
Comparative Toxicity of Vegan Red, E124, and E120 Food Dyes on Three Rapidly Proliferating Model Systems

 ,
,  ,
, 

 , ,
, ,  , ,
, ,  , , ,
, , , 
Abstract
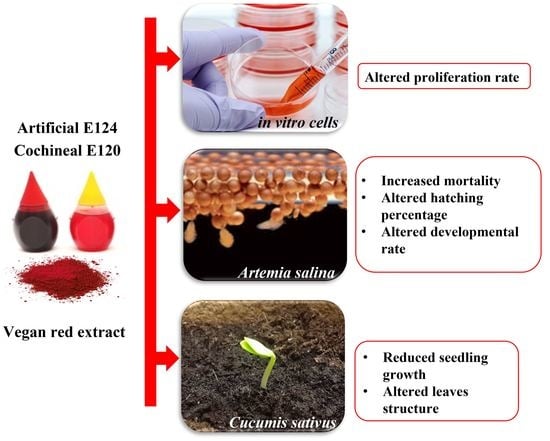
:
1. Introduction
2. Materials and Methods
2.1. Preparation of Red Food Dyes Solutions
2.2. Cell’s Maintenance, Growth, and Viability Test
2.3. Artemia salina Maintenance and Evaluation of Hatching and Growth Rate
2.4. Cucumis sativus Maintenance, Seedling Growth, and Analysis of Leaf Anatomical Structure
2.5. Statistical Analysis
3. Results
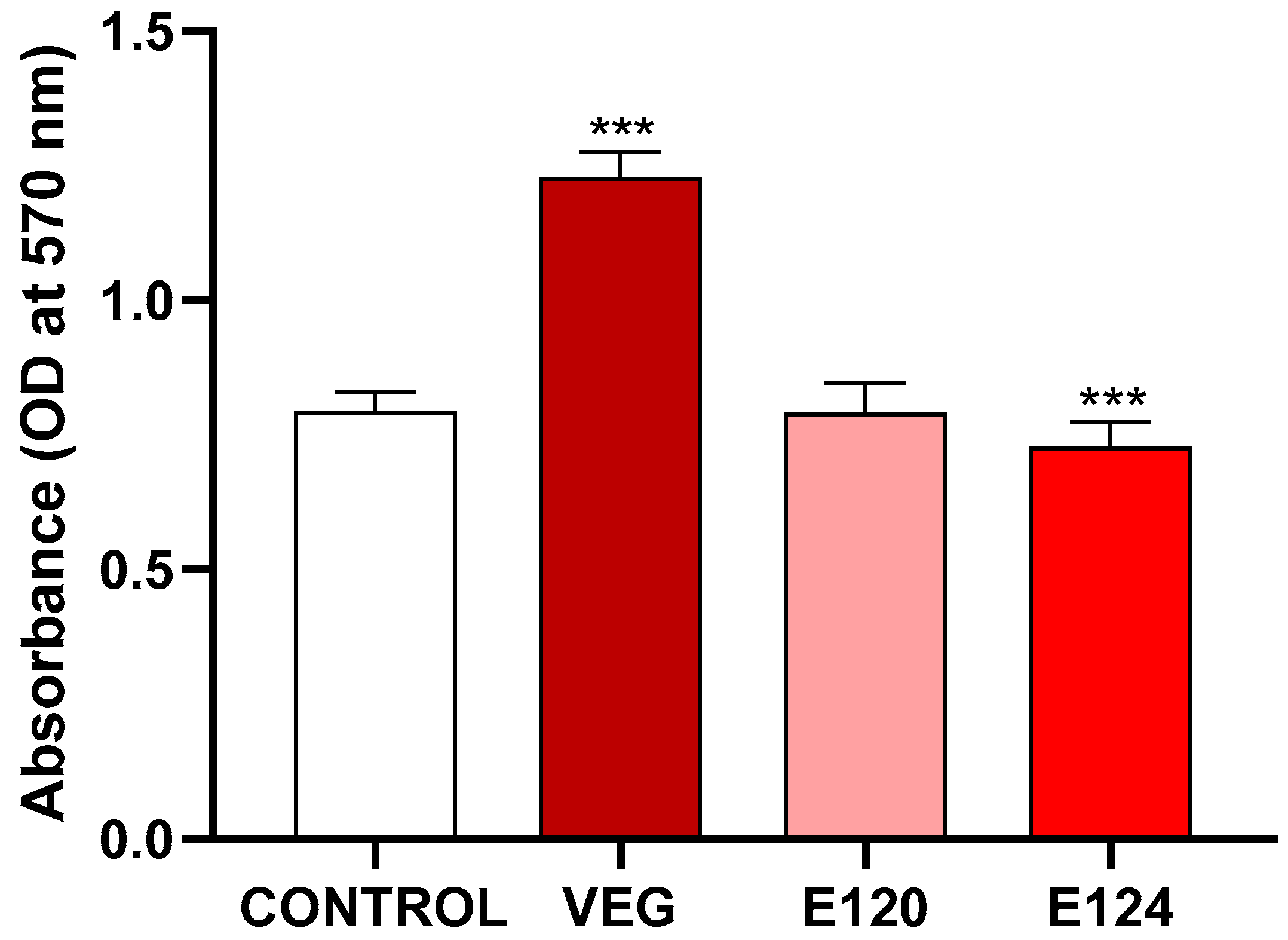
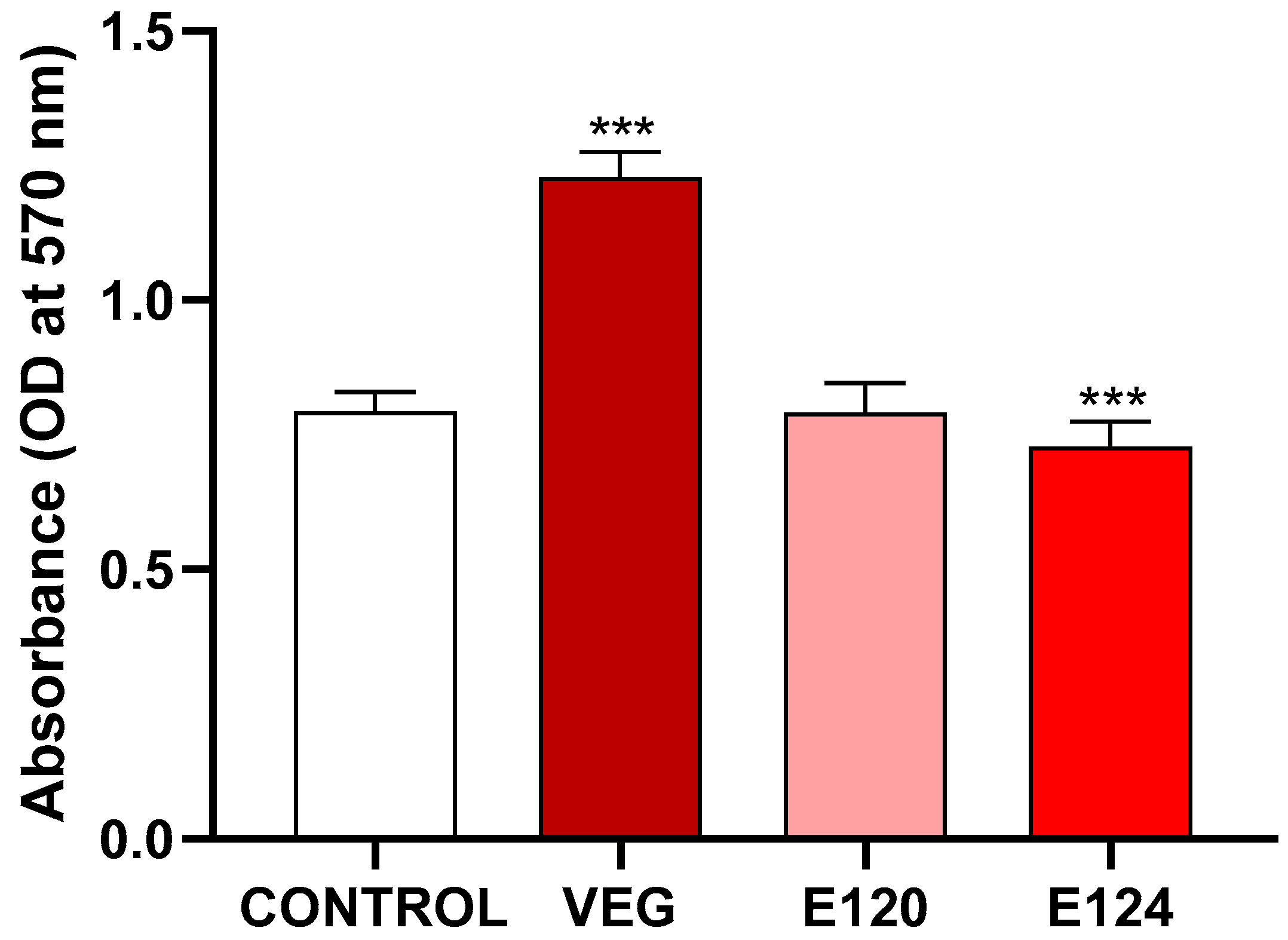
3.1. Effects of the Red Food Dyes on Cell Viability
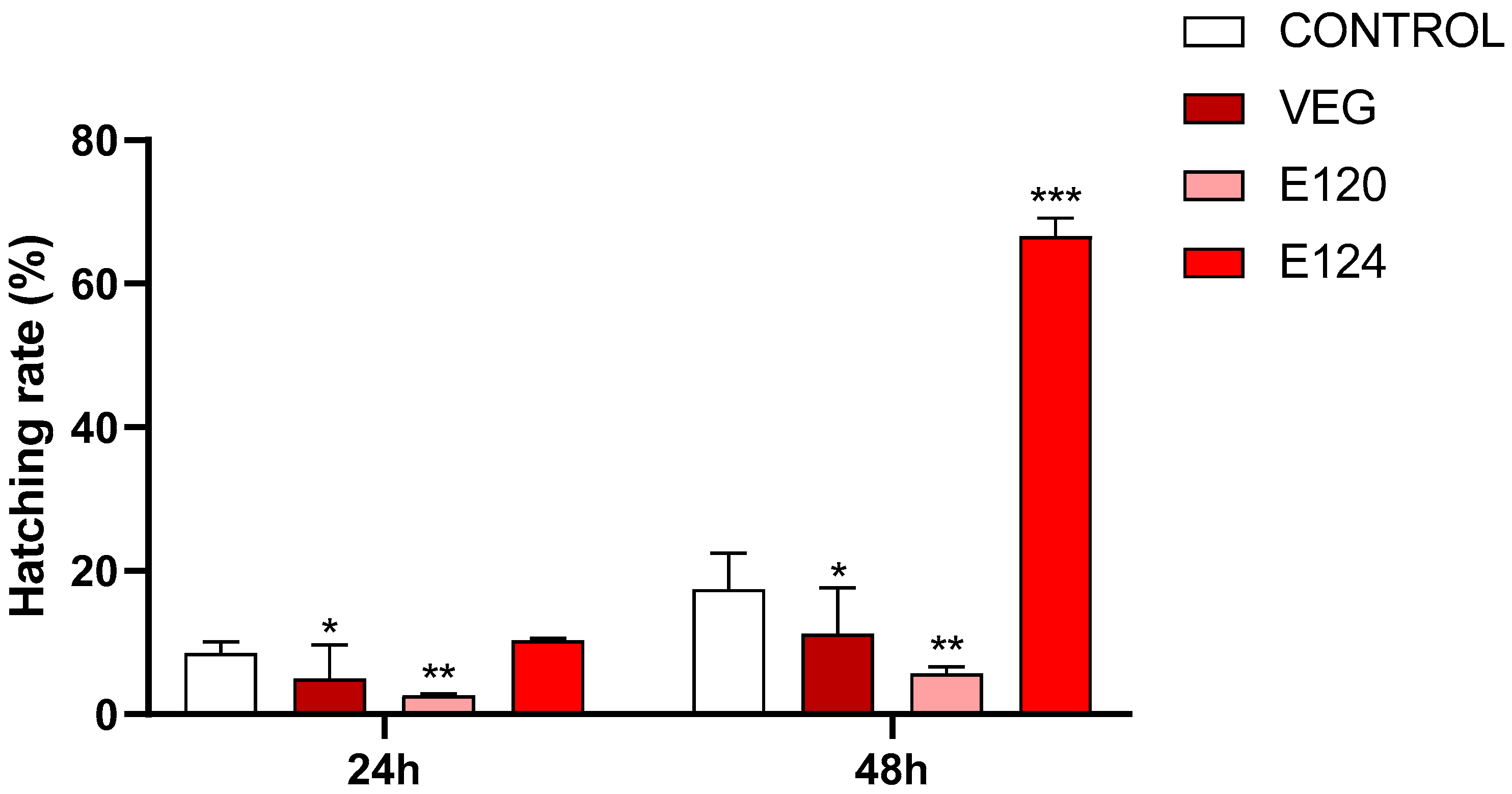
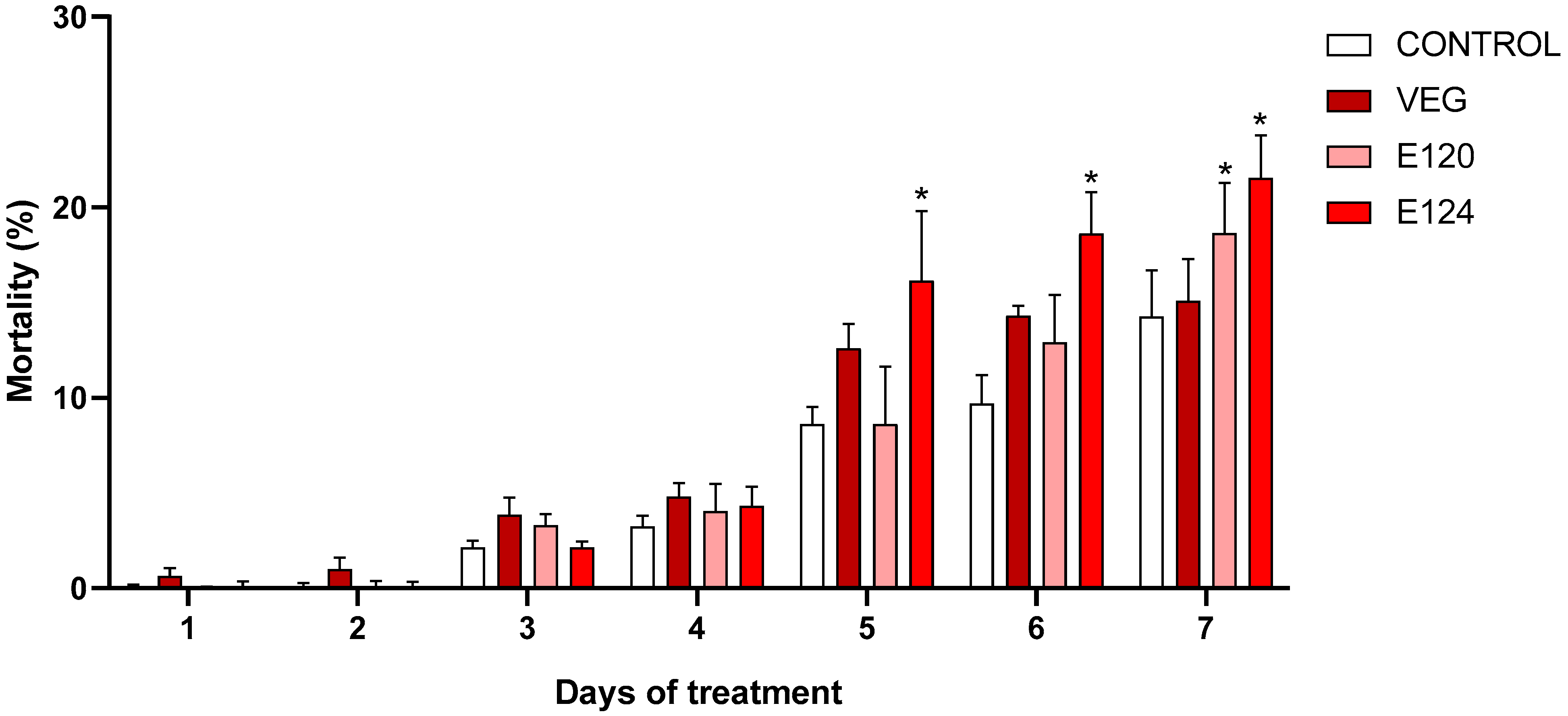
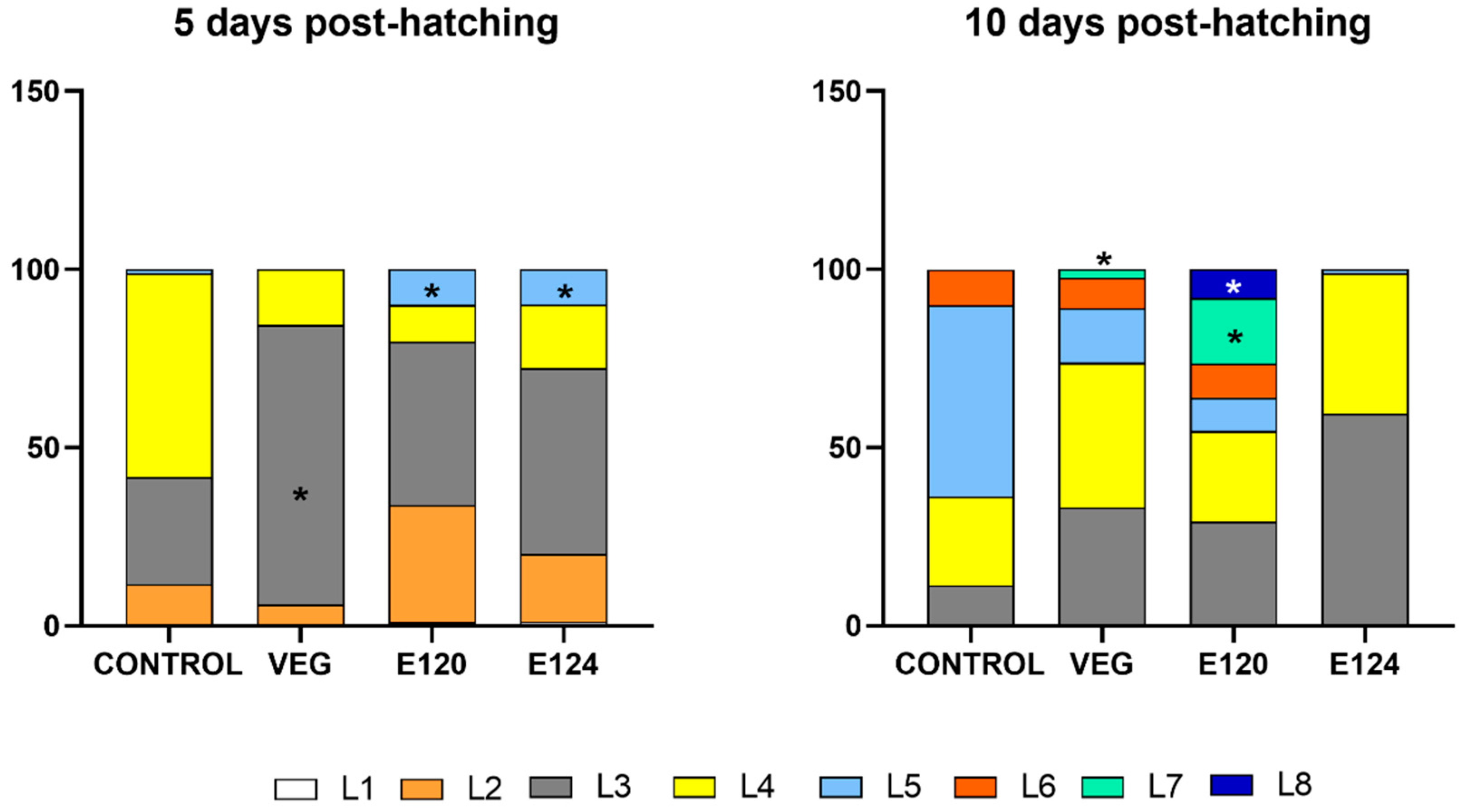
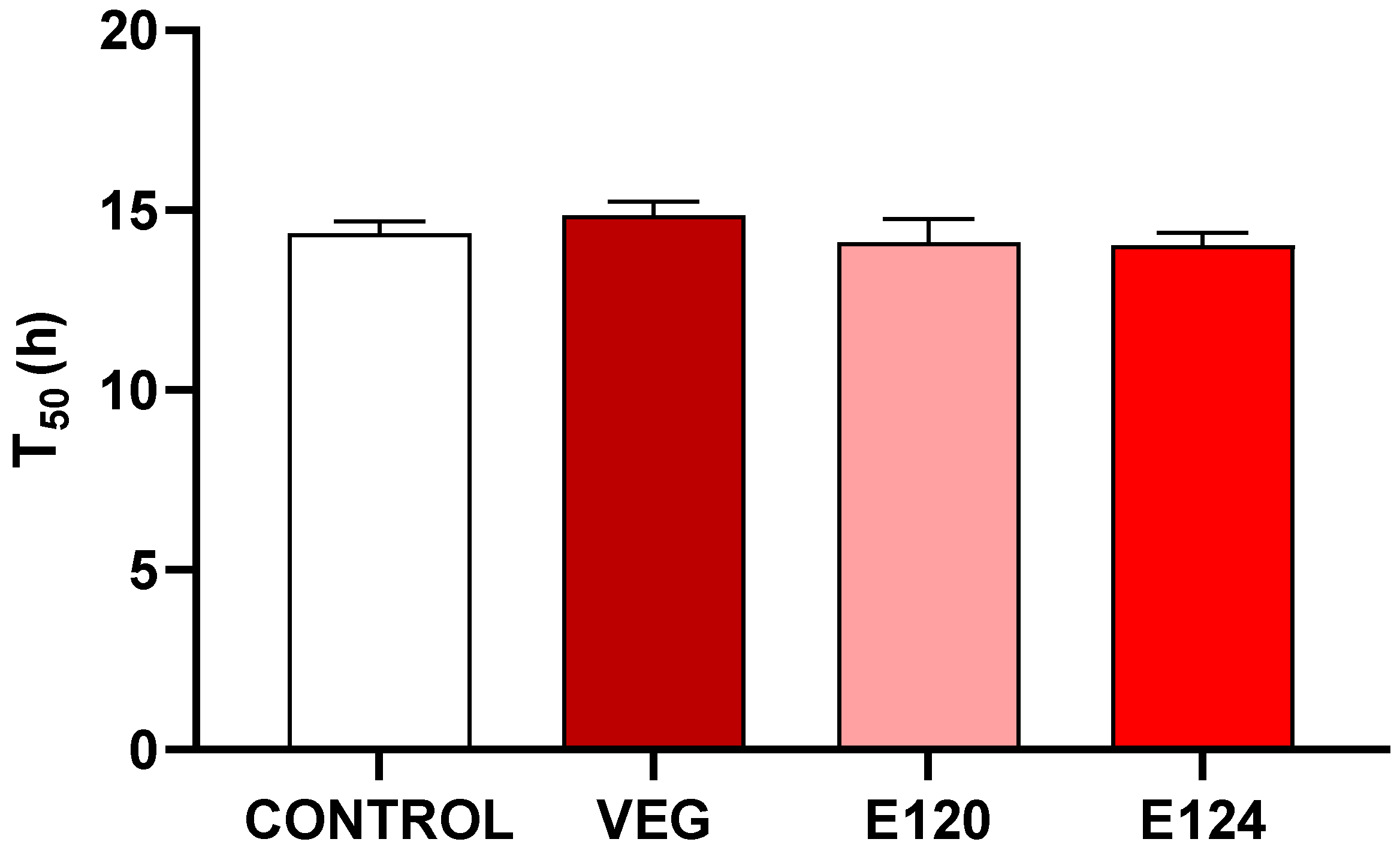
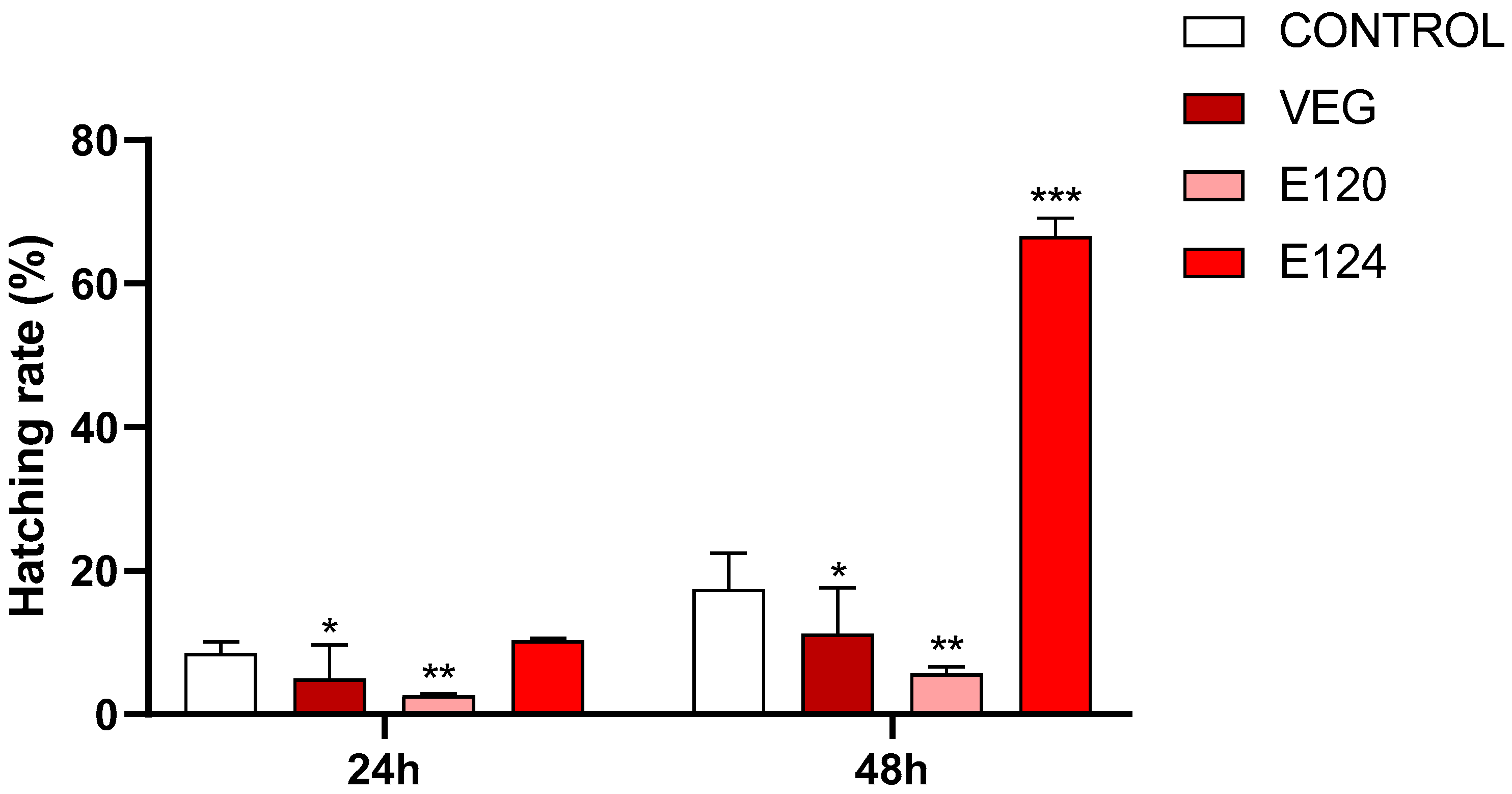
3.2. Effects of Red Food Dyes on the Early Development of Artemia salina Nauplii
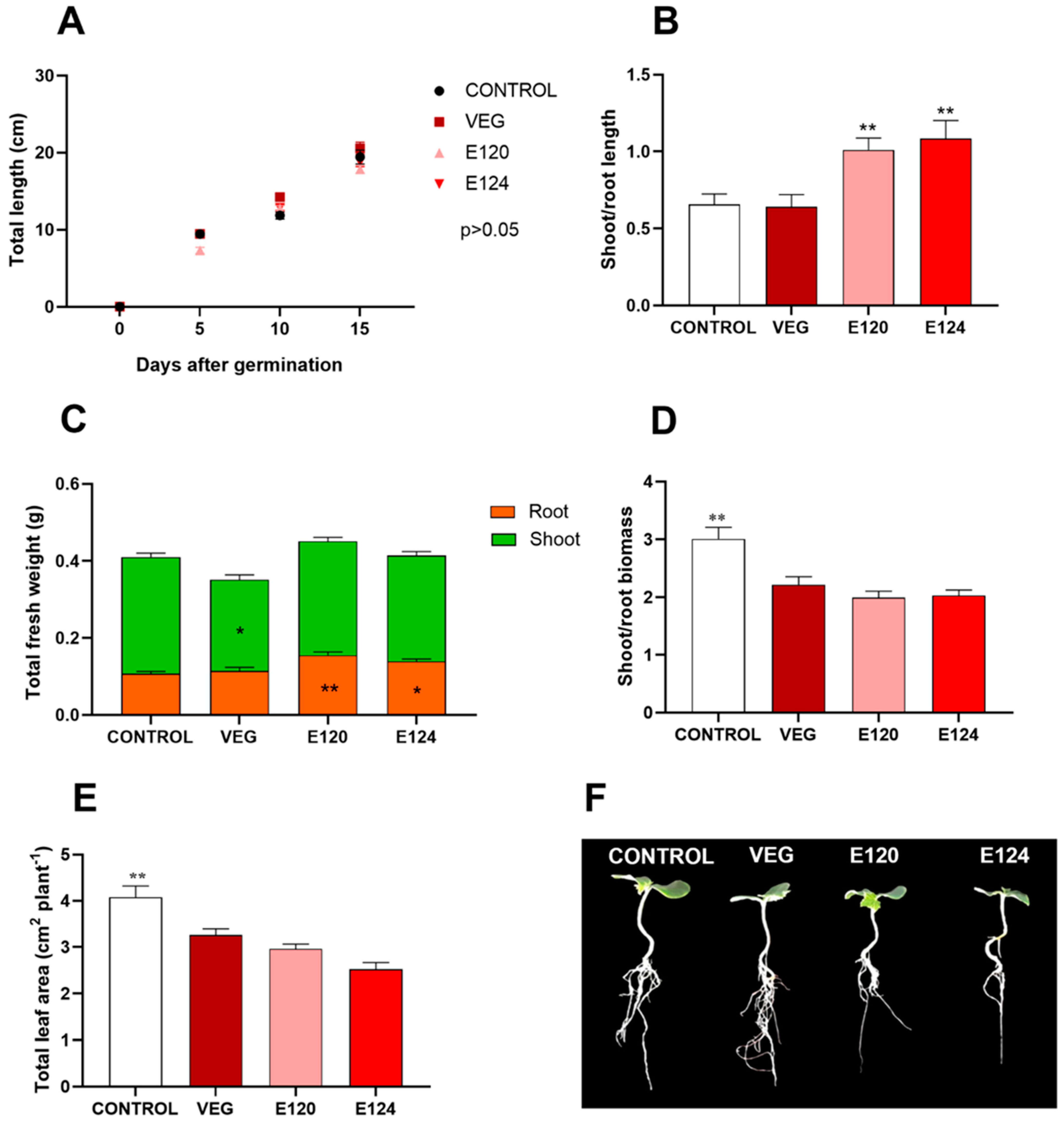
3.3. Effects of Red Food Dyes on the Early Development of Cucumis sativus Seedlings
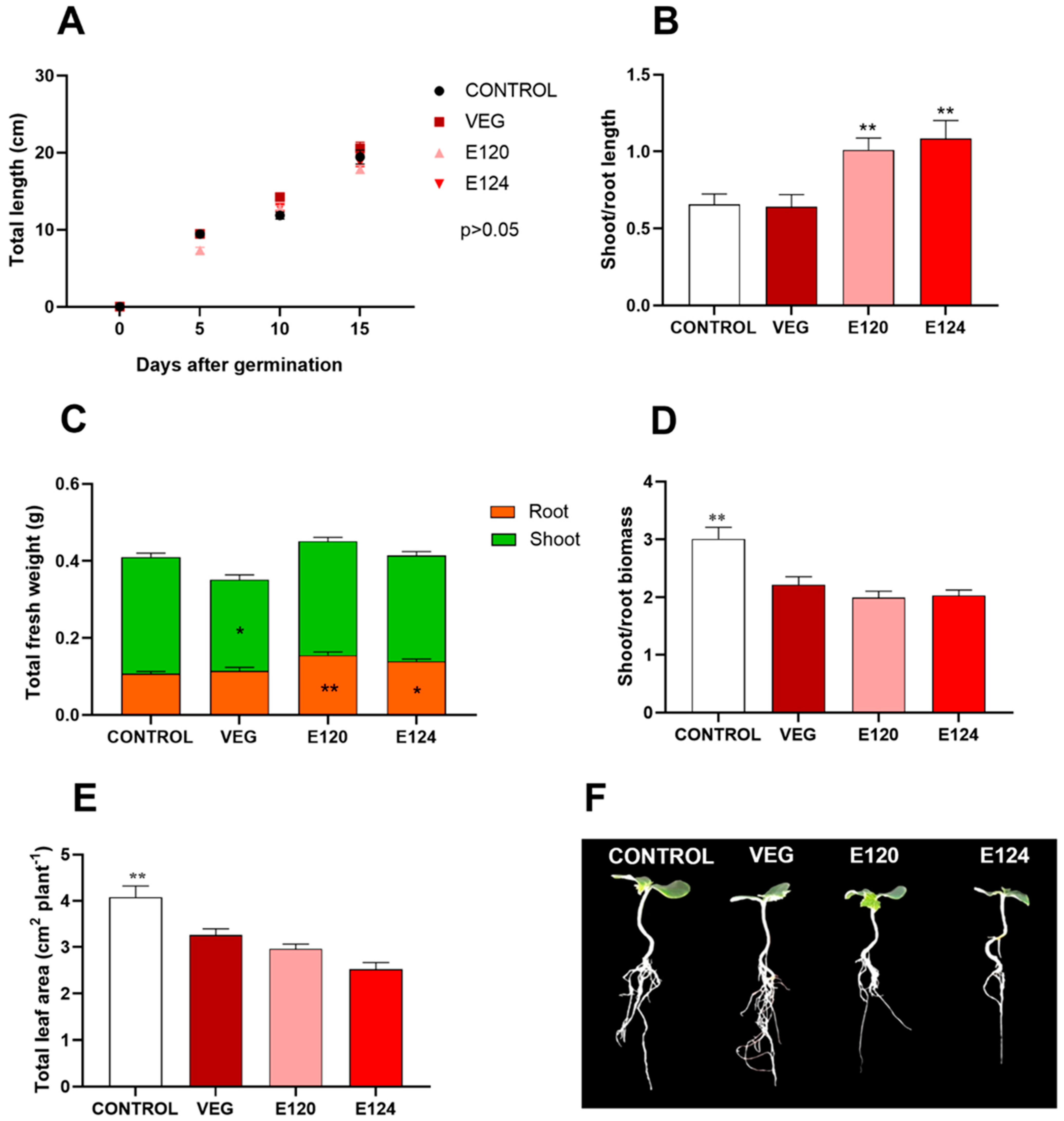
3.3.1. Germination Percentage
3.3.2. Seedling Growth
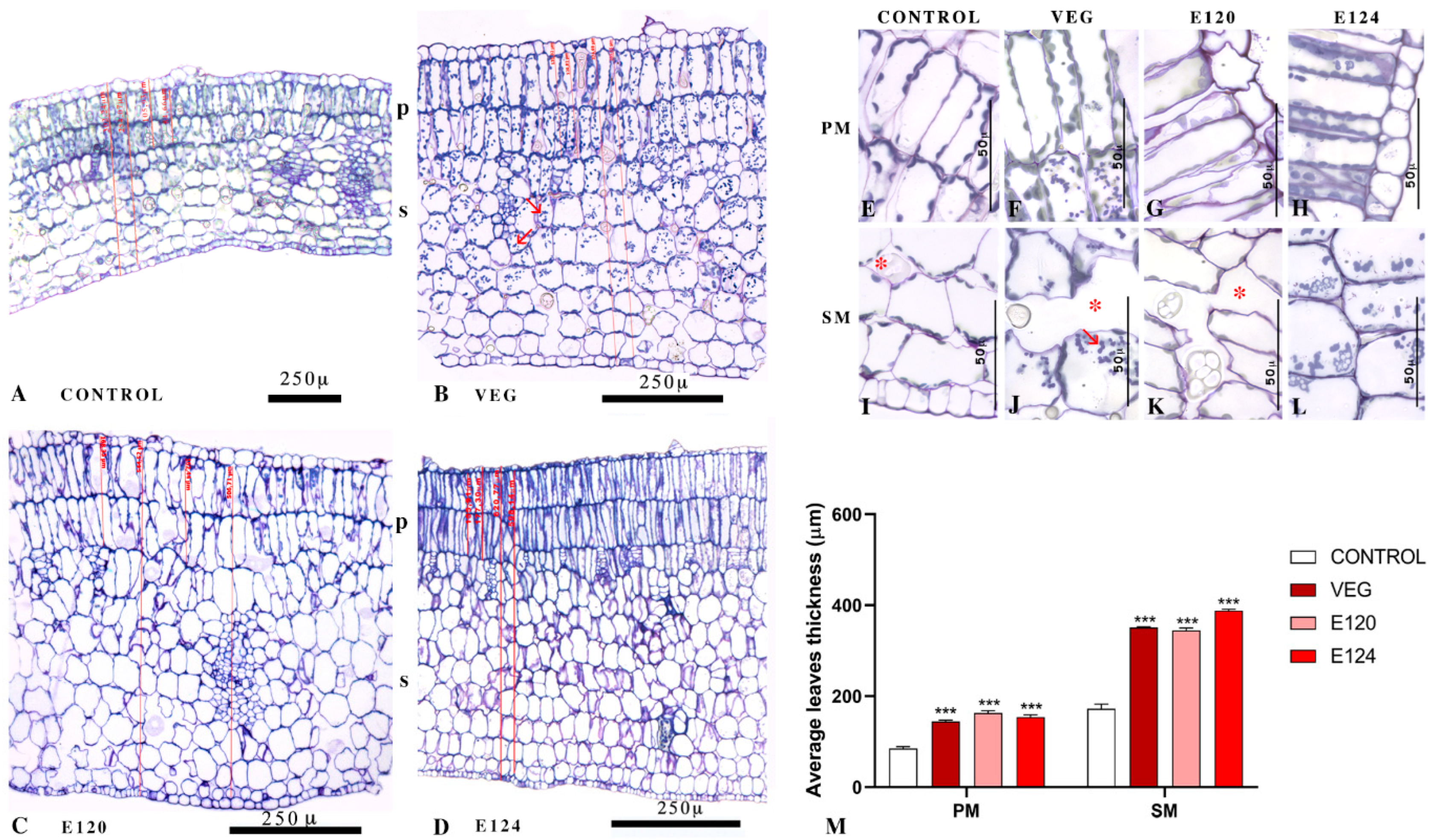
3.3.3. Cotyledonary Leaf Anatomical Structures
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nojavan, S.; Tahmasebi, Z.; Bidarmanesh, T.; Behdad, H.; Nasiri-Aghdam, M.; Mansori, S.; Pourahadi, A. Electrically enhanced liquid-phase microextraction of three textile azo dyes from wastewater and plant samples. J. Sep. Sci. 2013, 36, 3256–3263. [Google Scholar] [CrossRef] [PubMed]
- Natural Food Colors Market Size, Share & Trends Analysis Report by Product (Curcumin, Carotenoids, Anthocyanin, Carmine, Chlorophyllin), by Application, by Region, and Segment Forecasts, 2018–2025. Available online: https://www.grandviewresearch.com/industry-analysis/natural-food-colors-market (accessed on 12 May 2018).
