Mercury Bioavailability in Fluvial Sediments Estimated Using Chironomus riparius and Diffusive Gradients in Thin-Films (DGT)



Abstract
1. Introduction
2. Materials and Methods
2.1. Sediment Sampling and Analysis
2.2. Mercury Bioaccumulation in Chironomus riparius
2.3. Evaluation of Mercury Bioavailability Using DGT Passive Samplers
2.4. Statistical Analysis
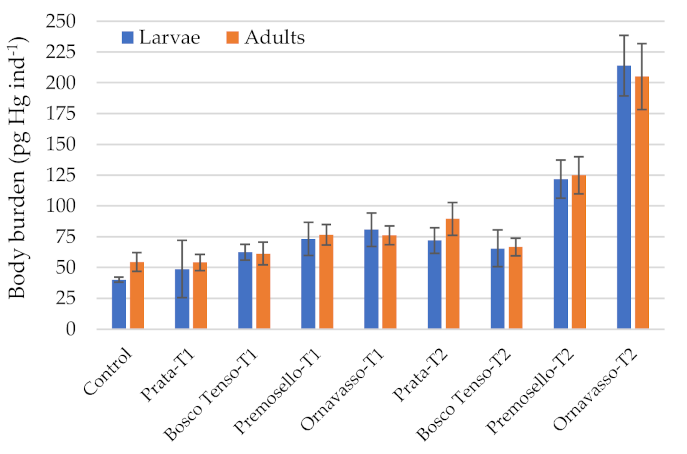
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lavoie, R.A.; Jardine, T.D.; Chumchal, M.M.; Kidd, K.A.; Campbell, L.M. Biomagnification of mercury in aquatic food webs: A worldwide meta-analysis. Environ. Sci. Technol. 2013, 47, 13385–13394. [Google Scholar] [CrossRef] [PubMed]
- Amos, H.M.; Jacob, D.J.; Kocman, D.; Horowitz, H.M.; Zhang, Y.; Dutkiewicz, S.; Horvat, M.; Corbitt, E.S.; Krabbenhoft, D.P.; Sunderland, E.M. Global biogeochemical implications of mercury discharges from rivers and sediment burial. Environ. Sci. Technol. 2014, 48, 9514–9522. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, S.M.; Tanton, T.W.; Abdrashitova, S.A. Mercury in the aquatic environment: A review of factors affecting methylation. Crit. Rev. Environ. Sci. Technol. 2001, 31, 241–293. [Google Scholar] [CrossRef]
- Fernández-Gómez, C.; Dimock, B.; Hintelmann, H.; Díez, S. Development of the DGT technique for Hg measurement in water: Comparison of three different types of samplers in laboratory assays. Chemosphere 2011, 85, 1452–1457. [Google Scholar] [CrossRef] [PubMed]
- Amirbahman, A.; Massey, D.I.; Lotufo, G.; Steenhaut, N.; Brown, L.E.; Biedenbach, J.M.; Magar, V.S. Assessment of mercury bioavailability to benthic macroinvertebrates using diffusive gradients in thin films (DGT). Environ. Sci. Process. Impacts 2013, 15, 2104–2114. [Google Scholar] [CrossRef] [PubMed]
- Harrahy, E.A.; Clements, W.H. Toxicity and bioaccumulation of a mixture of heavy metals in Chironomus tentans (Diptera: Chironomidae) in synthetic sediment. Environ. Toxicol. Chem. 1997, 16, 317–327. [Google Scholar] [CrossRef]
- Žižek, S.; Horvat, M.; Gibičar, D.; Fajon, V.; Toman, M.J. Bioaccumulation of mercury in benthic communities of a river ecosystem affected by mercury mining. Sci. Total. Environ. 2007, 377, 407–415. [Google Scholar] [CrossRef]
- Riva-Murray, K.; Chasar, L.C.; Bradley, P.M.; Burns, D.A.; Brigham, M.E.; Smith, M.J.; Abrahamsen, T.A. Spatial patterns of mercury in macroinvertebrates and fishes from streams of two contrasting forested landscapes in the eastern United States. Ecotoxicology 2011, 20, 1530–1542. [Google Scholar] [CrossRef]
- Bervoets, L.; De Bruyn, L.; Van Ginneken, L.; Blust, R. Accumulation of137Cs by larvae of the midge Chironomus riparius from sediment: Effect of potassium. Environ. Toxicol. Chem. 2003, 22, 1589–1596. [Google Scholar] [CrossRef]
- Chételat, J.; Amyot, M.; Cloutier, L.; Poulain, A.J. Metamorphosis in chironomids, more than mercury supply, controls methylmercury transfer to fish in high arctic lakes. Environ. Sci. Technol. 2008, 42, 9110–9115. [Google Scholar] [CrossRef]
- Kraus, J.M.; Walters, D.M.; Wesner, J.S.; Stricker, C.A.; Schmidt, T.S.; Zuellig, R.E. Metamorphosis alters contaminants and chemical tracers in insects: Implications for food webs. Environ. Sci. Technol. 2014, 48, 10957–10965. [Google Scholar] [CrossRef] [PubMed]
