.28 ± 0.11a |
| CD2 | 4.04 ± 0.31b | 3.53 ± 0.27b | 1.25 ± 0.11a |
| D1D2 | 2.57 ± 0.35c | 1.79 ± 0.30c | 1.56 ± 0.12a |
| Variety | |||
| Lamia | 5.10 ± 0.21a | 4.42 ± 0.18a | 1.23 ± 0.07b |
| Chaironia | 4.00 ± 0.32b | 2.90 ± 0.28b | 1.50 ± 0.11a |
| Source of variation | |||
| Treatment (A) | p ˂ 0.05 | p ˂ 0.05 | Ns |
| Variety (B) | p < 0.05 | p ˂ 0.05 | p ˂ 0.05 |
| AXB (Interaction) | p < 0.05 | p < 0.05 | Ns |
| Polymorphic Markers | Lamia CC | Lamia CD2 | Lamia D1D2 | Chaironia CC | Chaironia CD2 | Chaironia D1D2 |
|---|---|---|---|---|---|---|
| h methylation | 114 | 87 | 79 | 120 | 79 | 133 |
| m methylation | 102 | 80 | 69 | 146 | 66 | 134 |
| Uninformative | 99 | 101 | 91 | 108 | 104 | 102 |
| Total polymorphic markers | 315 | 268 | 239 | 374 | 249 | 369 |
| Total methylation (h + m) | 216 | 167 | 148 | 266 | 145 | 267 |
| Percentage of total methylation % | 68.57 | 62.31 | 61.92 | 71.12 | 58.23 | 72.36 |
| h Alleles | u Alleles | m Alleles | Total Methylation (h + m) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Iepi | hepi | Iepi | hepi | Iepi | hepi | Iepi | hepi | ||
| Lamia CC | Mean | 0.168 | 0.109 | 0.194 | 0.127 | 0.168c | 0.111c | 0.204f | 0.142f |
| SE | 0.013 | 0.009 | 0.016 | 0.010 | 0.014 | 0.010 | 0.011 | 0.008 | |
| Lamia CD2 | Mean | 0.125 | 0.081 | 0.213 | 0.143 | 0.128 | 0.083 | 0.157 | 0.110 |
| SE | 0.012 | 0.008 | 0.017 | 0.012 | 0.013 | 0.008 | 0.011 | 0.007 | |
| Lamia D1D2 | Mean | 0.118 | 0.077 | 0.189 | 0.126 | 0.112c | 0.073c | 0.140f | 0.097f |
| SE | 0.012 | 0.008 | 0.016 | 0.011 | 0.012 | 0.008 | 0.010 | 0.007 | |
| Chaironia CC | Mean | 0.173a | 0.112a | 0.208 | 0.135 | 0.234d | 0.153d | 0.251g | 0.175g |
| SE | 0.013 | 0.009 | 0.016 | 0.010 | 0.015 | 0.010 | 0.012 | 0.008 | |
| Chaironia CD2 | Mean | 0.114a,b | 0,073a,b | 0.212 | 0.140 | 0.105d,e | 0.068d,e | 0.137g,h | 0095g,h |
| SE | 0.011 | 0,007 | 0.017 | 0.011 | 0.012 | 0.008 | 0.010 | 0.007 | |
| Chaironia D1D2 | Mean | 0.188b | 0.120b | 0.207 | 0.136 | 0.210e | 0.136e | 0.252h | 0.176h |
| SE | 0.013 | 0.008 | 0.016 | 0.011 | 0.014 | 0.009 | 0.012 | 0.008 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ventouris, Y.E.; Tani, E.; Avramidou, E.V.; Abraham, E.M.; Chorianopoulou, S.N.; Vlachostergios, D.N.; Papadopoulos, G.; Kapazoglou, A. Recurrent Water Deficit and Epigenetic Memory in Medicago sativa L. Varieties. Appl. Sci. 2020, 10, 3110. https://doi.org/10.3390/app10093110
Ventouris YE, Tani E, Avramidou EV, Abraham EM, Chorianopoulou SN, Vlachostergios DN, Papadopoulos G, Kapazoglou A. Recurrent Water Deficit and Epigenetic Memory in Medicago sativa L. Varieties. Applied Sciences. 2020; 10(9):3110. https://doi.org/10.3390/app10093110
Chicago/Turabian StyleVentouris, Yannis E., Eleni Tani, Evangelia V. Avramidou, Eleni M. Abraham, Styliani N. Chorianopoulou, Dimitrios N. Vlachostergios, Georgios Papadopoulos, and Aliki Kapazoglou. 2020. "Recurrent Water Deficit and Epigenetic Memory in Medicago sativa L. Varieties" Applied Sciences 10, no. 9: 3110. https://doi.org/10.3390/app10093110
APA StyleVentouris, Y. E., Tani, E., Avramidou, E. V., Abraham, E. M., Chorianopoulou, S. N., Vlachostergios, D. N., Papadopoulos, G., & Kapazoglou, A. (2020). Recurrent Water Deficit and Epigenetic Memory in Medicago sativa L. Varieties. Applied Sciences, 10(9), 3110. https://doi.org/10.3390/app10093110