- Lepri, L.; Desideri, P.G.; Coas, V. Separation and identification of water-soluble food dyes by ion-exchange and soap thin-layer chromatography. J. Chromatogr. A 1978, 161, 279–286. [Google Scholar] [CrossRef]
- Perez-Urquiza, M.; Ferrer, R.; Beltran, J.L. Determination of sulfonated azo dyes in river water samples by capillary zone electrophoresis. J. Chromatogr. A 2000, 883, 277–283. [Google Scholar] [CrossRef]
- Vaiano, V.; Iervolino, G.; Rizzo, L.; Sannino, D. Advanced oxidation processes for the removal of food dyes in wastewater. Curr. Org. Chem. 2017, 21, 1068–1073. [Google Scholar] [CrossRef]
- Weiss, B.; Williams, J.H.; Margen, S.; Abrams, B.; Caan, B.; Citron, L.J.; Cox, C.; McKibben, J.; Ogar, D.; Schultz, S. Behavioral responses to artificial food colors. Science 1980, 207, 1487–1489. [Google Scholar] [CrossRef]
- Santana, G.M.; do Anjos Sousa, J.J.; Peron, A.P. Action of Ponceau 4R (E-124) food dye on root meristematic cells of Allium cepa L. Acta Sci. Biol. Sci. 2015, 37, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Motta, C.M.; Simoniello, P.; Arena, C.; Capriello, T.; Panzuto, R.; Vitale, E.; Agnisola, C.; Tizzano, M.; Avallone, B.; Ferrandino, I. Effect of four food dyes on development of three model species, Cucumis sativus, Artemia salina and Danio rerio, assessment of potential risk for environment. Environ. Pollut. 2019, 253, 1126–1135. [Google Scholar] [CrossRef]
- Napolitano, G.; Motta, C.M.; Agnisola, C.; Venditti, P.; Fasciolo, G.; Ferrandino, I.; Capriello, T.; Vitale, E.; Costanzo, G.; Avallone, B.; et al. Commercial red food dyes preparations modulate the oxidative state in three model organisms (Cucumis sativus, Artemia salina and Danio rerio). Environments 2022, 9, 63. [Google Scholar] [CrossRef]
- Shaw, D.W. Allergic contact dermatitis from carmine. Dermatitis. 2009, 20, 292–295. [Google Scholar] [CrossRef]