- Simon, E.; Tóthmérész, B.; Kis, O.; Jakab, T.; Szalay, P.E.; Vincze, A.; Baranyai, E.; Harangi, S.; Miskolczi, M.; Dévai, G.; et al. Environmental-friendly Contamination Assessment Of Habitats Based On The Trace Element Content Of Dragonfly Exuviae. Water 2019, 11, 2200. [Google Scholar] [CrossRef]
- Gimbert, F.; Geffard, A.; Guédron, S.; Dominik, J.; Ferrari, B.J.D. Mercury tissue residue approach in Chironomus riparius: Involvement of toxicokinetics and comparison of subcellular fractionation methods. Aquat. Toxicol. 2016, 171, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.P.; Laporte, J.-M.; Andres, S. Factors controlling the bioaccumulation of mercury, methylmercury, arsenic, selenium, and cadmium by freshwater invertebrates and fish. Arch. Environ. Contam. Toxicol. 2000, 38, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Jędruch, A.; Bełdowska, M.; Ziółkowska, M. The role of benthic macrofauna in the trophic transfer of mercury in a low-diversity temperate coastal ecosystem (Puck Lagoon, southern Baltic Sea). Environ. Monit. Assess. 2019, 191, 1–25. [Google Scholar] [CrossRef]
- Zhang, H.; Davison, W. Performance characteristics of diffusion gradients in thin films for the in situ measurement of trace metals in aqueous solution. Anal. Chem. 1995, 67, 3391–3400. [Google Scholar] [CrossRef]
- Ren, M.; Wang, Y.; Ding, S.; Yang, L.; Sun, Q.; Zhang, L. Development of a new diffusive gradient in the thin film (DGT) method for the simultaneous measurement of CH3Hg+ and Hg2+. New J. Chem. 2018, 42, 7976–7983. [Google Scholar] [CrossRef]
- Clarisse, O.; Lotufo, G.R.; Hintelmann, H.; Best, E.P.H. Biomonitoring and assessment of monomethylmercury exposure in aqueous systems using the DGT technique. Sci. Total. Environ. 2012, 416, 449–454. [Google Scholar] [CrossRef]
- Ndu, U.; Christensen, G.A.; Rivera, N.A.; Gionfriddo, C.M.; Deshusses, M.A.; Elias, D.A.; Hsu-Kim, H. Quantification of mercury bioavailability for methylation using diffusive gradient in thin-film samplers. Environ. Sci. Technol. 2018, 52, 8521–8529. [Google Scholar] [CrossRef]
- Xu, X.; Bryan, A.L.; Mills, G.L.; Korotasz, A.M. Mercury speciation, bioavailability, and biomagnification in contaminated streams on the Savannah River Site (SC, USA). Sci. Total. Environ. 2019, 668, 261–270. [Google Scholar] [CrossRef]
- Pisanello, F.; Marziali, L.; Rosignoli, F.; Poma, G.; Roscioli, C.; Pozzoni, F.; Guzzella, L. In situ bioavailability of DDT and Hg in sediments of the Toce River (Lake Maggiore basin, Northern Italy): Accumulation in benthic invertebrates and passive samplers. Environ. Sci. Pollut. Res. 2015, 23, 10542–10555. [Google Scholar] [CrossRef] [PubMed]
- Marziali, L.; Rosignoli, F.; Drago, A.; Pascariello, S.; Valsecchi, L.; Rossaro, B.; Guzzella, L. Toxicity risk assessment of mercury, DDT and arsenic legacy pollution in sediments: A triad approach under low concentration conditions. Sci. Total. Environ. 2017, 593–594, 809–821. [Google Scholar] [CrossRef] [PubMed]
- US Environmental Protection Agency (US-EPA). Method 7473—Mercury in Solids and Solutions by Thermal Decomposition, Amalgamation, and Atomic Absorption Spectrophotometry; US-EPA: Washington, DC, USA, 1998. [Google Scholar]
- Schumacher, B.A. Methods for the Determination of Total Organic Carbon (TOC) in Soils and Sediments; Ecological Risk Assessment Support Center: Las Vegas, NV, USA, April 2002. [Google Scholar]
- De Vos, B.; Lettens, S.; Muys, B.; Deckers, J.A. Walkley–Black analysis of forest soil organic carbon: Recovery, limitations and uncertainty. Soil Use Manag. 2007, 23, 221–229. [Google Scholar] [CrossRef]
- Azevedo-Pereira, H.M.V.S.; Soares, A.M.V.M. Effects of mercury on growth, emergence, and behavior of Chironomus riparius Meigen (Diptera: Chironomidae). Arch. Environ. Contam. Toxicol. 2010, 59, 216–224. [Google Scholar] [CrossRef]
- Organisation for Economic Co-operation and Development (OECD). Test No. 233: Sediment-Water Chironomid Toxicity Using Spiked Sediment. OECD Guidelines for the Testing of Chemicals; OECD Publishing: Paris, France, 2004. [Google Scholar]
- Vogt, C.; Nowak, C.; Diogo, J.B.; Oetken, M.; Schwenk, K.; Oehlmann, J. Multi-generation studies with Chironomus riparius—Effects of low tributyltin concentrations on life history parameters and genetic diversity. Chemosphere 2007, 67, 2192–2200. [Google Scholar] [CrossRef]
- Gimbert, F.; de Vaufleury, A.; Douay, F.; Scheifler, R.; Coeurdassier, M.; Badot, P.-M. Modelling chronic exposure to contaminated soil: A toxicokinetic approach with the terrestrial snail Helix aspersa. Environ. Int. 2006, 32, 866–875. [Google Scholar] [CrossRef]
- Docekalová, H.; Diviš, P. Application of diffusive gradient in thin films technique (DGT) to measurement of mercury in aquatic systems. Talanta 2005, 65, 1174–1178. [Google Scholar] [CrossRef]
- Diviš, P.; Leermakers, M.; Dočekalová, H.; Gao, Y. Mercury depth profiles in river and marine sediments measured by the diffusive gradients in thin films technique with two different specific resins. Anal. Bioanal. Chem. 2005, 382, 1715–1719. [Google Scholar] [CrossRef]
- Pelcová, P.; Dočekalová, H.; Kleckerová, A. Development of the diffusive gradient in thin films technique for the measurement of labile mercury species in waters. Anal. Chim. Acta 2014, 819, 42–48. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Chibunda, R.T. Chronic toxicity of mercury (HgCl2) to the benthic midge Chironomus riparius. Int. J. Environ. Res. 2009, 3, 455–462. [Google Scholar] [CrossRef]
- Valsecchi, L.; Roscioli, C.; Zanini, C.; Schiavon, A.; Guzzella, L.; Marziali, L. Determinazione del metilmercurio in sedimenti e biota d’acqua dolce mediante analizzatore automatico di mercurio e GC-MS. Not. Metodi Anal. IRSA News 2020, 1, 20–29. (In Italian) [Google Scholar]
- Hong, Y.S.; Rifkin, E.; Bouwer, E.J. Combination of diffusive gradient in a thin film probe and IC-ICP-MS for the simultaneous determination of CH3Hg+and Hg2+in oxic water. Environ. Sci. Technol. 2011, 45, 6429–6436. [Google Scholar] [CrossRef] [PubMed]
- Clarisse, O.; Foucher, D.; Hintelmann, H. Methylmercury speciation in the dissolved phase of a stratified lake using the diffusive gradient in thin film technique. Environ. Pollut. 2009, 157, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, B.J.D.; Vignati, D.A.L.; Roulier, J.-L.; Coquery, M.; Szalińska, E.; Bobrowski, A.; Czaplicka, A.; Dominik, J. Chromium bioavailability in aquatic systems impacted by tannery wastewaters. Part 2: New insights from laboratory and in situ testing with Chironomus riparius Meigen (Diptera, Chironomidae). Sci. Total. Environ. 2019, 653, 1–9. [Google Scholar] [CrossRef]