- Wüthrich, B.; Kägi, M.K.; Stücker, W. Anaphylactic reactions to ingested carmine (E120). Allergy 1997, 52, 1133–1137. [Google Scholar] [CrossRef]
- Quirce, S.; Cuevas, M.; Olaguibel, J.; Tabar, A.I. Occupational asthma and immunologic responses induced by inhaled carmine among employees at a factory making natural dyes. J. Allergy Clin. Immunol. 1994, 93, 44–52. [Google Scholar] [CrossRef]
- EFSA Panel on Food Additives and Nutrient Sources Added to Food. Scientific Opinion on the re-evaluation of Ponceau 4R (E124) as a food additive. EFSA J. 2009, 7, 1328. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Food Additives and Nutrient Sources added to Food. Scientific Opinion on the re-evaluation of cochineal, carminic acid, carmines (E 120) as a food additive. EFSA J. 2015, 13, 4288. [Google Scholar] [CrossRef]
- Brauch, J.E. Underutilized fruits and vegetables as potential novel pigment sources. In Handbook on Natural Pigments in Food and Beverages; Woodhead Publishing: Sawston, UK, 2016; pp. 305–335. [Google Scholar] [CrossRef]
- Ullah, I.; Park, H.Y.; Kim, M.O. Anthocyanins Protect against Kainic Acid-induced Excitotoxicity and apoptosis via ROS-activated AMPK pathway in hippocampal neurons. CNS Neurosci. Ther. 2014, 20, 327–338. [Google Scholar] [CrossRef]
- Manju, M.; Akbarsha, M.A.; Oommen, O.V. In vivo protective effect of dietary curcumin in fish Anabas testudineus (Bloch). Fish Physiol. Biochem. 2012, 38, 309–318. [Google Scholar] [CrossRef]
- Araújo, M.C.; Dias, F.L.; Takahashi, C.S. Potentiation by turmeric and curcumin of gamma-radiation-induced chromosome aberrations in Chinese hamster ovary cells. Teratog. Carcinog. Mutagen. 1999, 19, 9–18. [Google Scholar] [CrossRef]
- Gunes, H.; Gulen, D.; Mutlu, R.; Gumus, A.; Tas, T.; Topkaya, A.E. Antibacterial effects of curcumin, an in vitro minimum inhibitory concentration study. Toxicol. Ind. Health 2016, 32, 246–250. [Google Scholar] [CrossRef]
- Wachter, B.; Syrowatka, M.; Obwaller, A.; Walochnik, J. In vitro efficacy of curcumin on Trichomonas vaginalis. Wien. Klin. Wochenschr. 2014, 126, 32–36. [Google Scholar] [CrossRef]
- Lazzè, M.C.; Savio, M.; Pizzala, R.; Cazzalini, O.; Perucca, P.; Scovassi, A.I.; Bianchi, L. Anthocyanins induce cell cycle perturbations and apoptosis in different human cell lines. Carcinogenesis 2004, 25, 1427–1433. [Google Scholar] [CrossRef]
- Krishnaraju, A.V.; Rao, T.V.; Sundararaju, D.; Vanisree, M.; Tsay, H.S.; Subbaraju, G.V. Assessment of bioactivity of Indian medicinal plants using brine shrimp (Artemia salina) lethality assay. Int. J. Appl. Sci. Eng. 2005, 3, 125–134. [Google Scholar]
- Forte, M.; Di Lorenzo, M.; Iachetta, G.; Mita, D.G.; Laforgia, V.; De Falco, M. Nonylphenol acts on prostate adenocarcinoma cells via estrogen molecular pathways. Ecotoxicol. Environ. Saf. 2019, 180, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Hannas, B.R.; Das, P.C.; Li, H.; LeBlanc, G.A. Intracellular conversion of environmental nitrate and nitrite to nitric oxide with resulting developmental toxicity to the crustacean Daphnia magna. PLoS ONE 2010, 5, e12453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhaecke, P.; Persoone, G.; Claus, C.; Sorgeloos, P. Proposal for a short-term toxicity test with Artemia nauplii. Ecotoxicol. Environ. Saf. 1981, 5, 382–387. [Google Scholar] [CrossRef]
- Motta, C.M.; Cerciello, R.; De Bonis, S.; Mazzella, V.; Cirino, P.; Panzuto, R.; Ciaravolo, M.; Simoniello, P.; Toscanesi, M.; Trifuoggi, M.; et al. Potential toxicity of improperly discarded exhausted photovoltaic cells. Environ. Pollut. 2016, 216, 786–792. [Google Scholar] [CrossRef]
- Copf, T.; Rabet, N.; Celniker, S.E.; Averof, M. Posterior patterning genes and the identification of a unique body region in the brine shrimp Artemia franciscana. Development 2003, 130, 5915–5927. [Google Scholar] [CrossRef] [Green Version]