- Tsui, M.T.K.; Wang, W.-X. Uptake and elimination routes of inorganic mercury and methylmercury in Daphnia magna. Environ. Sci. Technol. 2003, 38, 808–816. [Google Scholar] [CrossRef]
- Pan, K.; Wang, W.-X. Mercury accumulation in marine bivalves: Influences of biodynamics and feeding niche. Environ. Pollut. 2011, 159, 2500–2506. [Google Scholar] [CrossRef]
- Rossaro, B.; Gaggino, G.F.; Marchetti, F. Accumulation of mercury in larvae and adults, Chironomus riparius (Meigen). Bull. Environ. Contam. Toxicol. 1986, 37, 402–406. [Google Scholar] [CrossRef]
- Cid, N.; Ibáñez, C.; Palanques, A.; Prat, N. Patterns of metal bioaccumulation in two filter-feeding macroinvertebrates: Exposure distribution, inter-species differences and variability across developmental stages. Sci. Total. Environ. 2010, 408, 2795–2806. [Google Scholar] [CrossRef]
- Lepom, P.; Irmer, U.; Wellmitz, J. Mercury levels and trends (1993–2009) in bream (Abramis brama L.) and zebra mussels (Dreissena polymorpha) from german surface waters. Chemosphere 2012, 86, 202–211. [Google Scholar] [CrossRef]
- International Commission for the Protection of the Italian-SwissWaters. Indagini Sulle Sostanze Pericolose Nell’ecosistema del Lago Maggiore. Programma 2016–2018; CIPAIS: Verbania, Italy, 2018; Available online: http://www.cipais.org/ (accessed on 13 January 2021).
- International Commission for the Protection of the Italian-Swiss Waters. Indagini Sulle Sostanze Pericolose Nell’ecosistema del Lago Maggiore. Programma 2019–2021; CIPAIS: Verbania, Italy, 2020; Available online: http://www.cipais.org/ (accessed on 13 January 2021).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test 1 | Test 2 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C. riparius | DGT Pistons | DGT Probes | C. riparius | ||||||||||||
| Exposure (days) | 7 | 11 | 16 | 0 | 7 | 11 | 16 | 28 | 0 | 7 | 11 | 16 | 28 | 21 | |
| Sediments | |||||||||||||||
| Blank | 2 | 2 | |||||||||||||
| Control | 4 | - | 3(2) | - | - | 2 | - | - | - | 2 | - | 3(2) | |||
| Prata | - | - | 3(2) | - | - | 2 | - | - | - | 2 | - | 3(2) | |||
| Bosco Tenso | - | - | 3(2) | - | - | 2 | - | - | - | 2 | - | 3(2) | |||
| Premosello | 4 | 3 | 3(2) | 2 | 2 | 2 | 2 | 2 | 2 | 2 | - | 3(2) | |||
| Ornavasso | 4 | 3 | 3(2) | 2 | 2 | 2 | 2 | 2 | 2 | 2 | - | 3(2) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marziali, L.; Valsecchi, L. Mercury Bioavailability in Fluvial Sediments Estimated Using Chironomus riparius and Diffusive Gradients in Thin-Films (DGT). Environments 2021, 8, 7. https://doi.org/10.3390/environments8020007
Marziali L, Valsecchi L. Mercury Bioavailability in Fluvial Sediments Estimated Using Chironomus riparius and Diffusive Gradients in Thin-Films (DGT). Environments. 2021; 8(2):7. https://doi.org/10.3390/environments8020007
Chicago/Turabian StyleMarziali, Laura, and Lucia Valsecchi. 2021. "Mercury Bioavailability in Fluvial Sediments Estimated Using Chironomus riparius and Diffusive Gradients in Thin-Films (DGT)" Environments 8, no. 2: 7. https://doi.org/10.3390/environments8020007
APA StyleMarziali, L., & Valsecchi, L. (2021). Mercury Bioavailability in Fluvial Sediments Estimated Using Chironomus riparius and Diffusive Gradients in Thin-Films (DGT). Environments, 8(2), 7. https://doi.org/10.3390/environments8020007