- Noman, A.; Ali, Q.; Maqsood, J.; Iqbal, N.; Javed, M.T.; Rasool, N.; Naseem, J. Deciphering physio-biochemical, yield, and nutritional quality attributes of water-stressed radish (Raphanus sativus L.) plants grown from Zn-Lys primed seeds. Chemosphere 2018, 195, 175–189. [Google Scholar] [CrossRef]
- Terras, F.R.; Eggermont, K.; Kovaleva, V.; Raikhel, N.V.; Osborn, R.W.; Kester, A.; Rees, S.B.; Torrekens, S.; Van Leuven, F.; Vanderleyden, J. Small cysteine-rich antifungal proteins from radish, their role in host defense. Plant Cell 1995, 7, 573–588. [Google Scholar] [CrossRef] [Green Version]
- Landbo, A.K.; Meyer, A.S. Enzyme-assisted extraction of antioxidative phenols from black currant juice press residues (Ribes nigrum). J. Agric. Food Chem. 2001, 49, 3169–3177. [Google Scholar] [CrossRef]
- Youdim, K.A.; Shukitt-Hale, B.; Martin, A.; Wang, H.; Denisova, N.; Bickford, P.C.; Joseph, J.A. Short-term dietary supplementation of blueberry polyphenolics, beneficial effects on aging brain performance and peripheral tissue function. Nutr. Neurosci. 2000, 3, 383–397. [Google Scholar] [CrossRef]
- Beevi, S.S.; Mangamoori, L.N.; Subathra, M.; Edula, J.R. Hexane extract of Raphanus sativus L. roots inhibits cell proliferation and induces apoptosis in human cancer cells by modulating genes related to apoptotic pathway. Plant Foods Hum. Nutr. 2010, 65, 200–209. [Google Scholar] [CrossRef]
- Jia, N.; Xiong, Y.L.; Kong, B.; Liu, Q.; Xia, X. Radical scavenging activity of black currant (Ribes nigrum L.) extract and its inhibitory effect on gastric cancer cell proliferation via induction of apoptosis. J. Funct. Foods 2012, 4, 382–390. [Google Scholar] [CrossRef]
- Westfall, A.; Sigurdson, G.T.; Rodriguez-Saona, L.E.; Giusti, M.M. Ex Vivo and In Vivo Assessment of the penetration of topically applied anthocyanins utilizing ATR-FTIR/PLS regression models and HPLC-PDA-MS. Antioxidants 2020, 9, 486. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, C.D.C.; Flores, C.S.; He, J.; Tian, Q.; Schwartz, S.J.; Giusti, M.M. Characterization and preliminary bioactivity determination of Lechler fruit anthocyanins. Food Chem. 2011, 128, 717–724. [Google Scholar] [CrossRef]
- Colak, N.; Zengin, A.Y.; Ayaz, F.A. The effect of anthocyanin-rich bilberry extract on the antioxidant system in roots of barley (Hordeum vulgare L.) cultivars under ionizing radiation. Acta Physiol. Plant. 2015, 37, 187. [Google Scholar] [CrossRef]
- Glińska, S.; Bartczak, M.; Oleksiak, S.; Wolska, A.; Gabara, B.; Posmyk, M.; Janas, K. Effects of anthocyanin-rich extract from red cabbage leaves on meristematic cells of Allium cepa L. roots treated with heavy metals. Ecotoxicol. Environ. Saf. 2007, 68, 343–350. [Google Scholar] [CrossRef]
- Tsoyi, K.; Park, H.B.; Kim, Y.M.; Chung, J.I.; Shin, S.C.; Lee, W.S.; Seo, H.G.; Lee, J.H.; Chang, K.C.; Kim, H.J. Anthocyanins from black soybean seed coats inhibit UVB-induced inflammatory cylooxygenase-2 gene expression and PGE2 production through regulation of the nuclear factor-κB and phosphatidylinositol 3-kinase/Akt pathway. J. Agric. Food Chem. 2008, 56, 8969–8974. [Google Scholar] [CrossRef]
- Płatosz, N.; Bączek, N.; Topolska, J.; Szawara-Nowak, D.; Skipor, J.; Milewski, S.; Wiczkowski, W. Chokeberry anthocyanins and their metabolites ability to cross the blood-cerebrospinal fluid barrier. Food Chem. 2021, 346, 128730. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Do anthocyanins function as osmoregulators in leaf tissues? Adv. Bot. Res. 2002, 37, 103–127. [Google Scholar] [CrossRef]
- Carvalho, F.B.; Gutierres, J.M.; Bohnert, C.; Zago, A.M.; Abdalla, F.H.; Vieira, J.M.; Palma, H.E.; Oliveira, S.M.; Spanevello, R.M.; Duarte, M.M.; et al. Anthocyanins suppress the secretion of proinflammatory mediators and oxidative stress, and restore ion pump activities in demyelination. J. Nutr. Biochem. 2015, 26, 378–390. [Google Scholar] [CrossRef]
- Trotman, C.N.A.; Mansfield, B.C.; Tate, W.P. Inhibition of emergence, hatching, and protein biosynthesis in embryonic Artemia salina. Dev. Biol. 1980, 80, 167–174. [Google Scholar] [CrossRef]
- Lye, H.M.; Chiew, J.C.; Siddique, M.M. Cytotoxic effect of commonly used food dyes on human hepatoma cell line. Int. Food Res. J. 2018, 25, 1457–1463. [Google Scholar]
- Dwivedi, K.; Kumar, G. Genetic damage induced by a food coloring dye (sunset yellow) on meristematic cells of Brassica campestris L. J. Environ. Public Health 2015, 2015, 319727. [Google Scholar] [CrossRef] [Green Version]
- e Silva, A.P.S.; de Sousa Silva, T.; dos Santos, A.D.A.; Ribeiro, K.G.; Marques, M.M.M.; de Almeida, P.M.; Peron, A.P. Toxicity of carmine cochineal and caramel iv dyes to terrestrial plants and micro-crustaceans. Water Air Soil Pollut 2020, 231, 313. [Google Scholar] [CrossRef]
- Martinez, H.E.P.; Maia, J.T.L.S.; Ventrella, M.C.; Milagres, C.; Cecon, P.R.; Clemente, J.M.; Garbin, C.Z. Leaf and stem anatomy of cherry tomato under calcium and magnesium deficiencies. Braz. Arch. Biol. Technol. 2020, 63, e20180670. [Google Scholar] [CrossRef]
- Gamboa-Tec, N.; Kú-González, A.; Gutiérrez-Pacheco, L.C.; Castaño, E.; López-Ochoa, L. Anatomic alterations and hyperplasia induced by Euphorbia mosaic virus Yucatan Peninsula isolate at the meshophyll. Rev. Mex. Fitopatol. 2018, 36, 287–297. [Google Scholar] [CrossRef] [Green Version]
- Santos, L.D.T.; Meira, R.M.S.A.; Ferreira, F.A.; Sant’Anna-Santos, B.F.; Ferreira, L.R. Morphological responses of different eucalypt clones submitted to glyphosate drift. Environ. Exp. Bot. 2007, 59, 11–20. [Google Scholar] [CrossRef]
- Merinas-Amo, R.; Martínez-Jurado, M.; Jurado-Güeto, S.; Alonso-Moraga, Á.; Merinas-Amo, T. Biological effects of food coloring in in vivo and in vitro model systems. Foods 2019, 8, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cells | Artemia salina Nauplii | Cucumis sativus Seedlings | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MTT | Hatch | Develop | Mort | Germ | Length | Fresh Biomass | Leaves | ||||||||
| 24 h | 48 h | 5 d | 10 d | 7 d | % | rt | sh/rt | rt | sh | sh/rt | Area | SM | PM | ||
| VEG | + | − | − | − | + | 0 | 0 | 0 | 0 | 0 | − | − | − | + | + |
| E120 | 0 | − | −- | + | + | + | 0 | 0 | + | + | 0 | − | − | + | + |
| E124 | − | 0 | + | + | − | + | 0 | 0 | + | + | 0 | − | − | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avallone, B.; Arena, C.; Simoniello, P.; Di Lorenzo, M.; Vitale, E.; Capriello, T.; Ferrandino, I.; Raggio, A.; Sasso, M.; Napolitano, G.; et al. Comparative Toxicity of Vegan Red, E124, and E120 Food Dyes on Three Rapidly Proliferating Model Systems. Environments 2022, 9, 89. https://doi.org/10.3390/environments9070089
Avallone B, Arena C, Simoniello P, Di Lorenzo M, Vitale E, Capriello T, Ferrandino I, Raggio A, Sasso M, Napolitano G, et al. Comparative Toxicity of Vegan Red, E124, and E120 Food Dyes on Three Rapidly Proliferating Model Systems. Environments. 2022; 9(7):89. https://doi.org/10.3390/environments9070089
Chicago/Turabian StyleAvallone, Bice, Carmen Arena, Palma Simoniello, Mariana Di Lorenzo, Ermenegilda Vitale, Teresa Capriello, Ida Ferrandino, Anja Raggio, Margherita Sasso, Gaetana Napolitano, and et al. 2022. "Comparative Toxicity of Vegan Red, E124, and E120 Food Dyes on Three Rapidly Proliferating Model Systems" Environments 9, no. 7: 89. https://doi.org/10.3390/environments9070089
APA StyleAvallone, B., Arena, C., Simoniello, P., Di Lorenzo, M., Vitale, E., Capriello, T., Ferrandino, I., Raggio, A., Sasso, M., Napolitano, G., De Bonis, S., Fogliano, C., Agnisola, C., & Motta, C. M. (2022). Comparative Toxicity of Vegan Red, E124, and E120 Food Dyes on Three Rapidly Proliferating Model Systems. Environments, 9(7), 89. https://doi.org/10.3390/environments9070089